Морфолиновые олигонуклеотиды
Морфоли́новые олигонуклеоти́ды[1][2][3], или морфоли́новые олигоме́ры[4] (англ. morpholino, morpholino oligomer, phosphorodiamidate morpholino oligomer, PMO) — синтетические олигонуклеотиды, использующиеся в молекулярной биологии для изменения экспрессии генов. Основу молекулярной структуры морфолинового олигонуклеотида составляют метиленморфолиновые кольца и фосфородиамидатные связи. Морфолиновые олигонуклеотиды блокируют доступ других молекул к небольшим (длиной около 25 нуклеотидов) специфическим последовательностям за счёт комплементарного спаривания с соответствующей РНК. Морфолиновые олигонуклеотиды служат исследовательскими инструментами в обратной генетикe для нокдауна гена.
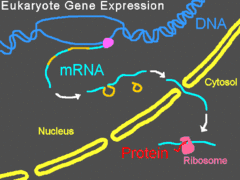
Нокдаун гена заключается в предотвращении синтеза клеткой соответствующего белка[5]. Морфолиновые олигонуклеотиды также могут влиять на сплайсинг пре-мРНК[6]. Нокдаун гена является мощным средством для изучения функции конкретного гена. Аналогичным образом, вырезание специфического экзона из транскрипта позволяет определить функции, выполняемые соответствующими аминокислотными остатками белка, а иногда приводит к полной инактивации белка. Морфолиновые олигонуклеотиды используются в изучении многих модельных организмов, в том числе мышей, данио-рерио, лягушек и морских ежей[7].
Разрабатываются пути применения морфолиновых олигонуклеотидов в медицине против патогенных организмов (бактерий[8] и вирусов[9]), а также для ослабления проявления генетических заболеваний[10].
Структура
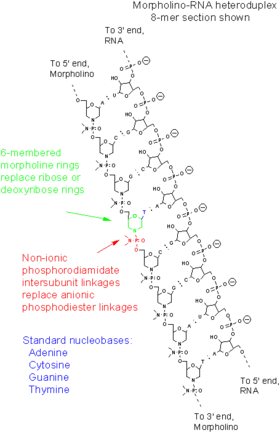
Морфолиновые олигонуклеотиды — это синтетические молекулы, представляющие собой измененные нуклеиновые кислоты[11]. Они, как правило, имеют длину 25 нуклеотидов и комплементарно связываются с соответствующими последовательностями РНК за счёт спаривания оснований. Морфолиновые олигонуклеотиды имеют обычные азотистые основания, которые вместо дезоксирибозы соединены с кольцами морфолина, а в связи отдельных нуклеотидов участвуют не фосфатные, а фосфородиамидатные группы. Замена отрицательно заряженных фосфатных групп незаряженной фосфородиамидатной группой устраняет ионизацию при физиологических значениях рН, так что в живых клетках эти молекулы не заряжены. Вся молекула морфолинового олигонуклеотида состоит из таких изменённых нуклеотидов[11].
Функции
Механизмы действия морфолиновых олигонуклеотидов
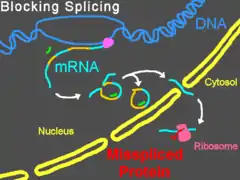
Морфолиновые олигонуклеотиды не вызывают деградации своей РНК-мишени, в отличие от многих антисмысловых молекул (например, малых интерферирующих РНК и фосфоротиоатов). Вместо этого они выступают в роли «пространственных блокаторов», связываясь с последовательностью-мишенью и физически не давая другим молекулам связаться с этой РНК[5]. Морфолиновые олигонуклеотиды часто используются для изучения роли специфических мРНК в развивающемся зародыше. Эмбриологи инъецируют их в яйца или зародыши данио-рерио[12], шпорцевой лягушки[13], морских ежей[14] и рыбы Fundulus heteroclitus или доставляют эти молекулы путём электропорации в зародыши курицы на более поздних стадиях развития[15]; обработанные таким образом зародыши называют морфантами. При наличии в цитозоле правильных систем доставки морфолиновых олигонуклеотидов могут быть эффективными и в культурах клеток[16][17].
Блокирование трансляции
Связываясь с 5'-нетранслируемой областью мРНК, морфолиновые олигонуклеотиды могут мешать рибосоме в комплексе с эукариотическими факторами инициации трансляции двигаться от кэпа к старт-кодону. Это предотвращает трансляцию кодирующей области транскрипта-мишени (нокдаун гена). Такой приём очень удобен, когда исследователь хочет определить функцию конкретного белка; по эффектам, оказываемым нокдауном гена на клетку или организм, судят о функции белка. Некоторые морфолиновые олигонуклеотиды блокируют экспрессию столь эффективно, что после деградации белка, синтезированного до введения олигонуклеотидов, этот белок становится неопределимым при помощи вестерн-блоттинга[18].
Изменение сплайсинга пре-мРНК
Морфолиновые олигонуклеотиды могут вмешиваться в процессинг пре-мРНК следующими способами:
- предотвращая связывание направляющих сплайсинг малых ядерных рибонуклеопротеинов (snRNP) со своими мишенями на границах интронов в пре-мРНК;
- блокируя остаток аденина, осуществляет нуклеофильную атаку в ходе сплайсинга и образует структуру лассо;
- мешая связыванию регуляторных белков сплайсинга, таких как сайленсеры[19] и энхансеры сплайсинга[20].
Предотвращение связывания рибонуклеопротеинов U1 (в сайте-доноре) или U2/U5 (в полипиримидиновом тракте или сайте-акцепторе) может привести к модифицированному сплайсингу, при котором из зрелой мРНК исключаются экзоны. Воздействие на другие сайты сплайсинга приводит к включению интронов в зрелую мРНК, в то время как активация скрытых сайтов сплайсинга может приводит и к включениям, и к исключениям[21]. Морфолиновые олигонуклеотиды могут также блокировать мишени snRNP U11/U12[22]. Изменения в сплайсинге удобно отслеживать при помощи ПЦР с обратной транскрипцией (RT-PCR) и электрофореза (при электрофорезе продуктов RT-PCR наблюдается смещение полос в геле)[6].
Другие применения
Морфолиновые олигонуклеотиды использовались для блокировки активности микроРНК[23][24], а также их созревания[25]. Морфолиновые олигонуклеотиды, меченные флуоресцеином, в сочетании с антителами, специфичными к флуоресцеину, могут использоваться в качестве проб in situ-гибридизации с микроРНК[26]. Кроме того, они могут блокировать активность рибозимов[27], а также функционирование snRNP U2 и U12[28]. Морфолиновые олигонуклеотиды, нацеленные на «оголённые» последовательности мРНК в пределах кодирующей области, могут вызывать сдвиг рамки считывания при трансляции[29]. Они также могут блокировать редактирование РНК. Эффективность морфолиновых олигонуклеотидов по отношению ко многим мишеням делает их универсальным средством для подавления взаимодействия белков или нуклеиновых кислот с мРНК[30].
Специфичность, стабильность и не-антисмысловые эффекты
Морфолиновые олигонуклеотиды стали стандартным инструментом для нокдауна генов в эмбриональных системах животных, в которых экспрессируется больше генов, чем во взрослых клетках. После инъекции морфолинового олигонуклеотида в зародыши лягушки или рыбы на стадии одной или нескольких клеток наибольший эффект проявляется к пятому дню[31], когда большая часть органогенеза и дифференцировки клеток и тканей уже позади; наблюдаемые фенотипы соответствуют нокауту определённого гена. Контрольные нуклеотиды, не нацеленные на какие-либо последовательности, как правило, не вызывают изменений в фенотипе зародыша, подтверждая, что морфолиновые олигонуклеотиды действуют специфично к последовательностям и, как правило, не имеют не-антисмысловых эффектов. Дозу, необходимую для нокдауна гена, можно сократить, одновременно вводя несколько морфолиновых олигонуклеотидов, нацеленных на одну и ту же мРНК; этот приём позволяет сократить или вовсе исключить зависимые от дозы взаимодействия олигонуклеотидов с РНК, не являющимися непосредственными мишенями[32].
В экспериментах по «спасению» мРНК у зародышей часто удавалось вернуть фенотип дикого типа. В ходе этих экспериментов морфолиновый олигонуклеотид вводится в клетку вместе с мРНК, кодирующей тот белок, на нокдаун которого этот олигонуклеотид и направлен. Однако вводимая мРНК имеет изменённую 5'-нетранслируемую область, а потому не имеет последовательности-мишени для морфолинового олигонуклеотида, но её кодирующая область сохранена и с неё синтезируется интересующий белок. Трансляция с мРНК-«спасателя» восстанавливает образование белка, остановленное морфолиновым олигонуклеотидом. Спасательная мРНК не вызывает фенотипических отклонений, так как не действует на экспрессию генов, не являющихся мишенью введенного олигонуклеотида, поэтому возврат к фенотипу дикого типа служит ещё одним доказательством специфичности морфолиновых олигонуклеотидов по отношению к последовательности[31].
Из-за совершенно неестественного химического строения морфолиновые олигонуклеотиды не распознаются клеточными белками. Нуклеазы их не разрушают[33], поэтому они не разрушаются ни в клетках, ни в плазме крови[34]. Морфолиновые олигонуклеотиды не активируют Toll-подобные рецепторы, а потому не запускают врождённый иммунный ответ, выражающийся, в частности, в образовании интерферона или в NF-κB-опосредованном воспалении[35].
До 18 % морфолиновых олигонуклеотидов вызывают не связанные с исходными мишенями фенотипы, в частности, смерть клеток центральной нервной системы и сомитов у зародышей данио-рерио[36]. Было показано, что большая часть этих эффектов связана с активацией p53-опосредованного апоптоза; их можно подавить, одновременно вводя экспериментальный морфолиновый олигонуклеотид и его анти-p53-аналог. Более того, вызванную морфолиновым олигонуклеотидом активацию p53-опосредованного апоптоза удалось повторить с использованием других антисмысловых структур, что говорит в пользу того, что p53-опосредованный апоптоз может быть вызван утратой белка-мишени и не зависит от типа олигонуклеотида, используемого для нокдауна[37].
Морфолиновые олигонуклеотиды следует использовать с осторожностью из-за их потенциальных эффектов вне белка-мишени. Для проверки того, является ли наблюдаемый фенотип морфанта результатом нокдауна задуманного гена или же он вызван взаимодействием с РНК, не являющейся непосредственной мишенью введенного олигонуклеотида, можно провести второй эксперимент. Этот эксперимент заключается в повторении фенотипа (фенокопировании) морфанта с использованием другого морфолинового олигонуклеотида, нацеленного на ту же мишень, но не перекрывающегося с первым олигонуклеотидом[31].
Доставка
Для того, чтобы морфолиновый олигонуклеотид мог подействовать, он должен попасть в цитозоль клетки и преодолеть клеточную мембрану. После попадания в цитозоль он начинает свободно диффундировать между цитозолем и ядром, как было показано в экспериментах, при которых введение морфолинового нуклеотида в цитозоль клетки изменяло сплайсинг в ядре[6]. Для доставки морфолинового олигонуклеотида в зародыши, клетки культуры или клетки взрослых животных используются различные методы.
Для доставки в зародыши, как правило, используют микроинъекции, причём введение олигонуклеотидов обычно осуществляют на стадии одной или нескольких клеток[38]. Альтернативный метод для доставки морфолиновых олигонуклеотидов в эмбрионы — электропорация, которая позволяет доставить их в ткани на более поздних стадиях развития зародыша[39].
Наиболее распространённые методы для доставки морфолиновых олигонуклеотидов в клетки культуры — использование эндопортерного пептида (англ. Endo-Porter peptide), который вызывает высвобождение морфолиновых олигонуклеотидов из эндосом[17]; система специальной доставки (англ. Special Delivery system), использующая гетеродуплекс ДНК с морфолиновым олигонуклеотидом и этоксилированный полиэтиленимин в качестве реагента доставки (больше коммерчески недоступна)[16]; электропорация[40]; метод соскоба-загрузки (англ. scrape loading)[41]. В 2015 году была описана транс-активирующая амфипатическая система доставки, основанная на ДНК, которая предназначена для удобной доставки незаряженных нуклеиновых кислот, имеющих поли(А)-хвост, таких как пептидо-нуклеиновые кислоты и морфолиновые олигонуклеотиды[42].
Доставка морфолиновых олигонуклеотидов в ткани взрослых животных затруднительна, хотя разработано несколько систем, позволяющих доставить в ткани немодифицированные морфолиновые олигонуклеотиды (в частности, в от природы слабые мышечные клетки при мышечной дистрофии Дюшена[43] или в клетки сосудистого эндотелия, подвергающиеся стрессу при ангиопластии[44]). Хотя морфолиновые олигонуклеотиды эффективно проходят через межклеточные промежутки в тканях, доставка их в клетки сложна. Системная доставка в клетки многих типов может быть осуществлена при помощи морфолиновых олигонуклеотидов, ковалентно связанных с проникающими в клетки пептидами (англ. Cell-penetrating peptide), и, в то время как токсичный эффект оказывают средние дозы связанных пептидов[45][46], в условиях in vivo удалось добиться эффективной доставки морфолиновых олигонуклеотидов при дозах пептидов ниже токсичных[9][47]. Октагуанидиновый дендример, прикреплённый к концу морфолинового олигонуклеотида (т. н. Vivo-Morpholino), обеспечивает его доставку из крови в цитозоль при систематическом введении во взрослого животного[48][49].
Медицинское значение
Морфолиновые олигонуклеотиды, способные к эффективной доставке (такие как связанные с пептидами морфолиновые олигонуклеотиды и Vivo-Morpholino), могут быть перспективными средствами для лечения вирусных и генетических заболеваний[50]. Например, Sarepta Therapeutics Inc. уже разрабатывает морфолиновые олигонуклеотиды — потенциальные лекарства под названием NeuGene. В настоящее время проходят клинические испытания морфолиновые олигонуклеотиды, которые могут использоваться для лечении миодистрофии Дюшенна у человека[51].
Примечания
- Аппель и др., 2013, с. 206.
- П. В. Золотухин, Ю. А. Лебедева, О. Н. Кузьминова, Е. К. Брюханова. Модификации и аналоги нуклеиновых кислот: инструменты современной молекулярной биологии // Валеология. — 2013. — № 2. — С. 27—33. — ISSN 2218-2268.
- Н. М. Белоногова. «Прямая» и «обратная» генетика. Генетика количественных признаков // Вавиловский журнал генетики и селекции. — 2014. — Т. 18, № 1. — С. 147—157.
- Т. В. Абрамова, В. Н. Сильников. Синтез и свойства модифицированных по углеводофосфатному остову аналогов олигонуклеотидов и миметиков нуклеиновых кислот // Усп. хим.. — 2011. — Т. 80, № 5. — С. 452—476.
- Summerton J. Morpholino antisense oligomers: the case for an RNase H-independent structural type. (англ.) // Biochimica et biophysica acta. — 1999. — Vol. 1489, no. 1. — P. 141—158. — PMID 10807004.
- Draper B. W., Morcos P. A., Kimmel C. B. Inhibition of zebrafish fgf8 pre-mRNA splicing with morpholino oligos: a quantifiable method for gene knockdown. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3. — P. 154—156. — PMID 11477696.
- Heasman J. Morpholino oligos: making sense of antisense? (англ.) // Developmental biology. — 2002. — Vol. 243, no. 2. — P. 209—214. — doi:10.1006/dbio.2001.0565. — PMID 11884031.
- Geller B. L. Antibacterial antisense. (англ.) // Current opinion in molecular therapeutics. — 2005. — Vol. 7, no. 2. — P. 109—113. — PMID 15844617.
- Deas T. S., Bennett C. J., Jones S. A., Tilgner M., Ren P., Behr M. J., Stein D. A., Iversen P. L., Kramer L. D., Bernard K. A., Shi P. Y. In vitro resistance selection and in vivo efficacy of morpholino oligomers against West Nile virus. (англ.) // Antimicrobial agents and chemotherapy. — 2007. — Vol. 51, no. 7. — P. 2470—2482. — doi:10.1128/AAC.00069-07. — PMID 17485503.
- McClorey G., Fall A. M., Moulton H. M., Iversen P. L., Rasko J. E., Ryan M., Fletcher S., Wilton S. D. Induced dystrophin exon skipping in human muscle explants. (англ.) // Neuromuscular disorders : NMD. — 2006. — Vol. 16, no. 9-10. — P. 583—590. — doi:10.1016/j.nmd.2006.05.017. — PMID 16919955.
- Summerton J., Weller D. Morpholino antisense oligomers: design, preparation, and properties. (англ.) // Antisense & nucleic acid drug development. — 1997. — Vol. 7, no. 3. — P. 187—195. — PMID 9212909.
- Nasevicius A., Ekker S. C. Effective targeted gene 'knockdown' in zebrafish. (англ.) // Nature genetics. — 2000. — Vol. 26, no. 2. — P. 216—220. — doi:10.1038/79951. — PMID 11017081.
- Heasman J., Kofron M., Wylie C. Beta-catenin signaling activity dissected in the early Xenopus embryo: a novel antisense approach. (англ.) // Developmental biology. — 2000. — Vol. 222, no. 1. — P. 124—134. — doi:10.1006/dbio.2000.9720. — PMID 10885751.
- Howard E. W., Newman L. A., Oleksyn D. W., Angerer R. C., Angerer L. M. SpKrl: a direct target of beta-catenin regulation required for endoderm differentiation in sea urchin embryos. (англ.) // Development (Cambridge, England). — 2001. — Vol. 128, no. 3. — P. 365—375. — PMID 11152635.
- Kos R., Reedy M. V., Johnson R. L., Erickson C. A. The winged-helix transcription factor FoxD3 is important for establishing the neural crest lineage and repressing melanogenesis in avian embryos. (англ.) // Development (Cambridge, England). — 2001. — Vol. 128, no. 8. — P. 1467—1479. — PMID 11262245.
- Morcos P. A. Achieving efficient delivery of morpholino oligos in cultured cells. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3. — P. 94—102. — PMID 11477682.
- Summerton J. E. Endo-Porter: a novel reagent for safe, effective delivery of substances into cells. (англ.) // Annals of the New York Academy of Sciences. — 2005. — Vol. 1058. — P. 62—75. — doi:10.1196/annals.1359.012. — PMID 16394126.
- Stancheva I., Collins A. L., Van den Veyver I. B., Zoghbi H., Meehan R. R. A mutant form of MeCP2 protein associated with human Rett syndrome cannot be displaced from methylated DNA by notch in Xenopus embryos. (англ.) // Molecular cell. — 2003. — Vol. 12, no. 2. — P. 425—435. — PMID 14536082.
- Bruno I. G., Jin W., Cote G. J. Correction of aberrant FGFR1 alternative RNA splicing through targeting of intronic regulatory elements. (англ.) // Human molecular genetics. — 2004. — Vol. 13, no. 20. — P. 2409—2420. — doi:10.1093/hmg/ddh272. — PMID 15333583.
- Vetrini F., Tammaro R., Bondanza S., Surace E. M., Auricchio A., De Luca M., Ballabio A., Marigo V. Aberrant splicing in the ocular albinism type 1 gene (OA1/GPR143) is corrected in vitro by morpholino antisense oligonucleotides. (англ.) // Human mutation. — 2006. — Vol. 27, no. 5. — P. 420—426. — doi:10.1002/humu.20303. — PMID 16550551.
- Morcos P. A. Achieving targeted and quantifiable alteration of mRNA splicing with Morpholino oligos. (англ.) // Biochemical and biophysical research communications. — 2007. — Vol. 358, no. 2. — P. 521—527. — doi:10.1016/j.bbrc.2007.04.172. — PMID 17493584.
- König H., Matter N., Bader R., Thiele W., Müller F. Splicing segregation: the minor spliceosome acts outside the nucleus and controls cell proliferation. (англ.) // Cell. — 2007. — Vol. 131, no. 4. — P. 718—729. — doi:10.1016/j.cell.2007.09.043. — PMID 18022366.
- Kloosterman W. P., Wienholds E., Ketting R. F., Plasterk R. H. Substrate requirements for let-7 function in the developing zebrafish embryo. (англ.) // Nucleic acids research. — 2004. — Vol. 32, no. 21. — P. 6284—6291. — doi:10.1093/nar/gkh968. — PMID 15585662.
- Flynt A. S., Li N., Thatcher E. J., Solnica-Krezel L., Patton J. G. Zebrafish miR-214 modulates Hedgehog signaling to specify muscle cell fate. (англ.) // Nature genetics. — 2007. — Vol. 39, no. 2. — P. 259—263. — doi:10.1038/ng1953. — PMID 17220889.
- Kloosterman W. P., Lagendijk A. K., Ketting R. F., Moulton J. D., Plasterk R. H. Targeted inhibition of miRNA maturation with morpholinos reveals a role for miR-375 in pancreatic islet development. (англ.) // Public Library of Science Biology. — 2007. — Vol. 5, no. 8. — P. e203. — doi:10.1371/journal.pbio.0050203. — PMID 17676975.
- Lagendijk A. K., Moulton J. D., Bakkers J. Revealing details: whole mount microRNA in situ hybridization protocol for zebrafish embryos and adult tissues. (англ.) // Biology open. — 2012. — Vol. 1, no. 6. — P. 566—569. — doi:10.1242/bio.2012810. — PMID 23213449.
- Yen L., Svendsen J., Lee J. S., Gray J. T., Magnier M., Baba T., D'Amato R. J., Mulligan R. C. Exogenous control of mammalian gene expression through modulation of RNA self-cleavage. (англ.) // Nature. — 2004. — Vol. 431, no. 7007. — P. 471—476. — doi:10.1038/nature02844. — PMID 15386015.
- Matter N., König H. Targeted 'knockdown' of spliceosome function in mammalian cells. (англ.) // Nucleic acids research. — 2005. — Vol. 33, no. 4. — P. e41. — doi:10.1093/nar/gni041. — PMID 15731334.
- Howard M. T., Gesteland R. F., Atkins J. F. Efficient stimulation of site-specific ribosome frameshifting by antisense oligonucleotides. (англ.) // RNA (New York, N.Y.). — 2004. — Vol. 10, no. 10. — P. 1653—1661. — doi:10.1261/rna.7810204. — PMID 15383681.
- Penn A. C., Balik A., Greger I. H. Steric antisense inhibition of AMPA receptor Q/R editing reveals tight coupling to intronic editing sites and splicing. (англ.) // Nucleic acids research. — 2013. — Vol. 41, no. 2. — P. 1113—1123. — doi:10.1093/nar/gks1044. — PMID 23172291.
- Bill B. R., Petzold A. M., Clark K. J., Schimmenti L. A., Ekker S. C. A primer for morpholino use in zebrafish. (англ.) // Zebrafish. — 2009. — Vol. 6, no. 1. — P. 69—77. — doi:10.1089/zeb.2008.0555. — PMID 19374550.
- Kamachi Y., Okuda Y., Kondoh H. Quantitative assessment of the knockdown efficiency of morpholino antisense oligonucleotides in zebrafish embryos using a luciferase assay. (англ.) // Genesis (New York, N.Y. : 2000). — 2008. — Vol. 46, no. 1. — P. 1—7. — doi:10.1002/dvg.20361. — PMID 18196596.
- Hudziak R. M., Barofsky E., Barofsky D. F., Weller D. L., Huang S. B., Weller D. D. Resistance of morpholino phosphorodiamidate oligomers to enzymatic degradation. (англ.) // Antisense & nucleic acid drug development. — 1996. — Vol. 6, no. 4. — P. 267—272. — PMID 9012862.
- Youngblood D. S., Hatlevig S. A., Hassinger J. N., Iversen P. L., Moulton H. M. Stability of cell-penetrating peptide-morpholino oligomer conjugates in human serum and in cells. (англ.) // Bioconjugate chemistry. — 2007. — Vol. 18, no. 1. — P. 50—60. — doi:10.1021/bc060138s. — PMID 17226957.
- Jon D. Moulton. A Brief Introduction to Morpholino Antisense. — 2011.
- Ekker S. C., Larson J. D. Morphant technology in model developmental systems. (англ.) // Genesis (New York, N.Y. : 2000). — 2001. — Vol. 30, no. 3. — P. 89—93. — PMID 11477681.
- Robu M. E., Larson J. D., Nasevicius A., Beiraghi S., Brenner C., Farber S. A., Ekker S. C. p53 activation by knockdown technologies. (англ.) // PLoS genetics. — 2007. — Vol. 3, no. 5. — P. e78. — doi:10.1371/journal.pgen.0030078. — PMID 17530925.
- Rosen J. N., Sweeney M. F., Mably J. D. Microinjection of zebrafish embryos to analyze gene function. (англ.) // Journal of visualized experiments : JoVE. — 2009. — No. 25. — doi:10.3791/1115. — PMID 19274045.
- Cerda G. A., Thomas J. E., Allende M. L., Karlstrom R. O., Palma V. Electroporation of DNA, RNA, and morpholinos into zebrafish embryos. (англ.) // Methods (San Diego, Calif.). — 2006. — Vol. 39, no. 3. — P. 207—211. — doi:10.1016/j.ymeth.2005.12.009. — PMID 16837210.
- Jubin R. Optimizing electroporation conditions for intracellular delivery of morpholino antisense oligonucleotides directed against the hepatitis C virus internal ribosome entry site. (англ.) // Methods in molecular medicine. — 2005. — Vol. 106. — P. 309—322. — PMID 15375324.
- Partridge M., Vincent A., Matthews P., Puma J., Stein D., Summerton J. A simple method for delivering morpholino antisense oligos into the cytoplasm of cells. (англ.) // Antisense & nucleic acid drug development. — 1996. — Vol. 6, no. 3. — P. 169—175. — PMID 8915501.
- H. V. Jain, D. Verthelyi and S. L. Beaucage. Amphipathic trans-acting phosphorothioate DNA elements mediate the delivery of uncharged nucleic acid sequences in mammalian cells // RSC Adv.. — 2015. — Vol. 5. — P. 65245—65254. — doi:10.1039/C5RA12038A.
- Fletcher S., Honeyman K., Fall A. M., Harding P. L., Johnsen R. D., Wilton S. D. Dystrophin expression in the mdx mouse after localised and systemic administration of a morpholino antisense oligonucleotide. (англ.) // The journal of gene medicine. — 2006. — Vol. 8, no. 2. — P. 207—216. — doi:10.1002/jgm.838. — PMID 16285002.
- Kipshidze N. N., Kim H. S., Iversen P., Yazdi H. A., Bhargava B., New G., Mehran R., Tio F., Haudenschild C., Dangas G., Stone G. W., Iyer S., Roubin G. S., Leon M. B., Moses J. W. Intramural coronary delivery of advanced antisense oligonucleotides reduces neointimal formation in the porcine stent restenosis model. (англ.) // Journal of the American College of Cardiology. — 2002. — Vol. 39, no. 10. — P. 1686—1691. — PMID 12020498.
- Abes S., Moulton H. M., Clair P., Prevot P., Youngblood D. S., Wu R. P., Iversen P. L., Lebleu B. Vectorization of morpholino oligomers by the (R-Ahx-R)4 peptide allows efficient splicing correction in the absence of endosomolytic agents. (англ.) // Journal of controlled release : official journal of the Controlled Release Society. — 2006. — Vol. 116, no. 3. — P. 304—313. — doi:10.1016/j.jconrel.2006.09.011. — PMID 17097177.
- Burrer R., Neuman B. W., Ting J. P., Stein D. A., Moulton H. M., Iversen P. L., Kuhn P., Buchmeier M. J. Antiviral effects of antisense morpholino oligomers in murine coronavirus infection models. (англ.) // Journal of virology. — 2007. — Vol. 81, no. 11. — P. 5637—5648. — doi:10.1128/JVI.02360-06. — PMID 17344287.
- Amantana A., Moulton H. M., Cate M. L., Reddy M. T., Whitehead T., Hassinger J. N., Youngblood D. S., Iversen P. L. Pharmacokinetics, biodistribution, stability and toxicity of a cell-penetrating peptide-morpholino oligomer conjugate. (англ.) // Bioconjugate chemistry. — 2007. — Vol. 18, no. 4. — P. 1325—1331. — doi:10.1021/bc070060v. — PMID 17583927.
- Li Y. F., Morcos P. A. Design and synthesis of dendritic molecular transporter that achieves efficient in vivo delivery of morpholino antisense oligo. (англ.) // Bioconjugate chemistry. — 2008. — Vol. 19, no. 7. — P. 1464—1470. — doi:10.1021/bc8001437. — PMID 18564870.
- Morcos P. A., Li Y., Jiang S. Vivo-Morpholinos: a non-peptide transporter delivers Morpholinos into a wide array of mouse tissues. (англ.) // BioTechniques. — 2008. — Vol. 45, no. 6. — P. 613—614. — PMID 19238792.
- Moulton J. D., Jiang S. Gene knockdowns in adult animals: PPMOs and vivo-morpholinos. (англ.) // Molecules (Basel, Switzerland). — 2009. — Vol. 14, no. 3. — P. 1304—1323. — doi:10.3390/molecules14031304. — PMID 19325525.
- Efficacy Study of AVI-4658 to Induce Dystrophin Expression in Selected Duchenne Muscular Dystrophy Patients.
Литература
- Б. Аппель и др. Нуклеиновые кислоты: от А до Я / под ред. С. Мюллер. — М.: БИНОМ. Лаборатория знаний, 2013. — С. 206. — 413 с. — ISBN 978-5-9963-0376-2.
- Subbotina E., Koganti S. R., Hodgson-Zingman D., Zingman L. V. Morpholino-driven gene editing: A new horizon for disease treatment and prevention. (англ.) // Clinical pharmacology and therapeutics. — 2015. — doi:10.1002/cpt.276. — PMID 26474085.
- Bhadra J., Pattanayak S., Sinha S. Synthesis of Morpholino Monomers, Chlorophosphoramidate Monomers, and Solid-Phase Synthesis of Short Morpholino Oligomers. (англ.) // Current protocols in nucleic acid chemistry / edited by Serge L. Beaucage ... [et al.]. — 2015. — Vol. 62. — P. 4—65. — doi:10.1002/0471142700.nc0465s62. — PMID 26380905.
- Moulton H. M. In vivo delivery of morpholino oligos by cell-penetrating peptides. (англ.) // Current pharmaceutical design. — 2013. — Vol. 19, no. 16. — P. 2963—2969. — PMID 23140456.
Ссылки
Типы нуклеиновых кислот | ||||
|---|---|---|---|---|
| Азотистые основания | ||||
| Нуклеозиды | ||||
| Нуклеотиды | ||||
| РНК | ||||
| ДНК |
| |||
| Аналоги |
| |||
| Типы векторов |
| |||
| ||||
