Вестерн-блот
Вестерн-блоттинг (вестерн-блот, белковый иммуноблот, англ. Western blot) — аналитический метод, используемый для определения в образце специфичных белков. На первом этапе используют электрофорез белков в полиакриламидном геле для разделения денатурированных полипептидов по длине (как правило, в присутствии SDS) или по трехмерной структуре белка (в нативном состоянии). Далее белки переносят на нитроцеллюлозную или PVDF-мембрану, затем детектируют с использованием антител, специфичных к заданному белку[1][2].
Существует множество коммерческих компаний, специализирующихся на производстве антител (как моноклональных, так и поликлональных) к десяткам тысяч различных белков[3].
Вестерн-блоттинг используется в молекулярной биологии, биохимии, генетике и в других естественно-научных дисциплинах.
Другие сходные методы используют антитела для определения белков в тканях и клетках посредством иммуноокрашивания и иммуноферментного анализа (ИФА, англ. ELISA).
Вестерн-блоттинг был разработан в лаборатории Джорджа Старка (англ. George Stark) в Стэнфорде. Название «вестерн-блот» технике было дано У. Нейлом Бурнеттом (англ. W. Neal Burnette)[4] и является игрой слов от названия «Саузерн-блоттинг», — методики определения ДНК, разработанной ранее Эдвином Саузерном. Аналогичный метод определения РНК называется нозерн-блоттингом, детекция посттрансляционных модификаций белков — истерн-блоттингом (англ. Eastern blotting).
Подготовка образца
Образец может быть взят из цельной ткани или из клеточной культуры. В большинстве случаев твёрдые ткани сначала измельчаются с использованием блендера (для образцов большого объёма), гомогенизатора (меньшие объёмы) или обработки ультразвуком. При этом бактерии, вирусы и другие компоненты окружающей среды также являются источником белков.
Для улучшения лизиса клеток и растворения белков могут быть применены различные детергенты, соли и буферы. Для предотвращения расщепления образцов их собственными ферментами часто добавляют ингибиторы протеаз и фосфатаз. Подготовка тканей часто выполняется при низких температурах, чтобы избежать денатурации белка.
Для разделения различных клеточных компартментов и органелл применяют комбинации биохимических и механических методик фракционирования, в том числе различные типы фильтрации и центрифугирования.
Гель-электрофорез
Белки разделяются при помощи электрофореза в полиакриламидном геле. Разделение белков можно производить по изоэлектрической точке (pI), молекулярной массе, электрическому заряду или по сочетанию этих параметров.
Наиболее распространённый способ разделения белков — электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (англ. SDS) по Лэммли.[5] SDS вызывает денатурацию белков и поддерживает их в денатурированном состоянии, для разрушения вторичных и третичных структур белков используют восстановители дисульфидных связей, например, дитиотреитол и меркаптоэтанол. Денатурированные полипептиды мигрируют в электрическом поле через акриламидный гель к аноду, при этом белки меньшего размера двигаются быстрее и, таким образом, разделяются в соответствии с молекулярной массой. Концентрация акриламида определяет разрешающую способность геля — чем выше концентрация акриламида, тем лучше разрешение низкомолекулярных белков. Низкая концентрация акриламида улучшает разрешающую способность для высокомолекулярных белков. Также возможно использование двумерного электрофореза (2-D). В таком случае разделение белков производят в двух направлениях — в соответствии с их изоэлектрической точкой в первом направлении и в соответствии с молекулярной массой — во втором.
Образцы на гель наносят в карманы. Как правило, одну из «дорожек» оставляют для маркеров молекулярной массы (смеси белков с известными массами). После подачи напряжения белки двигаются в электрическом поле с различной скоростью. Отличия в скорости продвижения — электрофоретической подвижности — приводит к разделению белков на полосы (англ. bands).
Перенос на мембрану
Чтобы сделать белки доступными для антител и дальнейшей детекции, их вместе с полоской геля переносят на мембрану, изготовленную из нитроцеллюлозы или поливинилиденфторида (англ. PVDF). Мембрана накладывается поверх геля, а поверх неё кладут стопку фильтровальной бумаги. Всю стопку помещают в буфер для переноса, который под действием капиллярных сил продвигается вверх по бумаге, унося с собой белки. Другой метод переноса белков называется электроблоттингом и использует электрический ток, который переносит белки из геля на мембрану. Белки перемещаются из геля на мембрану с сохранением своего расположения. В результате этого «промакивания» (от. англ. blotting) процесса белки удерживаются на тонком поверхностном слое мембраны для детекции. Оба варианта мембран используют из-за их свойства неспецифично связывать белки. Связывание белков основано как на гидрофобных взаимодействиях, так и на электростатических взаимодействиях между мембраной и белком. Нитроцеллюлозная мембрана дешевле PVDF, но гораздо более хрупкая и хуже выдерживает повторное нанесение меток.
Единообразие и общая эффективность переноса белков из геля на мембрану может быть проверена окрашиванием мембраны красителями Coomassie blue или Ponceau S. Coomassie наиболее распространённый из двух, а краситель Ponceau S обладает большей чувствительностью и лучше растворим в воде, что упрощает последующую отмывку и нанесение меток на мембрану.[6]
Блокирование
Как только выбрана мембрана за её способность связывать белки, выбраны антитела и целевой белок, должны быть приняты меры по исключению взаимодействия между мембраной и антителом, используемым для детекции целевого белка (ибо антитело само по себе белок). Блокирование неспецифичных связываний достигается помещением мембраны в разбавленный раствор белка — обычно бычий сывороточный альбумин или нежирное сухое молоко (оба недорогие), с небольшим процентом детергента типа Tween 20 или Triton X-100. Белок из разбавленного раствора прикрепляется к мембране во всех местах, где не прикрепился целевой белок. Поэтому при добавлении антител единственное свободное место на мембране, куда они могут прикрепиться, — это сайты связывания на специфичных целевых белках. Этот фоновый «шум» в окончательном продукте вестерн-блота приводит к чистым результатам и исключению ложно-положительных.
Детекция
Во время процесса детекции мембрана «метится» исследуемым белком с модифицированным антителом, которое связано с репортёрным ферментом, который выдерживается на соответствующей подложке, приводя к колориметрической реакции и давая цвет. В силу различных причин, детекция проводится в два этапа, хотя сейчас доступен одношаговый метод детекции для определённых областей применения.
Антитела вырабатывают, воздействуя на класс хозяйских или на культуру иммунных клеток некоторым белком (или его частью). Обычно это часть иммунного ответа, а здесь (в анализе) собранные антитела используются как специфичный и чувствительный инструмент детекции, который напрямую связывается с белком.
После блокирования разведённый раствор первичных антител (обычно между 0.5 и 5 мкг/мл) инкубируется с мембраной и слегка встряхивается. Обычно раствор состоит из буферного раствора соли с небольшим процентным содержанием детергента, иногда с сухим молоком или БСА. Раствор антител и мембрана могут быть вместе закрыты и инкубированы где угодно от 30 минут до оставления на ночь. Также они могут быть инкубированы при различных температурах, при повышенной температуре наблюдается лучшее связывание — и специфичное (целевого белка, «сигнал1») и неспецифичное («шум»).
После полоскания мембраны, служащего для удаления несвязавшихся первичных антител, мембрану выдерживают в других антителах, напрямую связывающихся с класс-специфическими участками первичных антител. Они известны как вторичные антитела и в соответствии с их целевыми свойствами, как правило называются по типу «anti-mouse», «anti-goat», и так далее. Антитела получают из животного источника (или животных — источников культуры гибридом); анти-мышиные вторичные антитела будут связываться с большинством первичных антител, полученных из мышей. Это создаёт некоторую экономию, позволяя отдельной лаборатории использовать один источник массового производства антител, и ведёт к намного более воспроизводимым результатам. Вторичные антитела обычно связывают с биотином или с репортёрным ферментом, таким как щелочная фосфатаза или пероксидаза хрена. Это значит, что несколько вторичных антител могут связываться с одним первичным и усиливать сигнал.
Наиболее распространённые, связанные с пероксидазой хрена вторичные антитела используются для разрезания хемилюминесцентного агента, и продукт реакции производит люминесцентное излучение пропорционально количеству белка. Лист светочувствительной фотографической плёнки помещается напротив мембраны и подвергается действию излучения реакции, создавая изображение полос антител на блоте. Более дешёвый, но менее чувствительный подход с использованием 4-хлоронафтольного окрашивания в смеси с 1 % перекисью водорода; реакция пероксидного радикала с 4-хлоронафтолом даёт темно-коричневое окрашивание, которое регистрируется без использования специальной фотографической плёнки.
Как и в методиках ELISPOT и ИФА, фермент может обеспечиваться молекулой субстрата, которая превращается ферментом в цветной продукт реакции, а он будет виден на мембране (смотри ниже, рисунок с синими бэндами).
Другой метод детекции вторичными антителами использует антитела со связанным флюорофором, который излучает в ближней инфракрасной области (NIR). Свет, излучаемый флюоресцентным красителем, постоянен и делает флюоресцентную детекцию более точным и чувствительным способом измерения разницы в сигнале, производимом белками, которые мечены антителами, на вестерн-блоте. Белки могут быть определены количественно, так как сигнал рассчитывается как разница (излучения) по отношению ко всем белкам на мембране, измеренном в покое, к (излучению) хемилюминесценции, которая измеряется в динамике.[7]
Третий альтернативный метод использует радиоактивную метку вместо фермента, связанного с вторичным антителом, такую как меченный антитело-связывающий белок типа Белка А Staphylococcus с радиоактивным изотопом йода. Другие методы безопаснее, быстрее и дешевле, поэтому радиоактивная детекция используется редко.
Исторически процесс нанесения меток проводится в два этапа, потому что относительно проще произвести первичные и вторичные антитела в раздельных процессах. Это даёт исследователям и компаниям огромные преимущества в плане гибкости, и добавляет шаг амплификации в процесс детекции. Учитывая появление высоко-пропускного анализа белка и низкий порог обнаружения, все же наблюдается интерес к развитию системы-в-один-шаг для нанесения меток, которая позволяет процессу проходить быстрее и с меньшими затратами. Она (система-в-один-шаг) требует антител-меток, которые распознавали бы исследуемый белок и одновременно несли маркер для детекции — метки, наиболее доступные для известных «белковых хвостов». Сначала метки инкубируют с мембраной в стиле метода-в-два-шага с первичными антителами, а затем они готовы для прямой детекции после серии промывок.
Анализ
После отмывки несвязавшихся меток вестерн-блот готов к детекции зондов, связавшихся с целевым белком. На практике, не во всех вестернах обнаруживают белки лишь по одному бэнду на мембране. Приблизительный размер вычисляют, сравнивая окрашенные бэнды с маркерами молекулярной массы, добавленными при электрофорезе. Процесс повторят с структурными белками, такими как актин или тубулин, которые не меняют между экспериментами. Количество целевого белка зависит от количества контрольного структурного белка между группами. Этот приём обеспечивает коррекцию количества общего белка на мембране в случае ошибки или неполного переноса.
Колориметрическая детекция
Метод колориметрической детекции основан на инкубации вестерн-блота с субстратом, который реагирует с репортёрным ферментом (таким как пероксидаза хрена, англ. horseradish peroxidase), «сидящем» на вторичном антителе. Растворимый краситель переходит в нерастворимую форму другого цвета, осаждаясь рядом с ферментом и окрашивая мембрану. Рост пятна ограничивается смыванием растворимого красителя. Уровень количества белка оценивается денситометрически по интенсивности окрашивания или спектрофотометрически.
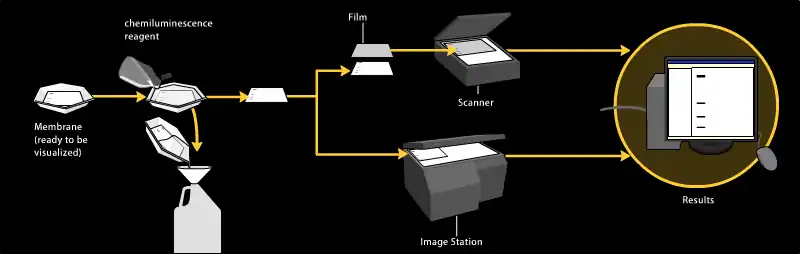
Хемилюминесцентная детекция
Метод хемилюминесцентной детекции основывается на инкубации нитроцеллюлозной мембраны с субстратом, который люминесцирует после взаимодействия с репортёром вторичного антитела. Свет регистрируется фотоплёнкой или CCD-камерой, которая производит цифровую съёмку вестерн-блота. Изображение анализируется денситометрически, оценивая относительное количество окрашенного белка и даёт количественный результат в единицах оптической плотности. Новое программное обеспечение позволяет провести дальнейший анализ данных, например, определить молекулярный вес, если использовался соответствующий стандарт.
Радиоактивная детекция
Радиоактивные метки не нуждаются в ферментных субстратах, а позволяют помещать медицинскую радиографическую плёнку напротив вестерн-блота, давая ей (плёнке) возможность взаимодействовать с метками и создавая тёмные участки, которые соответствуют полосам исследуемого белка (на изображении справа). Востребованность методов радиоактивной детекции снижается из-за их дороговизны, высокого риска для здоровья и безопасности и альтернатив, предоставляемых ECL.
Флюоресцентная детекция
Флюоресцентные метки возбуждаются светом и излучают более длинноволновый свет, регистрируемый фотосенсорами, такими как CCD-камера, снабжённая соответствующими фильтрами эмиссии. Камера делает цифровой снимок вестерн-блота, позволяя проводить дальнейший анализ полученных данных, такой как анализ молекулярного веса и количественный вестерн-блот-анализ.
Протоколы
- Western Blotting Protocol from Protocolmonkey
- Modified Western blotting protocols from Biotechniques
- Dr. Mark Barton Frank Lab protocol
- Kamps’s Western Blotting Protocol
- https://web.archive.org/web/20090906223659/http://www.currentprotocols.com/protocol/et0801
- https://web.archive.org/web/20100420005952/http://www.natureprotocols.com/2006/09/15/western_blot_analysis_of_subce.php
- http://www.prosci-inc.com/Western-Blot-Protocol.html
Примечания
- Towbin H., Staehelin T., Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1979. — Vol. 76, no. 9. — P. 4350—4354. — doi:10.1073/pnas.76.9.4350. — PMID 388439.
- Renart J., Reiser J., Stark G. R. Transfer of proteins from gels to diazobenzyloxymethyl-paper and detection with antisera: a method for studying antibody specificity and antigen structure. (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1979. — Vol. 76, no. 7. — P. 3116—3120. — doi:10.1073/pnas.76.7.3116. — PMID 91164.
- Western blot antibody (недоступная ссылка). exactantigen.com. Дата обращения: 29 января 2009. Архивировано 17 декабря 2009 года.
- W. Neil Burnette. 'Western blotting': electrophoretic transfer of proteins from sodium dodecyl sulfate — polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A (англ.) // Analytical Biochemistry : journal. — United States: Academic Press, 1981. — April (vol. 112, no. 2). — P. 195—203. — ISSN 0003-2697. — doi:10.1016/0003-2697(81)90281-5. — PMID 6266278. Архивировано 14 мая 2008 года.
- U. K. Laemmli. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature,1970; V.227, P.680 — 685
- A guide to methods in the biomedical … — Google Books
- Ambroz K., (2006). «Improving quantification accuracy for western blots» Image Analysis09/2006.アーカイブされたコピー. Дата обращения: 16 июля 2009. Архивировано 29 августа 2008 года.