Крокодилы
Крокоди́лы (лат. Crocodilia) — отряд вторично-водных животных класса пресмыкающихся из клады эузухий, которая в свою очередь вместе со множеством промежуточных клад относится к кладе круротарзы или псевдозухии. Из ныне живущих организмов ближайшие родственники крокодилов — птицы, потомки сестринской ветви архозавров. Хотя словом «крокодил» правильнее всего обозначать представителей семейства настоящих крокодилов, в целом этот термин употребим для обозначения любых представителей отряда, к которому также относятся аллигаторовые (аллигаторы и кайманы), гавиаловые и некоторые базальные его представители. Нередко «крокодилами» называют любых крокодиломорфов (Crocodylomorpha), что является ошибкой на фоне современной систематики данной группы рептилий. В современном объёме данной систематической группы, крокодилы впервые появились 83,5 млн лет назад в позднем меловом периоде. Современный отряд включает в себя не менее 15 видов настоящих крокодилов (семейство Crocodylidae), 8 видов аллигаторовых (семейство Alligatoridae) и 2 вида гавиаловых (семейство Gavialidae, иногда упраздняемое до подсемейства в составе семейства настоящих крокодилов[1][2]).
| Крокодилы | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Подкласс: Клада: Инфракласс: Клада: Клада: Клада: Клада: Надотряд: Клада: Отряд: Крокодилы |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Crocodilia Owen, 1842 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Современный ареал | |||||||||||||||||||||||||||||||||||||||||
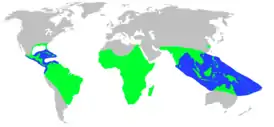 |
|||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 83,5 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
.jpg.webp)

_in_Florida.jpg.webp)
.jpg.webp)
Крокодилы имеют крупное, плотное, ящероподобное тело, вытянутую и приплюснутую морду, сжатые с боков хвосты, и глаза, уши и ноздри, располагающиеся на верхней части головы. Они хорошо плавают и могут достаточно быстро передвигаться по земле. Их кожа очень толстая и покрыта непересекающимися чешуями. Большинство современных крокодилов имеют сложный зубной ряд из зубов, отличающихся по формам, размерам и функциям (гетеродонтизм), а также объёмные челюстные мышцы, обеспечивающие большую силу укуса. Но некоторые виды имеют отличительные краниальные адаптации в силу особенностей питания: наглядным примером являются узкие челюсти и практически гомодонтная зубная система гавиалов.
Крокодилы имеют четырёхкамерное сердце с полной межжелудочковой перегородкой и, подобно птицам, однонаправленный цикл системы воздушного потока в лёгких. Однако, при этом крокодилы являются холоднокровными животными и их кровеносная система сохраняет возможность смешивания артериальной и венозной крови за счет наличия паницциева отверстия между левой и правой дугами аорты. В настоящее время считается, что холоднокровность является вторично приобретённым признаком этой группы, поскольку предки крокодилов, как и другие развитые архозавры, имели повышенный в сравнении с современными рептилиями уровень обмена веществ и скорее всего были теплокровными. Таким образом, такие признаки, как способность держать конечности относительно прямо под туловищем, четырёхкамерное сердце и более развитая, чем у большинства других современных пресмыкающихся, дыхательная система, являются не прогрессивными признаками крокодилов, а упрощением организации относительно предкового состояния.
Крокодилы встречаются в основном в тропических низинах, но аллигаторы обитают в более холодных регионах на юго-востоке США и в реке Янцзы в Китае. Все нынешние крокодилы — специализированные полуводные хищники, использующие в пищу широкий спектр водных, околоводных и приходящих на водопой животных. Некоторые виды, такие как гангский гавиал, имеют довольно узкий спектр питания. Крокодилы, как правило, одиночные и территориальные животные, хотя многие виды в целом терпимы к своим сородичам. Во время размножения доминирующие самцы всегда придерживаются своей территории и как правило являются полигамными по отношению к самкам. Самки откладывают яйца в ямах или холмиках, и некоторое время заботятся о своих детёнышах. Крокодилы, как известно, нападают на людей и являются наиболее опасными для человека крупными хищниками. Наибольшее количество нападений происходит со стороны нильского крокодила. За ним следует гребнистый и болотный крокодилы. В Новом Свете за наибольшее число нападений ответственны острорылый крокодил и чёрный кайман. Крокодилы почитались везде, где они обитают. Художественные и литературные представления о крокодилах появились в культурах по всему миру, начиная по меньшей мере со времён Древнего Египта.
Название
Слово «крокодил» происходит, в конечном счете, из древнегреческого языка. Там словом κροκόδειλος назвали и крокодилов, и ящериц (хотя для ящерицы было и другое слово — σαῦρος)[3]. Κροκόδειλος, вероятно, образовано от κρόκη δεῖλος, «галечный червь», — из-за сходства чешуи крокодилов и ящериц с галькой.
На протяжении десятилетий для научного наименования отряда использовались взаимозаменяемые термины Crocodilia и Crocodylia, пока К. П. Шмидт не предложил новое наименования Loricata[4]. В дальнейшем было принято первоначально предложенное для этой группы название Crocodilia[5][6].
До 2003 года отряд Crocodilia был группой, которая охватывала не только современных крокодилов и их ближайших вымерших родственников, но и приходящихся им достаточно дальними родственниками крокодилоподобных архозавров. В дальнейшем был введен надотряд Crocodylomorpha, в которого входит в том числе Crocodilia, в свою очередь включающая в себя только современных крокодилов и их ближайших предков. Это разделение было очень важно для палеонтологов, изучающих эволюцию крокодилов[7].
Древнерусское слово «крокодил» («коркодил»), транслитерирующее греческое слово κροκόδειλος, фиксируется с XII века[8]. Современное же русское слово, согласно этимологическому словарю Фасмера, заимствовано через нем. Krokodil в XVI веке.

Особенности строения
Длина большинства крокодилов 2—5,5 м. Наиболее крупный вид — гребнистый крокодил, достигает длины до 6,3 (потенциально до 7) метров. Среди наиболее мелких — гладколобый кайман Кювье и тупорылый крокодил, имеют максимальную зарегистрированную длину в 2,1 м и 1,9 м соответственно. Их внешность демонстрирует адаптацию к обитанию в водной среде: голова плоская, с длинным рылом (тем самым снижая сопротивление воды при развороте); туловище приплюснутое; хвост мощный, сжатый с боков; ноги довольно короткие, но сильные. На передних конечностях — 5 пальцев, на задних — 4 (нет мизинца), соединённых небольшой перепонкой. Глаза с вертикально-щелевидным зрачком расположены на верхней части головы, так что животное может выглядывать из воды, выставляя наружу только ноздри и глаза.
Разница между крокодилами и аллигаторами
Несмотря на внешнюю схожесть, настоящие крокодилы и аллигаторовые приходятся дальними родственниками и имеют множество анатомических и даже физиологических различий[9]. Исследователи отмечают различия в форме стопы[10], строении пояса передних конечностей и плечевой кости[11], мускулатуре конечностей и её аллометрии, физиологии упражнений[12] и кинематике движений[13], а также пропорциях конечностей или позвонков[14][15][16]. Аллигаторовые, в отличие от настоящих крокодилов, не способны на галоп[9]. Аллигаторы имеют относительно более короткие остистые отростки позвонков, короткую и широкую плечевую кость и коракоид, поддерживающие менее дифференцированную мускулатуру передних конечностей. Крестцовые позвонки, кости таза и задние конечности у крокодилов как правило длиннее, чем у аллигаторов[16]. У крокодилов более развит солевой обмен. Излишек солей выводится из организма крокодила с помощью специальных солевых желез, расположенных на языке (лингвальные железы), а также, в меньшей степени, с выделениями Гардеровых желез («крокодиловы слёзы»)[17]. У аллигаторов лингвальные железы представляют рудименты слюнных желез и выделяют только небольшое количество слизи. Поэтому крокодилы могут жить в солёной воде, а аллигаторы, за редкими исключениями, живут только в пресной. Чувствительные рецепторы, визуально различающиеся как небольшие точки, покрывают всё тело крокодилов, но только морду у аллигаторов. Ротовая полость аллигаторов не кератинизированна и таким образом намного нежнее, чем у крокодилов[2]. Недавно было показано, что аллигаторовые, в отличие от настоящих крокодилов, по крайней мере в некоторых случаях способны включать растительную пищу в свой рацион[18][19].
Визуально аллигаторовых от настоящих крокодилов легче всего отличать по строению челюстей. Когда челюсти у крокодила сомкнуты, то видны его 64—68 (или же, 68-72 у австралийского узкорылого крокодила[20], 60-64 у тупорылых крокодилов[21] и 64-70 у африканского узкорылого крокодила[22]) относительно длинных и острых зубов, особенно — увеличенный четвёртый зуб на нижней челюсти и пятый зуб на верхнечелюстной кости[23]. У аллигаторов зубы более многочисленны — их количество варьирует от 72 до 76 у китайского аллигатора[24] и черного каймана[25] до 84—86 у Апапорисского крокодилового каймана[26]. У представителей рода Alligator, а также черного и широконосого кайманов, зубы значительно мельче и имеют более притупленные коронки, чем у большинства настоящих крокодилов. У всех аллигаторовых четвёртый зуб нижней челюсти меньших размеров и верхняя челюсть полностью закрывает его. Также их можно отличить по форме морды: у настоящих крокодилов морда как правило острая, V-образная, а у аллигаторовых (кроме Апапорисского крокодилового каймана) — тупая, U-образная[2]. Однако, идентификация по строению челюстей может быть неприменима к старым животным с деформированными зубами и как правило очень массивными, вне зависимости от вида (за исключением более рыбоядных форм), челюстями.
 |
 |
Кожный покров
Как и у всех высших позвоночных, кожа крокодилов состоит из двух основных слоев — эпидермиса и дермы.
У крокодилов эпидермис составляет 12—14 % от всей кожи, при этом в нём располагается от 82 до 98 % пигментов, определяющих окраску кожи. Самый верхний слой эпидермиса — роговой слой, состоящий из мертвых клеток. В роговом слое крокодилов отсутствует кутикулярный слой, что исключает возможность линьки, характерной для большинства пресмыкающихся: их кожа обновляется постоянно, по мере изнашивания. Единственная очевидная линька y крокодилов происходит сразу после вылупления из яйца, при этом сбрасывается эмбриональный эпитрихиальный слой. На теле крокодилов располагаются роговые щитки, являющиеся, как и кости, производными эпидермиса.
.jpg.webp)
Дерма образована двумя слоями — губчатым и компактным. У крокодилов оба этих слоя толстые и пронизаны большим количеством соединительнотканных волокон, которые расположены слоями. Число слоев c возрастом увеличивается. Ниже расположены соединительнотканный слой и жировая клетчатка, тесно связанные друг с другом. B дерме крокодилов имеются зоны окостенения, образующие так называемые остеодермы — костные пластины примерно овальной формы, вытянутые в продольном направлении, иногда c продольным ребром. Они располагаются под роговыми щитками. Остеодермы растут по направлению к краю кожного щитка и одновременно в толщину. Они или отделены друг от друга (в большинстве случаев), или соприкасаются краями, не образуя швов, но между продольными рядами спинных щитков могут образовываться шовные соединения. Остеодермы по структуре кости близки к губчатой кости, имеют вертикальные углубления и ветвящиеся каналы, связанные c внутренними полостями. Эта стpуктуpа каналов и полостей снабжена кровеносными сосудами, имеет нервные клетки, a также содержит соединительную ткань и пигментные клетки. У всех видов крокодилов наиболее явно остеодермы выражены в дорсальных щитках, однако у некоторых видов они имеются в латеральных или даже вентральных щитках. Щитки служат защитой для мелких крокодилов, часто прикрывая их уязвимые места. У крупных же они в основном увеличивающий площадь поверхности тела при баскинге, а также способны накапливать минеральные вещества и даже небольшое количество выделяемого мышцами лактата. Некоторые остеодермы формируются по мере роста крокодила, и у взрослых животных их количество возрастает — к примеру, y взрослых крокодилов могут окостеневать щитки верхнего века.
Кожные железы
Кожные железы являются производным кожи. Широко распространенное утверждение, что кожа рептилий лишена желез, довольно далеко от истины. У крокодилов выделяют три группы желез: дорсальные, мандибулярные и клоакальные. Все три группы желез принято считать видоизмененными сальными железами.
Дорсальные железы располагаются под передними наружными углами второго ряда спинных щитков по обе стороны от серединной линии и могут быть обнаружены на всем протяжении спины от лопаток до основания хвоста. Эти железы представляют собой впячивания эпидермиса, возникающие на средних стадиях эмбриогенеза, их стенки выстланы однослойным базально-клеточным эпителием, продуцирующим масляно-жировой секрет. Эти выделения смазывают кожу между щитками, придавая ей эластичность. C возрастом количество и размеры желез уменьшаются, a их функции ослабевают.
Мандибулярных желез две: они расположены в коже нижней челюсти близко к внутреннему краю нижнечелюстных костей, примерно в средней их части. Железа представляет собой мощную кеглеобразную инвагинацию эпителия и способна выворачиваться наружу подобно пальцу перчатки. Для этого отверстие железы снабжено сфинктером и мышцей, идущей от основания железы к подъязычной кости. Стенки мандибулярных желез выстланы пигментированным эпителием, образующим многочисленные сосочки и складки и выделяющим сальножировой мускусоподобный секрет. Он служит для запаховых меток и опознавания партнёров.
Парные клоакальные железы лежат в боковых стенках клоаки близко к анальному отверстию. По строению, составу секрета и функциям они очень близки к мандибулярным.
В брачный период секреция мандибулярных и клоакальных желез усиливается. Выделяемый ими секрет коричневого цвета, имеет сильный мускусный запах и, вероятно, служит крокодилам разных полов помощником для нахождения друг друга.
Железы внутренней секреции
В отличие от большинства высших позвоночных, y крокодилов нет эпифиза и париетального органа. Причем следов их закладки не наблюдается даже на эмбриональных стадиях. Щитовидная железа состоит из двух долей, лежит перед сердцем, под трахеей. Паращитовидных желез одна пара, и их месторасположение варьирует в зависимости от вида крокодила. Практически от заднего края черепа через всю шею до легких и сердца тянется хорошо развитый тимус, подобный вилочковой железе птиц. Поджелудочная железа прилегает к первой петле тонкого кишечника. Надпочечники расположены дорсальнее почек и вытянуты вдоль них.
Костная система
Скелет крокодилов, как вымерших, так и современных, отличается рядом четких признаков.
Череп

Череп у крокодилов очень массивный и укреплен на всем протяжении, диапсидного типа — с двумя височными дугами; подобное строение черепа имеют лепидозавроморфы, динозавры, птицы и современные клювоголовые. Пасть укреплена более обширным, чем у млекопитающих, вторичным костным небом, неподвижно сочлененным c черепной коробкой. Это обеспечивает отделение носовых проходов от ротовой полости, что особенно важно для водных форм, однако первично данное укрепление связанно c повышением механической стойкости черепа, испытывающим большие перегрузки при сильном укусе и умерщвлении жертвы. Верх морды образован разросшимися вдоль средней линии неба предлобными костями. Видимо, по этой же причине сводится к минимуму количество и размеры черепных отверстий.
Череп крокодилов характерезуется плоской, низкой крышей, утратой заднелобной и заднетеменной костей и эпиптеригоида, слитыми по средней линии лобными и теменными костями. Свойственна сложная пневматизация почти всех костей черепа: это две системы воздухоносных каналов. Одна связана c полостью среднего уха и развилась из евстахиевых труб, a вторая — c носовыми проходами. Это придает легкость массивному черепу без утраты прочности. Однако точного объяснения такой системе пневматизации пока не предложено.
Посткраниальный скелет
Позвоночник крокодилов насчитывает 60-70 позвонков: 8-9 шейных, 10-11 грудных, 4-5 поясничных (всего — 24 предкрестцовых), 2 крестцовых и 32-42 хвостовых. Между последним грудным и первым поясничным позвонками нет четких различий, поэтому общее количество грудных и поясничных позвонков определяется примерно. Тела всех позвонков, кроме атланта, второго крестцового и первого хвостового процельные, как y ящериц и змей. Грудной и поясничный отделы позвоночного столба имеют высокую степень подвижности в латеральном и дорсовентральном направлении[27]. Важной особенностью скелета крокодилов становится двояковыпуклый первый хвостовой позвонок, который выполняет функцию шарового сустава. Это обеспечивает высокую подвижность хвоста как основного локомоторного органа в воде.
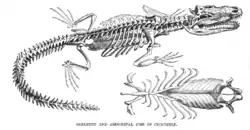
Первые девять позвонков имеют ребра, не достигающие грудины — они образуют шею. Шейные ребра, кроме связанных c атлантом и эпистрофеем, имеют широко разделенные головки, при этом дорсальная (направленная к спине) крепится к поперечному отростку, a основная — к парапофизу в нижней части тела позвонка. Таким образом, образуется система «арок», внутри которых проходит шейная артерия. Туловищных ребер, как правило, десять-тринадцать пар. Ребра задних туловищных позвонков постепенно укорачиваются и сливаются c поперечным отростком. У гавиалов нет последних трех, a y крокодилов и аллигаторов — четырёх-пяти туловищных ребер. Имеются брюшные ребра, а также грудина — сложная хрящевая структура, соединяющая концы ребер, основания коракоидов и межключицу. Так называемые крестцовые ребра — это крупные кости, которые соединяются суставами с поперечными отростками крестцовых позвонков. Не существует единого мнения, являются ли они истинными ребрами или утолщенными поперечными отростками. Однако, они не срастаются с позвоночником, и по этой причине их обычно называют рёбрами.
Передние и задние конечности крокодилов морфологически сходны с конечностями млекопитающих и крепятся к поясам конечностей аналогичным образом. Бедренная кость длиннее плечевой, в результате чего задняя часть тела располагается выше передней. На передних конечностях по пять пальцев, внутренние три из которых снабжены когтями. Задние конечности имеют по четыре пальца, на внутренних трех из которых также есть когти. В поясе передних конечностей крокодилов утрачена ключица, a идущие в спинно-брюшном направлении коракоиды удлинены и y современных видов равны длине лопатки. Эти признаки свидетельствуют o более эффективной работе мышц, поднимающих тело и отводящих конечность назад. Тазовый пояс состоит из короткой подвздошной, широкой лонной и короткой широкой седалищной костей. Для пояса задних конечностей характерно, что два крестцовых позвонка не срастаются в крестец, a в образовании вертлужной впадины не участвуют лобковые кости. В скелете задних конечностей крокодилов особенно важен хорошо развитый пяточный бугор, который представляет собой мощный рычаг для сгибания стопы. Это обусловлено тем, что крокодилы при передвижении по суше могут ходить, не опуская туловища на землю, что гораздо ближе к способу передвижения млекопитающих и наземных архозавров.
Мышечная система
Мышечная масса обычно составляет более 50 % от массы тела крокодила, оставляя небольшое пространство для других внутренних органов[28]. В скелетной мускулатуре абсолютно преобладают быстрые мышечные волокна.
Поскольку крокодилы имеют медленный обмен веществ и полагаются в первую очередь на анаэробный метаболизм, при повышенных физических нагрузках в их организме накапливается большее количество молочной кислоты. Лактат, образующийся в чрезмерно-больших количествах, потенциально может даже привести к лактатацидозу и тем самым представлять угрозу для жизни животного. Однако, крокодилы являются чемпионами среди всех позвоночных в способности накопить относительное и абсолютное количество молочной кислоты в крови и мышцах без значительного вреда для себя: мышцы крокодилов сами по себе крайне устойчивы к падениям значения pH, а часть продуктов анаэробного метаболизма откладывается в остеодермах и костях черепа. У молодого гребнистого крокодила массой 32 кг было зафиксировано падение pH крови от нормальных 7,4 до 6,42, которое он благополучно пережил[28].
При поимке крупные крокодилы способны активно сопротивляться в течение нескольких часов, в то время как особи весом менее 200 кг выдыхаются менее чем за 40 минут[28]. Это объясняется тем, что крупные крокодилы более устойчивы к накоплению молочной кислоты и имеют более развитый аэробный метаболизм за счет аллометрического увеличения объёма легких. С другой стороны, после полного выматывания у крупного крокодила может повышаться вероятность гибели от необратимых метаболических нарушений[2][28]. Поскольку крокодилы имеют низкий уровень обмена веществ и генерируют меньшее количество энергии, чем теплокровные животные, при длительных нагрузках их движения становятся не столь интенсивны.
Сила укуса
Аллигаторы и крокодилы обладают самым мощным укусом из всех животных существующих на планете. В исследовании, опубликованном в 2003 году, укус 272-килограммового миссисипского аллигатора был измерен в 9452 Н[29]. В исследовании 2012 года большее число в 16414 Н было получено от 531-килограммового гребнистого крокодила, содержащегося в неволе. Сила укуса 1308-килограммового гребнистого крокодила при этом была рассчитана в 34424 Н. Это исследование также показало отсутствие четкой корреляции между силой укуса, формой морды и рационом крокодила: лишь очень узкие челюсти гавиалов генерируют примерно вдвое меньшую силу, чем челюсти остальных крокодилов такого же веса. Их челюсти приспособлены скорее для быстрого, чем сильного захвата жертвы[30]. Сила укуса вымершего крокодила дейнозуха могла превышать 356450,6 Н[31].
Столь высокая сила укуса крокодилов обуславливается их уникальной краниальной анатомией: в черепе имеется очень много пространства для крепления челюстных мышц, но при этом они настолько объемны, что все равно легко различаются снаружи как заметные выпуклости по бокам у основания черепа. Эти мышцы невероятно плотные, они почти такие же твердые на ощупь, как кость. Другой примечательной характеристикой является то, что большая часть челюстных мышц крокодилов создана только для мощного закрытия пасти, в то время как мышцы, служащие для разжатия челюстей, очень маленькие и слабые. Пасть взрослого крокодила может быть надежно закрыта несколькими слоями скотча[32].
Чувствительные рецепторы, располагающиеся у ротовой полости крокодила, выступают в качестве механорецепторов и позволяют очень точно регулировать силу сжатия челюстей, что позволяет осуществлять ими такие аккуратные манипуляции, как перенос новорожденных детенышей в пасти, и эффективно контролировать захваченную челюстями жертву.
Органы дыхания
Носоглоточный проход отделён от ротовой полости вторичным костным нёбом. Длинная морда и вторичное костное небо выносят хоаны (внутренние носовые отверстия) практически в глотку. Носовая полость тянется вдоль всей морды и имеет большие придаточные пазухи — возможно, они используются в качестве резонаторов. Ротовая полость отделена от глотки плотно замыкающимся клапаном (нёбной завеской). Такое строение позволяет крокодилам манипулировать добычей или предметами под водой c помощью пасти и при этом дышать, выставив на поверхность кончик морды. При этом воду в дыхательное горло не пропускает нёбная завеска, а воздух проходит в трахею по носоглоточному ходу позади неё. Ноздри, расположенные на конце морды на вершине носового бугра, снабжены клапанами, закрывающимися при погружении.
Ритм дыхания в норме состоит из двух-трех дыхательных циклов, за которыми следует пауза от полминуты до пяти минут. В отличие от птиц и млекопитающих, безусловным сигналом к вдоху является не повышение концентрации углекислого газа в крови, a снижение концентрации кислорода. Максимальный срок пребывания крокодилов под водой без дыхания специфичен для разных видов и в норме колеблется от тридцати минут в активном состоянии, до нескольких часов при пониженной активности.
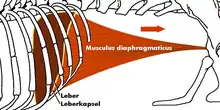
Гортань лежит на широком щитообразном теле подъязычной кости и соединена c ней связкой. Прямая трахея делится на два бронха, каждый из которых входит в легкие в центре на нижней стороне. Легкие крупные, устроены сложно, вмещают большой запас воздуха; с помощью лёгких крокодил способен регулировать плавучесть. Часть мышц, окружающих лёгкие, используется для перемещения воздуха в лёгких, а вместе с ним и центра плавучести[33]. Брюшная полость крокодилов отделена от грудной клетки диафрагмой. Однако эта диафрагма отличается от диафрагмы млекопитающих — у крокодилов она образована соединительнотканным и мышечным сращением между желудком, печенью и плеврой легкого. Приводится в действие с помощью печёночного насоса, в котором печень и свободно соединённые с тазом лобковые кости двигаются в переднезаднем направлении, меняя центр тяжести тела, что также участвует в регуляции плавучести и позволяет крокодилу принимать более удобное положение во время движения в воде и когда он охотится, прячась под водой.
Кровеносная система
Кровеносная система крокодилов отличается от таковой у других современных рептилий и птиц и считается наиболее сложной среди всех позвоночных.
Сердце четырёхкамерное: имеет два предсердия и два желудочка, полностью разделённых перегородкой. Однако имеется механизм управляемого смешивания артериальной крови с венозной. В отличие от млекопитающих и птиц, у крокодилов сохраняется обе дуги аорты — в системном круге кровообращения имеется «дополнительная» левая дуга аорты, берущая начало в правом желудочке (то есть там же, где и легочная артерия) за счет специфичного расположения межжелудочковой перегородки. При этом левая и правая дуги аорты на выходе из сердца очень плотно прилегают друг к другу, в месте их перекреста имеется анастомоз («паницциево отверстие»), и кровь может поступать из одной дуги в другую[34].
В нормальном режиме работы сердца часть артериальной крови попадает в открытое паниццево отверстие и поступает из правой дуги аорты в левую дугу аорты. Это наиболее выражено во время диастолы. Артериальная кровь также может перемещаться между обеими дугами аорты через анастомоз позади сердца. Давление в правой дуге аорты выше, чем в легочной артерии, и поэтому венозная кровь из правого желудочка не может преодолеть клапан, находящийся на пути из правого желудочка в левую дугу аорты. То есть, когда крокодил активен и не находится под водой, обе дуги аорты несут артериальную кровь и никакого смешивания артериальной и венозной крови не происходит. В этом случае сердца крокодилов работает также, как и сердца млекопитающих и птиц. Однако, за счёт наличия специального клапана, который может плотно перекрывать легочную артерию, крокодилы обладают дополнительным механизмом, позволяющим искусственно нагнетать бедную кислородом венозную кровь из правого желудочка в левую дугу аорты, что приводит к смешиванию артериальной и венозной крови. При перекрытии легочной артерии зубцовым клапаном, насыщенная углекислым газом кровь течет справа налево — из правого желудочка в левую дугу аорты, на фоне того что в норме, наоборот, насыщенная кислородом кровь течет cлева направо — из правой дуги аорты в левую через паниццево отверстие. Этим так называемым зубцовым клапаном крокодилы могут управлять по своей воле. Данный механизм, называемый контролируемым шунтированием, имеет важные физиологические функции. Во-первых, вместе со снижением частоты сердечных сокращений, данный механизм играет роль во временном понижении уровня обмена веществ, что необходимо для снижения потребления кислорода при погружении под воду. Во-вторых, при длительном погружении зубцовый клапан выступает в качестве предохранительного клапана, когда избыточное давление в легких вызывает застой крови в легочной артерии и повышенное давление в легочной вене, чтo распространяется на левый желудочек и правую аорту[34]. В-третьих, для ускорения пищеварения кровь в левой дуге аорты, ведущей к пищеварительной системе, сменяется с артериальной на венозную, насыщенную углекислотой, что приводит к выработке желудочного сока, по кислотности десятикратно превосходящего нормальный для млекопитающих[35].
Сердце гребнистого крокодила (Crocodylus porosus) массой 1,09 кг занимает около 0,279 % от массы тела[36]. Аналогичные числа для миссисипских аллигаторов массой 1 кг, 70 кг и 124 кг составляют 0,25 %, 0,15 % и 0,125 % соответственно[34]. Нормальная частота сердечных сокращений при температурe тела в тридцать градусов по Цельсию достигает двадцати пяти — тридцати пяти ударов в минуту. При погружении в воду в случае опасности крокодилы сначала увеличивают частоту сердечных сокращений, a потом замедляют её, при этом она может упасть до двух ударов в минуту.
Кровь крокодилов принципиально не отличается от крови других рептилий. Эритроциты имеют ядра, a содержание гемоглобина колеблется от 6,9 дo 9,0 грамм-процентов. Белые кровяные тельца представлены агранулоцитами, большими и малыми моноцитами и эозинофильными и нейтрофильными гранулоцитами. Крокодилий гемоглобин переносит гораздо больше кислорода, чем человеческий, а химический механизм регулирования выпуска кислорода зависит не от фосфатов, как у человека, а от бикарбонатов. По этой причине крокодилий гемоглобин может работать вне эритроцитов[37] Сильные антибиотики, содержащиеся в крови, помогают избежать заражения в загрязнённой воде[38].
Лимфатическая система
Лимфатическая система развита y крокодилов лучше, чем y большинства рептилий, и имеет сосудистое строение. Сети лимфатических сосудов расположены y мест прикрепления брыжейки, y основания хвоста, под мышками и на затылке.
Органы пищеварения
.jpg.webp)
Зубы у крокодилов конической формы и полые внутри — в этих полостях по мере снашивания развиваются новые острые зубы. Каждый зуб сменяется примерно раз в 1-2 года в зависимости от размеров крокодила. При этом зубы верхней и нижней челюстей совмещены таким образом, что против самых крупных зубов одной челюсти находятся самые мелкие зубы второй. Зубы текодонтного типа и сидят в альвеолах, как у млекопитающих. В отличие от большинства рептилий, у крокодилов наблюдается своеобразная дифференциация зубов по размерам, форме и даже выполняемым функциям. В начале каждой челюсти присутствуют по четыре крупных клыковидных зуба (у настоящих крокодилов особенно увеличен четвёртый зуб на нижней челюсти), служащих для эффективного захвата добычи. В середине челюсти зубы толще, равномернее увеличиваются по длине и образуют своеобразный ножницеподобный прикус, позволяющий разрезать захваченную жертву. У основания челюсти зубы расширяются и уплощаются, что служит для эффективного раскусывания панцирей черепах, раковин моллюсков или дробления костей. Некоторые виды, такие как широкомордый кайман, имеют преимущественно дробящий функциональный морфотип зубов, а другие, такие как гангский гавиал, исключительно прокалывающий. Любопытно, что некоторые сухопутные родственники крокодилов из числа нотозухий (Notosuchia) имели полностью дифференцированные зубы, подобно млекопитающим.
Язык крокодилов на всём протяжении прикреплён широкой нижней частью к дну ротовой полости и подвижность его очень мала. К задней части языка примыкает вентральная складка глоточной заслонки. Основу её составляет продолжение базигиоидного хряща, а наиболее близко расположенной мышечной тканью является поперечнополосатая мускулатура языка, для которого базигиоидный хрящ является точкой прикрепления. И, хотя сама по себе вентральная складка не является мышечной структурой, она является частью подъязычного аппарата и связана с основанием языка, что означает возможность её подвижности, обусловленной индивидуальной или комбинированной мышечной активностью.
Желудок делится на два отдела: мускулистый и пилорический. Первый снабжен наружными мышечными тяжами, крепящимися к сухожильным пластинам по обеим сторонам желудка. Внутренняя поверхность снабжена ороговевшей радулой (тёркой). Второй отдел желудка предназначен для переваривания уже растертой в предыдущем отделе пищи. Он имеет складчатую слизистую оболочку. Эпителий образован призматическими клетками, между которыми открываются соковые железы. В желудках крокодилов часто находят камни (гастролиты) и другие инородные предметы. По одной из гипотез, крокодилы заглатывают их, чтобы лучше перетирать пищу, по другой — для увеличения массы и ускорения погружения, однако точного объяснения этому явлению пока нет. Дело в том, что размеры гастролитов иногда неоправданно велики и не позволяют использовать их для эффективного перетирания добычи, кроме того, в зоопарках крокодилы часто лишены возможности заглатывать камни, что вовсе не ухудшает их пищеварения.
Кишечник подразделяется на тонкий и толстый, толстый кишечник заканчивается прямой кишкой. Последняя открывается в клоаку, куда также открываются мочеточники и протоки половых желез. Слепая кишка y крокодилов рудиментарна и находится на границе толстого и тонкого кишечников. Общая длина кишечника в полтора — два раза превышает общую длину тела (от кончика морды до кончика хвоста).
Двухдольная печень крокодилов расположена в краниальном направлении от диафрагмы. Желчный пузырь крупный и находится рядом c нижней стороной нижней доли правого легкого.
Поджелудочная железа y взрослых крокодилов сильно варьируется по размерам и форме. Топографически она прилегает к первой петле тонкого кишечника в виде продолговатого тела.
До недавних пор считалось, что все представители отряда крокодилов являются облигатными гиперплотоядными. Однако, современные исследования показали, что это суждение верно только для настоящих крокодилов и гавиаловых. В отличие от них, аллигаторовые могут переваривать растительные белки и жиры, а также демонстрируют в 5 раз большую активность ферментов, ответственных за переваривание углеводов. При разведении аллигаторов на фермах весьма эффективны оказались сложные рационы из высушенных гранул, содержащих до одной трети углеводного материала, включая кукурузу и рис. Однако, в отличие от аллигаторовых, настоящие крокодилы являются облигатными хищниками, и при коммерческом разведении на фермах им необходима строго мясная диета[18].
Органы выделения
Как и y всех рептилий, почки крокодилов относятся к тазовому типу (метанефрос). Почки парные, бобовидные, дольчатого строения. Мочевые канальцы начинаются мальпигиевыми тельцами и открываются в мочевые протоки, а те в свою очередь — в мочеточник. Мочевого пузыря нет, что, вероятно, связано с жизнью в воде. Излишки внутриполостной жидкости выводятся через абдоминальные поры.
Крокодилы выделяют жидкую прозрачную мочу c нейтральной или основной реакцией, главные продукты выделения в которой — мочевина, мочевая кислота и аммиак.
Органы размножения
Клоака в виде продольной щели. У самцов канал функционирует как семяпровод, при этом половые продукты попадают в него из семенников через придатки семенников. Семенники расположены вентральнее почек. В клоаке самцов располагается пенис, который в эрегированном состоянии выдвигается из неё наружу. Пенис крокодилов представляет собой непарный конусовидный валик на вентральной стенке клоаки, снабженный желобком для выведения спермы.
Половая система самок представлена яичникaми, расположенными так же, как и семенники самцов, и яйцеводами. Яйцевод подразделяется на воронку, среднюю часть, в которой формируются третичные яйцевые оболочки, и расширенный маточный отдел, открывающийся в клоаку.
Нервная система
Как и y всех высших позвоночных, головной мозг крокодилов образован пятью отделами — передний мозг, промежуточный мозг, средний мозг, задний мозг, мозжечок и продолговатый мозг. Однако он имеет особенности, которые отличают его от мозга представителей других групп рептилий и по строению приближают к мозгу птиц.
Обонятельные луковицы (производные переднего мозга) вынесены на конец длинной морды парой обонятельных отростков. Полый мозжечок разделен на три поперечных сегмента, что напоминает складчатый мозжечок птиц или млекопитающих. От головного мозга крокодилов отходит двенадцать пар полностью разделенных нервов, в то время как y других рептилий десятый и одиннадцатый нервы разделены не полностью. Крокодилам свойственна высокая быстрота реакции.
Продолговатый мозг переходит в спинной. Последний имеет спинномозговой канал гипоцентрического строения, два утолщения, соответствующих поясам конечностей, и продолжается в начало хвостового отдела позвоночника.
Органы чувств
Органы чувств крокодилов также характеризуются рядом особенностей. Так, на первом месте по значимости y крокодилов стоят органы зрения, слуха и осязания. Последнее для большинства представителей сборной группы рептилий нетипично.
Зрение

Верхнее веко развито сильнее нижнего; третье веко, образующее мигательную перепонку, может полностью закрывать глаз от внутреннего угла глаза к наружному. Все три века не содержат хрящевой ткани, но y всех видов аллигаторовых, a также y ложного гавиала и тупорылого крокодила в верхнем веке имеется небольшая зона окостенения. Нижнее веко, напротив, подвижное и содержит множество мышечных волокон.
Глаз крокодилов лишён склеротикального кольца, которое формируется y многих других рептилий вокруг роговой оболочки и образовано из костных пластинок. Зрачок вертикальный и обеспечивает широкий угол, равный приблизительно 270° — небольшие слепые зоны находятся за затылком и прямо перед мордой (в связи со слабым развитием бинокулярного зрения). B связи c частой ночной активностью у этих животных в сетчатке преобладают палочки, которые отвечают за ночное зрение и образуют скопление, отражающее свет. Этим обусловлено также красноватое свечение глаз крокодилов в темноте. Глаза крокодилов лишены слёзных желез, но имеют хорошо развитую Гардерову железу, которая открывается несколькими протоками под третьим веком. Эта железа выделяет жирный секрет, смазывающий роговицу, — именно её выделения и образуют «крокодиловы слёзы»[17].
Слух
Из всех рептилий крокодилы обладают наиболее развитым органом слуха, который по значимости стоит на втором месте после зрения.
Крокодилы имеют хорошо развитый щелевидный наружный слуховой проход, который закрывается клапаном, когда крокодил ныряет. Среднее ухо одной стороны связанно со средним ухом другой и c глоткой целой системой придаточных полостей, открывающихся в барабанные полости. При этом до сих пор неизвестно, какую роль для слуха играет анатомически связанная c ухом сложная система полостей в черепе крокодилов. Также неизвестно, насколько хорошо крокодилы слышат под водой. Во внутреннем ухе имеется улитка, подобная таковой y птиц.
Обоняние
Особенности органов обоняния у крокодилов связаны со строением черепа, a само обоняние по значимости стоит на третьем месте после зрения и слуха. Носовые проходы крокодилов вытянуты в соответствии со строением морды и выстланы тремя видами эпителия: ороговевший эпителий выстилает преддверье носовой полости; за ним следует область респираторного эпителия, который выстилает хоаны, вентральную часть носовой раковины и частично придаточные пазухи; обонятельный эпителий выстилает дорсальную часть носовой полости. Якобсонов орган y крокодилов отсутствует, хотя и закладывается на эмбриональных стадиях.
Осязание
Несмотря на сильное ороговение кожных покровов, y крокодилов имеются зоны кожной чувствительности (осязания), которые представлены осязательными рецепторами, располагающимися в щитках кожи — это так называемые чувствительные почки или нейромасты. Такие щитки часто группируются в осязательные пятна, превосходящие по чувствительности кончики пальцев человека. Они позволяют чувствовать вибрацию, исходящую от животных, находящихся в воде на большом расстоянии (аналог боковой линии рыб). Больше всего осязательных рецепторов располагается на морде и в пасти. B этих областях их может быть до двадцати на каждом щитке. Именно поэтому крокодилам c помощью пасти удается проделывать весьма тонкие манипуляции, такие как контроль захваченной кончиком челюстей жертвы или перенос детенышей. В остальных частях тела осязательные рецепторы могут располагаться по одному на щитке. У молодых крокодилов их больше, чем y взрослых. Недавно было установлено, что осязательные рецепторы крокодилов также способны определять химический состав воды[39].
Органы вкуса
Органы вкуса y крокодилов расположены на слизистой оболочке, покрывающей крыловидную кость и вентральную стенку глотки. При этом покровы собственно пасти ороговевшие и вкусовых луковиц не имеют.
Физиология
Осморегуляция
Настоящие крокодилы и гавиалы демонстрируют ряд приспособлений для жизни в солёной воде: у них имеются лингвальные солевые железы, сильно ороговевающий эпителий ротовой полости, препятствующий диффузии ионов и осмотическим потерям воды, и активную роль в осморегуляции играет клоака. В солёной воде у них включается сложная система осморегуляции, включающая почечный ответ, пост-почечную модификацию мочи в клоаке и выведение избытка хлорида натрия солевыми железами.

Но несмотря на то, что крупные крокодилы могут проводить много месяцев в солёной воде без видимого вреда для себя, особенности их осморегуляции все-таки не способны обеспечить постоянное пребывание в море. Солевые железы, удаляющие излишки хлорида натрия, у современных морских крокодилов менее специализированы, чем у морских черепах. В отличие от черепах, крокодилы никогда не пьют солёную воду даже при обезвоживании (что заложено у них на уровне инстинкта) и полностью полагаются на поступление воды из пищи. Для экономии воды аммониотеличные в пресной воде крокодилы в море становятся урикотеличными. Вместо большого количества прозрачной, сильно разбавленной жидкой мочи, в море крокодилы выделяют небольшое количество тягучей, пастообразной белой мочи или даже твёрдые пеллеты, состоящие в основном из солей мочевой кислоты. Для молодых животных проблема обезвоживания стоит острее: время выживания без периодического доступа к пище или пресной воде для новорождённого крокодила массой 100 г оценивается в 21 день, для молодого массой 1 кг — в 50 дней, для молодого массой 10 кг — примерно в 116 дней при летальном обезвоживании 33 % от массы тела[40][41][42].
Для аллигаторов жизнь в соленой воде не представляется возможной. Аллигаторы лишены солевых желёз и сильно ороговевающего эпителия ротовой полости, которые есть у крокодилов. Пресноводные популяции аллигаторов не способны отличать солёную воду от пресной и, находясь в гиперосмотических условиях, пьют солёную воду. Однако популяции из эстуариев избегают пить солёную воду — вероятно, хотя аллигаторы не избегают питья солёной воды на уровне инстинкта, они могут этому научиться. Клоака настоящих крокодилов — активный орган осморегуляции в солёной и пресной воде, способствующий сохранению воды и экскреции NaCl солевыми железами. Клоака аллигаторов не оказывает влияния на состав выводимой из организма мочи. У аллигаторов основным ответом на ионный и осмотический стресс является почечный — выведение лишних ионов с мочой. Но почки аллигаторов не способны производить концентрированную мочу, гиперосмотичную по отношению к крови, поэтому данный механизм не слишком эффективен. Для адекватной регидратации аллигаторам обязательно нужен доступ к пресной или слабосолоноватой воде[42].
По-видимому предки аллигаторов эволюционировали преимущественно в пресной воде, а предки настоящих крокодилов — в солоноватой воде (например, в эстуариях).
Образ жизни
Крокодилы распространены во всех тропических странах, обитая в разнообразных пресных водоёмах; представители семейств крокодилов терпимо относятся к солёной воде и некоторые из них встречаются в прибрежной части морей (острорылый крокодил, гребнистый крокодил). Предки крокодилов, как и многие другие крокодиломорфы, жили на суше, и только позже перешли к жизни в воде. Все современные крокодилы приспособлены к полуводному образу жизни — обитая в воде, яйца они, однако, откладывают на суше. Хотя некоторые вымершие крокодилы, такие как, к примеру, квинканы, вели полностью наземный образ жизни[43].
Некоторые крокодилы в периоды засухи впадают в спячку в вырытых ими неглубоких шахтах в иле на дне пересыхающих водоёмов.
Локомоция

Крокодилы — отличные пловцы. Во время плаванья массивный хвост крокодила извивается из стороны в сторону, проталкивая животное через толщу воды, в то время как конечности прижимаются к телу, чтобы уменьшить сопротивление. Когда крокодил решает остановиться или повернуть в другом направлении, конечности раздвигаются в стороны. Крокодилы обычно медленно дрейфуют у поверхности или под водой, совершая еле заметные движения хвостом, но при преследовании добычи способны развивать скорость свыше 25—30 км/ч, поддерживая этот темп довольно продолжительное время[44][45].

На суше крокодилы довольно медлительны и неуклюжи, но некоторые виды способны иногда предпринимать значительные переходы, удаляясь от водоёмов на несколько километров, и даже охотиться на суше. Строение конечностей крокодилов заметно отличаются от такового у других рептилий, позволяя им держать конечности почти что вертикально. Они имеют два основных способа наземной локомоции: простое ползание на животе и так называемую «высокую походку». «Высокая походка» более свойственна меньшим по размерам крокодилам. Она во многом напоминает способ передвижения млекопитающих, с той же последовательностью движения конечностей: левая передняя, правая задняя, правая передняя, левая задняя. Некоторые настоящие крокодилы и молодые гавиалы порой даже переходят на галоп, на короткие дистанции развивая скорость свыше 17 км/ч[9]. Ползание более свойственно крупным и тяжелым крокодилам. Оно не сильно отличается от метода передвижения саламандр и ящериц. Передвигаясь таким образом, крокодил тратит меньше сил, но развивает скорость не более 10-14 км/ч. При движении крокодилы могут использовать любой из этих двух способов локомоции в зависимости от обстоятельств. В отличие от большинства других наземных позвоночных, когда крокодилы разгоняются, они увеличивают скорость, при которой нижняя половина каждой конечности перемещается вперёд; таким образом длина шага увеличивается, а продолжительность шага уменьшается. Это позволят крокодилам практически мгновенно развить свою максимальную скорость бега и застать свою жертву врасплох[46].
На суше и в воде крокодилы могут высоко прыгать, отталкиваясь хвостом или задними конечностями.
Терморегуляция
Значительную часть суток крокодилы проводят в воде. На прибрежные отмели они выходят утром и ближе к вечеру, чтобы принять «солнечные ванны» (баскинг). Для охлаждения крокодил открывает рот, и вода испаряется изо рта.
У мелких особей суточные колебания температуры могут превышать 5 градусов, но благодаря особенностям поведения и строения тела, у крупных крокодилов суточные колебания температуры тела могут быть очень незначительными — около 1 градуса по Цельсию летом и около 1,5 градуса зимой[47]. Таким образом, для больших крокодилов характерна инерциальная гомойотермия. Однако, их не следует отождествлять с истинно теплокровными животными (в наше время — только птицами и млекопитающими), у которых постоянство температуры тела поддерживается за счёт собственного метаболизма (производства тепла), а не долгим остыванием.
Социальное поведение
 (аудио) | |
| Рёв аллигатора. | |
| Звуки самца миссисипского аллигатора в брачный период. | |
| Помощь по воспроизведению | |
Большинство видов крокодилов являются достаточно социальными животными, терпящими рядом с собой соразмерных сородичей. Однако, часто случаются драки за пространство или еду. В сезон размножения все крокодилы (особенно крупные самцы) становятся более территориальными и нетерпимыми к сородичам своего пола. Гребнистые крокодилы асоциальны и ведут территориальный образ жизни круглогодично, а не только в сезон размножения[48].
Как и многие другие завропсиды, крокодилы — очень агрессивные и смелые животные, предпочитающие драку отступлению. Некоторым исключением являются обычно пугливые аллигаторы и кайманы.
Акустическая коммуникация
Крокодилы — одни из немногих рептилий, обладающие голосом. Голос крокодила — нечто среднее между лаем и рёвом, особенно часто и громко слышен у наиболее социальных видов в период размножения, таким образом являясь способом социальной коммуникации. Детеныши могут использовать звуковые сигналы, напоминающие квакание, для привлечения внимания самки. Раздражённый крокодил может громко шипеть или даже рычать.
Питание

Все крокодилы являются хищниками и охотятся на других животных. Однако, было показано, что аллигаторовые являются скорее всеядными животными (мезохищниками), и способны по крайней мере в некоторых случаях включать в свой рацион растительность без вреда для здоровья[18][19]. Настоящим крокодилам необходима исключительно мясная диета[18], и уникальные случаи поедания фруктов некоторыми видами могут быть связаны с непищевым поведением[19].
Взрослые представители большинства видов крокодилов занимают верхнее положение в пищевой цепи и охотятся на большой спектр водных, околоводных и наземных животных, а также могут поедать падаль, хотя гнилого мяса избегают. У всех крокодилов набор предпочитаемых кормов меняется с возрастом и зависит от размеров животного: пищей молодым крокодилам служат в основном различные водные и околоводные беспозвоночные или мелкая рыба. Рацион взрослых особей различных видов варьируется в зависимости от их размеров, формы морды и строения зубов, напрямую связанных с относительными размерами и особенностями их излюбленной добычи. Для ловли мелких и подвижных животных в воде лучше всего подходят длинные и узкие челюсти с небольшими, но многочисленными и очень острыми зубами, близкими по форме и размерами (ложный гавиал, крокодил Джонстона). Для поедания небольших, но защищенных твердым панцирем животных (например, черепах и моллюсков), хорошо подходят очень широкие, приплюснутые челюсти с короткими и притупленными зубами (широкомордый кайман, американский аллигатор). Крокодилы, регулярно охотящиеся на очень крупную и сильную добычу, как правило имеют умеренно широкие челюсти, с крупными и прочными, но при этом относительно острыми зубами, отличающимися друг от друга по форме и размерам (большинство представителей рода Crocodylus). Встречаются и переходные формы челюстей (острорылый крокодил, оринокский крокодил)[49]. Бытующее мнение, что виды с широкими челюстями способны справляться с более крупной добычей за счет того, что их черепа способны выдерживать большие нагрузки, не соответствует действительности. Виды, охотящиеся на наиболее крупную добычу (нильский крокодил, гребнистый крокодил), как правило имеют умеренно-широкие челюсти. Считается, что относительно широкие челюсти некоторых видов, таких как болотный крокодил, являются скорее адаптацией для перемещения по заросшим континентальным водоемам, чем для охоты на относительно более крупную добычу[50].
Как правило крокодилы охотятся из засады, хотя стратегия охоты варьируются в зависимости от отдельных видов и преследуемой добычи. Наземную добычу крокодилы атакуют, выпрыгивая на неё из воды, хватая челюстями и затаскивая под воду. Большинство представителей семейства настоящих крокодилов, за исключением узкомордых форм, хорошо приспособлены для охоты на крупных животных и порой одолевают даже значительно превосходящих себя по размерам млекопитающих, пришедших на водопой. Охотясь на водную добычу, крокодилы либо активно преследуют её под водой, либо захватывают резким боковым движением челюстей. Некоторые крокодилы используют свои хвосты и тела, чтобы загонять рыбу на мелководье, либо другое довольно сложное поведение для ловли рыбы[2][49]. Они также могут выкапывать донных беспозвоночных[51]. Кубинские крокодилы и гладколобые кайманы Шнайдера[52] регулярно охотятся на суше, в то время как для других видов такое поведение относительно редко[53]. Крокодилам также свойственно отнимать добычу у других хищников (явление клептопаразитизма)[54].

Некоторым видам крокодилов свойственны акты сезонного кормления, имеющего место только в определённые периоды времени. Например, нильские крокодилы в некоторых районах Африки охотятся на мигрирующих зебр и антилоп гну, когда их стада пересекают реки. Такое изобилие добычи позволяет крокодилам выбирать наиболее слабых животных и топить их, либо вовсе подбирать затоптанных сородичами животных. Мадагаскарские нильские крокодилы находят пропитание в пещерах — взрослые особи предпочитают охотиться на лемуров и кистеухих свиней (в сухой сезон пещеры здесь — единственное место, где можно найти воду). Некоторые гребнистые крокодилы в определённый промежуток времени посещают побережья или отдельные острова, служащие местом гнездования различных видов морских черепах, наиболее уязвимых в этот промежуток времени[55]. В Австралии возле города Дарвин туристов развлекают прыжками крокодилов, приманивая их кусками мяса, подвешенными на верёвке рядом с туристическими судами. Многие крокодилы отличают туристические от других судов и сами плывут к ним. Крокодил может съесть и человека. Любой крокодил, превышающий 2 метра в длину и 30 кг веса, считается потенциально опасным, но наибольшее число неспровоцированных нападений на людей лежит на счету у гребнистых, болотных, острорылых и нильских крокодилов, а также чёрных кайманов[56].
Хотя, подобно большинству других не-млекопитающих хищников, жевать крокодилы не могут, крупную добычу они легко разрывают на подходящие по размерам куски при помощи своей мускулистой шеи и сильных челюстей, после чего проглатывают их целиком. Нередко они, вцепившись в живую или мертвую добычу, вращаются вокруг своей оси, чтобы оторвать кусок мяса. В расчленении особо толстокожих животных настоящим крокодилам помогает увеличенный четвёртый зуб нижней челюсти. Ранее считалось, что крокодилы могут оставлять свою жертву под водой, чтобы та подпортилась и её было проще расчленить на куски. Однако данная интерпретация в настоящее время признается ошибочной, и крокодилы скорее всего просто прячут про запас еду подобно тому, как это делают другие хищники[49]. У многих видов распространён каннибализм[57]. Крокодилы охотно поедают падаль и являются важными санитарами рек, хотя протухшего мяса всё таки избегают; они также прячут не съеденные остатки добычи про запас[49]. За один раз крокодилы могут съесть до 23 % от массы своего тела[35]. Будучи холоднокровными животными, крокодилы требуют примерно в пять раз меньше пищи, чем теплокровные хищники такого же размера[58]. Крокодилы хорошо адаптированы для длительного голодания. Без еды взрослые крокодилы в случае крайней необходимости могут обходиться примерно до одного года[59]. Даже только что вылупившиеся детеныши за счет понижения уровня обмена веществ способны прожить без пищи около 58 дней, потеряв при этом 23 % от своей массы[58].
Размножение

Самки разных видов откладывают яйца в песок на отмели либо зарывают в гнездо из грязи и гниющей листвы. Число яиц в кладке колеблется от 10 до 100 в зависимости от размеров и вида самки. На тенистых местах ямки закладываются неглубокие, на солнечных — до полуметра. Яйца засыпаются землёй (вперемешку с листьями и травой) или даже песком.[60] Самки по возможности обычно остаются возле кладки, охраняя яйца от врагов. Из всех яиц кладки детеныши вылупляются одновременно. Температура инкубации влияет на пол: если яйцо хранилось при температуре от 31 до 32 °C, то выводятся самцы, если выше или ниже — то самки[61]. Молодые крокодилы ещё внутри яиц ко времени вылупления издают квакающие звуки, и мать раскапывает кладку, помогает потомству выбраться и переносит их в своей пасти в водоём, хотя подобное поведение наблюдается не у всех видов. Ложные гавиалы, например, не заботятся о вылупившихся детенышах и даже не охраняют гнезда[62]. Наличие барорецепторов в ротовой полости крокодила позволяет самке быть предельно осторожной при переносе потомства. Кроме собственных детёнышей она иногда случайно подбирает и переносит в воду также детёнышей некоторых черепах, откладывающих яйца для безопасности вблизи от кладки крокодила (детёныши черепахи вылупляются одновременно с детёнышами крокодила и также способны самостоятельно добраться до воды).
В кариотипе у различных видов крокодилов количество хромосом в диплоидном наборе варьируется от 30 до 42 (32 у представителей рода Alligator, 42 у Caiman, Melanosuchus и Paleosuchus; 32 у Gavialis и Tomistoma, 30 у Mecistops, 38 у Osteolaemus и от 30 до 34 у Crocodylus). Кариотипы кайманов демонстрируют большое количество хромосомных плеч (60), в сравнении с аллигаторами, а самая большая хромосомная вариация внутри рода наблюдается у Crocodylus. Диплоидный набор хромосом 4 из 12 видов (C. porosus, C. siamensis, Crocodylus palustris и Crocodylus rhombifer) отличается от 32, несмотря на то, что имеет такое же фундаментальное число (58)[63].
Взросление и смертность
У взрослых крокодилов крупных видов нет никаких естественных врагов за исключением их сородичей и человека. Однако, многие животные, в том числе черепахи, птицы (например, цапли), различные млекопитающие и рептилии (особенно вараны), охотно поедают яйца и детенышей крокодилов. Очень маленький процент молодняка достигает подросткового возраста. Но по достижении массы в несколько килограмм смертность крокодилов в результате хищничества заметно снижается. Главными врагами молодых особей, а также крокодилов более мелких видов, являются более крупные крокодилы. Реже на них могут нападать другие крупные хищники, такие как питоны, акулы и большие кошки. На взрослых якарских или крокодиловых кайманов в Южной Америке регулярно охотятся ягуары, анаконды и гигантские выдры, но их самыми главными врагами кайманов в большинстве регионов являются черные кайманы, оринокские и острорылые крокодилы[57].
Половой зрелости крокодилы достигают в возрасте от 4—6 (мелкие виды) до примерно 10 или 15 (крупные виды) лет. Кроме того, они продолжают активно расти достаточно большой период времени и после наступления половой зрелости, благодаря растущим хрящевым участкам в костях. Однако, то, что крокодилы растут всю жизнь, является мифом. Живут крокодилы в среднем свыше 50, возможно до 70—100 лет[64]. Гордон Григг описывает продолжительность жизни крокодилов, как сопоставимую с человеческой[2].
Значение для людей
Крокодилы в разной степени опасны для человека. Некоторые никогда не нападают на человека (гавиал), другие нападают систематически (гребнистый крокодил, нильский крокодил), третьи (черный кайман, острорылый крокодил) нападают изредка. В России крокодилы входят в Перечень животных, запрещенных к содержанию[65].
Несмотря на всю потенциальную опасность, а может быть и благодаря ей, крокодилы исторически являются одними из самых почитаемых животных на нашей планете. Во многих древних или традиционных культурах крокодил считался священным животным. Так, например, в Древнем Египте возводились храмы, посвящённые богу-крокодилу Себеку, а в прихрамовых гробницах было найдено множество забальзамированных рептилий. Крокодилы, как правило, ассоциировались с силой и властью[66].
Мясо крокодилов съедобно и употребляется в пищу населением многих тропических стран. Известно, что мясо крокодилов приобретает вкус того, чем они в основном питаются. Кожа крокодилов, особенно аллигаторов, используется для изготовления различных галантерейных изделий (портфелей, чемоданов и т. п.).
Хищническое истребление крокодилов в 19-20 веках привело к резкому сокращению их численности (что отразилось на других животных; в частности, в некоторых районах Южной Америки после исчезновения кайманов выросло число пираний) и принятию охранных мер. В ряде стран (например, в Таиланде) существуют специальные фермы по разведению крокодилов.
Джером Горсей в конце XVI века отметил в дневнике: «Я выехал из Варшавы вечером, переехал через реку, где на берегу лежал ядовитый мёртвый крокодил». Под 1582 годом в Псковской летописи можно прочесть: «Того ж лета изыдоша коркодили лютии звери из реки, и путь затвориша, людей много поядоша…». Слово «поядоша» соответствует современному «покусать», а не «съесть». Вероятно, молодых рептилий везли для чьего-либо зверинца, и они случайно оказались на свободе[67]. Однако, возможно, крокодилы в обоих случаях были ни причем[68].
Крокодиловы слёзы
Существует древняя легенда о том, что крокодил, поедая добычу, плачет «крокодиловыми слезами». Наиболее раннее известное упоминание о крокодиловых слезах встречается в Библиотеке константинопольского патриарха Фотия (около 820—896). Фотий сообщает[69] о крокодилах, «которые, как говорят, головы людей оплакивают, которых они сожрали, и слезы льют на останки убиенных», из-за того, что «костистая голова не годится в пищу».
На самом деле крокодилы «плачут» вовсе не от жалости. У многих морских рептилий и птиц для удаления переизбытка солей в организме служат специальные железы, открывающиеся наружу у самых глаз. По этой причине длительное время считалось, что «крокодиловы слёзы» — это защитная реакция организма, направленная на выведение избытка солей. Однако, таким образом настоящие крокодилы выделяют значительно меньшее количество солей, чем другие живущие в соленой воде рептилии, и куда более важную роль в осморегуляции у них имеют лингвальные (располагающиеся на языке) солевые железы[17]. Вероятно, «крокодиловы слёзы» в основном выделяются в результате раздражения глаз крокодилов на жарком сухом воздухе.
Эволюция

Крокодилы — единственные выжившие представители подклады архозавров (Archosauria), за исключением отделяемых в традиционной классификации от других завропсидов птиц. На филогенетическом древе они находятся ближе к динозаврам и птицам, нежели к чешуйчатым или клювоголовым (в то время как положение черепах остается спорным). Современные крокодилы являются остатками обширной группы крокодиломорфов, произошедшей от текодонтов в позднем триасе около 225 млн лет назад; большинство из них вымерло к началу кайнозоя вместе с динозаврами. Считается, что первые представители современного отряда крокодилов появились примерно 83.5 млн лет назад — и уже тогда произошло отделение более примитивных в эволюционном плане аллигаторовых от линии, ведущей к гавиалам и настоящим крокодилам, предки которых разделилась несколько позже[70]. Крокодилы современного типа эволюционировали в направлении наибольшего приспособления к водному образу жизни и хищничеству, хотя среди всего многообразия крокодиломорфов были не только морские, полуводные или наземные хищники, но и даже всеядные или растительноядные наземные формы.
Современные крокодилы приближены к классическому «рептильному» уровню организации. Однако, ряд особенностей их анатомии позволяет сделать вывод о том, что современные полуводные крокодилы происходят из высокоорганизованных наземных животных, по многим признакам напоминающих птиц или динозавров:
- 4-камерное сердце. Млекопитающие и птицы имеют четырёхкамерные сердца. У холоднокровных пресмыкающихся трехкамерные сердца, которые менее эффективны для аэробного метаболизма, потому что они позволяют венозной и артериальной крови смешиваться и, следовательно, отправляют часть венозной крови в тело, а не в легкие. Сердца современных крокодилов четырёхкамерные, но меньше по размеру и работают при более низком давлении, чем у современных млекопитающих и птиц. Они также функционируют как трехкамерные, когда животное находится под водой[71].
- Имеется диафрагма.
- Прогрессивный печеночно-поршневой механизм («печеночный насос») для вентиляции легких. Он отличается от механизмов вентиляции легких у млекопитающих, птиц и наиболее активных современных холоднокровных лепидозавров (варанов), и похоже что является уникальным «изобретением» крокодиломорфов.
- Крокодилы могут двигаться, держа конечности почти что парасагиттально.
- Вторичное костное небо, которое позволяет животному есть и дышать одновременно.
Ещё в конце 80-х годов было предположено, что предки крокодилов были активными, теплокровными хищниками, и что их предки — архозавры также были теплокровными. В последнее время исследования в области эмбриологии показывают, что сначала у крокодильих эмбрионов развиваются полностью сформированные четырёхкамерные сердца, а затем развиваются модификации, которые позволяют их сердцам функционировать под водой как трехкамерным. Используя принцип, согласно которому онтогенез частично повторяет филогенез, Сеймур и соавт. (2004) пришли к выводу о том, что у наземных предков крокодилов было обычное четырёхкамерное сердце, что они были теплокровными, активными животными, и что позже при переходе к полуводному образу жизни их сердце видоизменилось, чтобы функционировать как трехкамерное, уровень метаболизма заметно снизился, а конечности приняли более естественный для холоднокровных животных вид[34][72]. Однако, признаки активных наземных животных скорее всего сохранялись и совершенствовались у групп крокодиломорфов, продолживших вести наземный образ жизни[73].
Об этом же свидетельствуют и палеонтологические данные: многие наземные крокодиломорфы, в том числе самые базальные формы, такие как сфенозухии, имеют длинные парасагиттальные конечности, передвижение при помощи которых было бы крайне невыгодно для холоднокровных животных в энергетическом плане. Гистологические срезы костей крокодиломорфов и более базальных псевдозухий указывают на то, что они росли быстрее современных крокодилов и с сопоставимой скоростью со многими теплокровными животными[74][75]. Скорее всего, теплокровность — это базальный признак для всех архозавров, поскольку исследования костных структур и их связи с темпами роста свидетельствовали о том, что даже ранние архозавры имели довольно высокий уровень метаболизма[76][77].
Филогенетические отношения
Отряд крокодилов в настоящее время располагается внутри клады Eusuchia, включает в себя 3 надсемейства, 3 семейства и 8 родов (из которых половина — монотипные)[78] и не менее 24 видов, с учётом недавно отделенного пустынного крокодила[79].
Кладограмма от Эриксона и соавт. (2012)[30]:
| Crocodilia |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Классификация

Надотряд Крокодиломорфы Crocodylomorpha
- Отряд Крокодилы Crocodilia
- Надсемейство Crocodyloidea
- Семейство Крокодиловые Crocodylidae
- Подсемейство Crocodylinae
- Род Настоящие крокодилы Crocodylus
- Острорылый крокодил Crocodylus acutus
- Африканский узкорылый крокодил Crocodylus cataphractus
- Оринокский крокодил Crocodylus intermedius
- Австралийский узкорылый крокодил Crocodylus johnstoni или Crocodylus johnsoni
- Филиппинский крокодил Crocodylus mindorensis
- Центральноамериканский крокодил Crocodylus moreletii
- Нильский крокодил Crocodylus niloticus
- Новогвинейский крокодил Crocodylus novaeguineae
- Crocodylus halli
- Болотный крокодил Crocodylus palustris
- Гребнистый крокодил Crocodylus porosus
- Кубинский крокодил Crocodylus rhombifer
- Сиамский крокодил Crocodylus siamensis
- Западноафриканский крокодил Crocodylus suchus[80]
- † Crocodylus anthropophagus
- † Crocodylus checchiai
- † Crocodylus falconensis
- † Crocodylus palaeindicus
- † Crocodylus thorbjarnarsoni
- Род Osteolaemus
- Тупорылый крокодил Osteolaemus tetraspis
- Род † Euthecodon
- † Euthecodon arambourgi
- † Euthecodon brumpti
- † Euthecodon nitriae
- Род † Rimasuchus
- † Rimasuchus lloydi или Crocodylus lloydi
- Род † Voay
- † Мадагаскарский крокодил Voay robustus или Crocodylus robustus
- Род Настоящие крокодилы Crocodylus
- Подсемейство Crocodylinae
- Семейство † Mekosuchidae
- Подсемейство † Mekosuchinae
- Род † Australosuchus
- Род † Baru
- † Baru darrowi
- † Baru huberi
- † Baru wickeni
- Род † Harpacochampsa
- † Harpacochampsa camfieldensis
- Род † Kambara
- † Kambara implexidens
- † Kambara molnari
- † Kambara murgonensis
- † Kambara taraina
- Род † Mekosuchus
- † Mekosuchus inexpectatus
- † Mekosuchus kolpokasi
- † Mekosuchus sanderi
- † Mekosuchus whitehunterensis
- Род † Pallimnarchus
- † Pallimnarchus gracilis
- † Pallimnarchus pollens
- Род † Квинкана Quinkana
- † Quinkana babarra
- † Quinkana fortirostrum
- † Quinkana timara
- † Quinkana meboldi
- Род † Trilophosuchus
- † Trilophosuchus rackhami
- Род † Volia
- † Volia athollandersoni
- † ?Volia vitiensis
- Подсемейство † Mekosuchinae
- Семейство Крокодиловые Crocodylidae
- Надсемейство Alligatoroidea
- Род † Deinosuchus
- † Deinosuchus rugosus или Polyptychodon
- † Deinosuchus riograndensis или Phobosuchus
- Семейство Аллигаторовые Alligatoridae
- Род † Notocaiman
- † Notocaiman stromeri
- Подсемейство Alligatorinae
- Род † Albertochampsa
- † Albertochampsa langstoni
- Род Аллигаторы Alligator
- † Alligator mcgrewi
- † Alligator mefferdi
- Миссисипский аллигатор Alligator mississippiensis
- † Alligator olseni
- † Alligator prenasalis
- Китайский аллигатор Alligator sinensis
- Род † Allognathosuchus
- † Allognathosuchus polyodon или Crocodylus polyodon
- † ?Allognathosuchus gracilis
- † Allognathosuchus heterodon или Alligator heterodon
- † ?Allognathosuchus isolatus или Manracosuchus isolatus
- † ?Allognathosuchus wartheni
- † ?Allognathosuchus woutersi
- Род † Arambourgia
- † Arambourgia gaudryi
- Род † Ceratosuchus
- † Ceratosuchus burdoshi
- Род † Chrysochampsa
- † Chrysochampsa mlynarskii
- Род † Eoalligator
- † Eoalligator chunyii
- † Eoalligator huiningensis
- Род † Hassiacosuchus
- † Hassiacosuchus haupti
- Род † Hispanochampsa
- † Hispanochampsa muelleri
- Род † Krabisuchus
- † Krabisuchus siamogallicus
- Род † Navajosuchus
- † Navajosuchus mooki или Allognathosuchus mooki
- Род † Procaimanoidea
- † Procaimanoidea utahensis
- † Procaimanoidea kayi или Hassiacosuchus kayi
- Род † Wannaganosuchus
- † Wannaganosuchus brachymanus
- Род † Albertochampsa
- Подсемейство Caimaninae
- Род † Centenariosuchus
- † Centenariosuchus gilmorei
- Род † Culebrasuchus
- † Culebrasuchus mesoamericanus
- Род † Eocaiman
- † Eocaiman cavernensis
- † Eocaiman palaeocenicus
- † Eocaiman itaboraiensis
- Род † Globidentosuchus
- † Globidentosuchus brachyrostris
- Род † Mourasuchus
- † Mourasuchus amazonensis
- † Mourasuchus arendsi
- † Mourasuchus atopus
- † Mourasuchus nativus
- Род † Necrosuchus
- † Necrosuchus ionensis
- Род † Orthogenysuchus
- † Orthogenysuchus olseni
- Род Гладколобые кайманы Paleosuchus
- Гладколобый кайман Paleosuchus palpebrosus
- Гладколобый кайман Шнайдера Paleosuchus trigonatus
- Род † Purussaurus
- † Purussaurus brasiliensis
- † Purussaurus neivensis
- † Purussaurus mirandai
- Род † Tsoabichi
- † Tsoabichi greenriverensis
- Род Кайманы Caiman
- Крокодиловый кайман Caiman crocodilus
- Широкомордый кайман Caiman latirostros
- † Caiman lutescens
- † Caiman venezuelensis
- Парагвайский кайман Caiman yacare
- Род Melanosuchus
- † Melanosuchus fisheri
- Чёрный кайман Melanosuchus niger
- Род † Centenariosuchus
- Род † Notocaiman
- Род † Deinosuchus
- Надсемейство Gavialoidea
- Род † Argochampsa
- † Argochampsa krebsi
- Род † Eosuchus
- † Eosuchus lerichei
- † Eosuchus minor или Gavialis minor
- Род † Eothoracosaurus
- † Eothoracosaurus mississippiensis
- Род † Thoracosaurus
- † Thoracosaurus neocesariensis или Gavialis neocesariensis
- † Thoracosaurus macrorhynchus
- † Thoracosaurus scanicus
- Семейство Гавиаловые — Gavialidae
- Подсемейство Gavialinae
- Род † Eogavialis
- † Eogavialis africanus
- † Eogavialis andrewsi
- † Eogavialis gavialoides или Tomistoma gavialoides
- Род Гавиалы Gavialis
- † Gavialis curvirostris
- † Gavialis breviceps
- † Gavialis bengawanicus
- Гангский гавиал Gavialis gangeticus
- † Gavialis lewisi
- Род † Eogavialis
- Подсемейство Gryposuchinae
- Род † Aktiogavialis
- † Aktiogavialis puertoricensis
- Род † Gryposuchus
- † Gryposuchus colombianus
- † Gryposuchus croizati
- † Gryposuchus jessei
- † Gryposuchus neogaeus или Ramphostoma neogaeus
- Род † Hesperogavialis
- † Hesperogavialis cruxenti
- Род † Ikanogavialis
- † Ikanogavialis gameroi
- † ?Ikanogavialis papuensis или Gavialis papuensis
- Род † Piscogavialis
- † Piscogavialis jugaliperforatus
- Род † Siquisiquesuchus
- † Siquisiquesuchus venezuelensis
- Род † Aktiogavialis
- Подсемейство Tomistominae
- Род Tomistoma
- † Tomistoma cairense
- † ?Tomistoma lusitanicum
- Гавиаловый крокодил Tomistoma schlegelii
- Род † Dollosuchoides
- † Dollosuchoides densmorei
- Род † Dollosuchus
- † Dollosuchus dixoni
- Род † Ferganosuchus
- † Ferganosuchus planus
- Род † Gavialosuchus
- † ?Gavialosuchus americanus или Tomistoma americanus
- † ?Gavialosuchus carolinensis
- † Gavialosuchus eggenburgensis
- Род † Kentisuchus
- † Kentisuchus champsoides или Crocodylus champsoides
- † Kentisuchus spenceri или Crocodylus spenceri
- † Kentisuchus toliapicus или Crocodylus toliapicus
- Род † Maroccosuchus
- † Maroccosuchus zennaroi
- Род † Megadontosuchus
- † Megadontosuchus arduini или Crocodilius arduini
- Род † Paratomistoma
- † Paratomistoma courti
- Род † Penghusuchus
- † Penghusuchus pani
- Род † Rhamphosuchus
- Род † Toyotamaphimeia
- † Toyotamaphimeia machikanensis или Tomistoma machikanensis
- Род † Thecachampsa
- † Thecachampsa antiqua
- † Thecachampsa americana или Tomistoma americanus
- † Thecachampsa carolinense или Gavialosuchus carolinensis
- Род Tomistoma
- Подсемейство Gavialinae
- Род † Argochampsa
- Надсемейство Crocodyloidea
Такие группы крокодиломорфов, как Mesosuchia, Protosuchia, Sphenosuchia, Thalattosuchia и другие, вопреки распространенной ошибке, в настоящее время не включаются в отряд крокодилов и не являются крокодилами в правильном значении этого термина.
В искусстве
- Крокодилы в произведениях Корнея Чуковского
- Крокодил Гена
- Детское стихотворение «Крокодилы» Ирины Токмаковой
- Крокодил (журнал)
- Крокодилы присутствуют в таких фильмах ужасов, как «Аллигатор» (1980), «Река большого крокодила», «Век невежества», «Кровавый сёрфинг», «Лэйк Плэсид: Озеро страха» (1999), «Крокодил» (2000), «Крокодил» (2007).
В геральдике


.svg.png.webp) Герб французского города Ним
Герб французского города Ним
См. также
Примечания
- Raymond T. Hoser (2012), «A review of the taxonomy of the living crocodiles including the description of three new tribes, a new genus, and two new species». Australasian Journal of Herpetology 14, 9-16.
- Gordon Grigg, David Kirshner, Rick Shine. Biology and Evolution of Crocodylians. — 1 edition. — Comstock Publishing Associates, 2015-05-26. — 672 с. — ISBN 9780801454103.
- Liddell, Henry George; Scott, Robert (1901). «An Intermediate Greek-English Lexicon». Tufts University. Retrieved 22 October 2013.
- Schmidt, K.P. 1953. A Checklist of North American Amphibians and Reptiles. Sixth edition. Amer. Soc. Ichthy. Herp. Chicago, University of Chicago Press.
- Dundee, H.A. 1989. Higher Category Name Usage forAmphibians and Reptiles. Syst. Zool. Vol. 37:410-419.
- Wermuth, H. 1953. Systematik der Rezenten Krokodile. Mitt. Mus. Berlin. Vol. 29(2):275-514.
- Brochu, C.A. 2003. Phylogenetic Approaches Toward Crocodylian History. Annu. Rev. Earth Planet. Sci. Vol. 31:357-397.
- А. Богданов. О крокодилах в России. // Очерки из истории заимствований и экзотизмов. — М.: НЛО, 2006, с. 146—181 (Дата обращения: 25 мая 2011)
- John R. Hutchinson, Dean Felkler, Kati Houston, Yu-Mei Chang, John Brueggen. Divergent evolution of terrestrial locomotor abilities in extant Crocodylia (англ.) // Scientific Reports. — 2019-12-17. — Vol. 9, iss. 1. — P. 1–11. — ISSN 2045-2322. — doi:10.1038/s41598-019-55768-6.
- Milàn, J. & Hedegaard, R. Interspecific variation in tracks and trackways from extant crocodylians. Crocodyle Tracks and Traces. New Mexico Mus. Nat. Hist. Sci. Bull. 51, 15-29 (2010).
- Chamero, B., Buscalioni, Á. D. & Marugán-Lobón, J. Pectoral girdle and forelimb variation in extant Crocodylia: the coracoid-humerus pair as an evolutionary module. Biol. J. Linn. Soc. 108, 600—618 (2013).
- Owerkowicz, T. & Baudinette, R. V. Exercise training enhances aerobic capacity in juvenile estuarine crocodiles (Crocodylus porosus). Comp. Biochem. Physiol. A: Mol. Integr. Physiol. 150, 211—216 (2008).
- Kubo, T. Variation in modern crocodylian limb kinematics and its effect on trackways. Crocodyle Tracks and Traces. New Mexico Mus. Nat. Hist. Sci. Bull. 51, 51-53 (2010).
- Masaya Iijima, Tai Kubo, Yoshitsugu Kobayashi. Comparative limb proportions reveal differential locomotor morphofunctions of alligatoroids and crocodyloids (англ.) // Open Science. — 2018-03-01. — Vol. 5, iss. 3. — P. 171774. — ISSN 2054-5703. — doi:10.1098/rsos.171774.
- Iijima, M. & Kubo, T. Comparative morphology of presacral vertebrae in extant crocodylians: taxonomic, functional and ecological implications. Zool. J. Linn. Soc. 186, 1006—1025 (2019).
- Allen, V. et al. Comparative architectural properties of limb muscles in Crocodylidae and Alligatoridae and their relevance to divergent use of asymmetrical gaits in extant Crocodylia. J. Anat. 225, 569—582 (2014)
- Laurence E. Taplin, Gordon C. Grigg. Salt Glands in the Tongue of the Estuarine Crocodile Crocodylus porosus // Science. — 1981. — Т. 212, вып. 4498. — С. 1045–1047. — ISSN 0036-8075.
- Christopher R. Tracy, Todd J. McWhorter, C. M. Gienger, J. Matthias Starck, Peter Medley. Alligators and Crocodiles Have High Paracellular Absorption of Nutrients, But Differ in Digestive Morphology and Physiology // Integrative and Comparative Biology. — 2015-12. — Т. 55, вып. 6. — С. 986–1004. — ISSN 1557-7023. — doi:10.1093/icb/icv060.
- S. G. Platt, R. M. Elsey, H. Liu, T. R. Rainwater, J. C. Nifong. Frugivory and seed dispersal by crocodilians: an overlooked form of saurochory? (англ.) // Journal of Zoology. — 2013. — Vol. 291, iss. 2. — P. 87–99. — ISSN 1469-7998. — doi:10.1111/jzo.12052.
- Crocodilian Species - Johnston's Crocodile (Crocodylus johnstoni). crocodilian.com. Дата обращения: 1 мая 2020.
- Crocodilian Species - Dwarf Crocodile (Osteolaemus tetraspis). crocodilian.com. Дата обращения: 1 мая 2020.
- Crocodilian Species - Slender-snouted Crocodile (Mecistops cataphractus, Crocodylus cataphractus). crocodilian.com. Дата обращения: 1 мая 2020.
- Domenic C. D’Amore, Megan Harmon, Stephanie K. Drumheller, Jason J. Testin. Quantitative heterodonty in Crocodylia: assessing size and shape across modern and extinct taxa // PeerJ. — 2019-02-28. — Т. 7. — ISSN 2167-8359. — doi:10.7717/peerj.6485.
- Crocodilian Species - Chinese Alligator (Alligator sinensis). crocodilian.com. Дата обращения: 1 мая 2020.
- Crocodilian Species - Black Caiman (Melanosucus niger). crocodilian.com. Дата обращения: 1 мая 2020.
- Crocodilian Species - Rio Apaporis Caiman (Caiman crocodilus apaporiensis). crocodilian.com. Дата обращения: 1 мая 2020.
- Julia L. Molnar, Stephanie E. Pierce, Bhart-Anjan S. Bhullar, Alan H. Turner, John R. Hutchinson. Morphological and functional changes in the vertebral column with increasing aquatic adaptation in crocodylomorphs (англ.) // Open Science. — 2015-11-01. — Vol. 2, iss. 11. — P. 150439. — ISSN 2054-5703. — doi:10.1098/rsos.150439.
- A. F. Bennett, R. S. Seymour, D. F. Bradford, G. J. W. Webb. Mass-Dependence of Anaerobic Metabolism and Acid-Base Disturbance During Activity in the Salt-Water Crocodile, Crocodylus Porosus (англ.) // Journal of Experimental Biology. — 1985-09-01. — Vol. 118, iss. 1. — P. 161–171. — ISSN 1477-9145 0022-0949, 1477-9145.
- (2003) «The ontogeny of bite-force performance in American alligator (Alligator mississippiensis)» (PDF). Journal of Zoology 260(3): 317—327. DOI:10.1017/S0952836903003819.
- Gregory M. Erickson, Paul M. Gignac, Scott J. Steppan, A. Kristopher Lappin, Kent A. Vliet. Insights into the Ecology and Evolutionary Success of Crocodilians Revealed through Bite-Force and Tooth-Pressure Experimentation // PLOS ONE. — 2012-03-14. — Т. 7, вып. 3. — С. e31781. — ISSN 1932-6203. — doi:10.1371/journal.pone.0031781.
- Blanco, R. E.; Jones, W. W.; Villamil, J. N. (2014-04-16). «The 'death roll' of giant fossil crocodyliforms (Crocodylomorpha: Neosuchia): Allometric and skull strength analysis».
- Cogger, H.G. (1996). Reptiles and Amphibians of Australia. Chatswood, NSW: Reed Books.
- Аллигаторы перемещают лёгкие внутри тела. membrana (14 марта 2008). Дата обращения: 25 мая 2011.
- Seymour, R. S., Bennett-Stamper, C. L., Johnston, S. D., Carrier, D. R. and Grigg, G. C. (2004). «Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution». Physiological and Biochemical Zoology. 77 (6): 1051—1067. PMID 15674775. doi: 10.1086/422766.
- Крокодил переключает сердце для переваривания жертвы целиком. membrana (6 февраля 2008). Дата обращения: 25 мая 2011.
- null Franklin, null Axelsson. THE INTRINSIC PROPERTIES OF AN IN SITU PERFUSED CROCODILE HEART // The Journal of Experimental Biology. — 1994-1. — Т. 186, вып. 1. — С. 269–288. — ISSN 1477-9145.
- Кровь крокодилов может спасти людей. membrana (3 сентября 2006). Дата обращения: 11 апреля 2011.
- Кровь крокодилов спасёт людей от инфекций. membrana (16 августа 2005). Дата обращения: 25 мая 2011.
- Administrator. У крокодилов обнаружили суперрецепторы (недоступная ссылка). www.worldofanimals.ru. Дата обращения: 11 сентября 2017. Архивировано 30 мая 2017 года.
- L. E. Taplin, G. C. Grigg, L. Beard. Osmoregulation of the Australian freshwater crocodile, Crocodylus johnstoni, in fresh and saline waters (англ.) // Journal of Comparative Physiology B. — 1993-02-01. — Vol. 163, iss. 1. — P. 70–77. — ISSN 1432-136X 0174-1578, 1432-136X. — doi:10.1007/BF00309668.
- L. J. Kuchel, C. E. Franklin. Morphology of the cloaca in the estuarine crocodile, Crocodylus porosus, and its plastic response to salinity // Journal of Morphology. — August 2000. — Т. 245, вып. 2. — С. 168–176. — ISSN 0362-2525. — doi:10.1002/1097-4687(200008)245:23.0.CO;2-1.
- Osmoregulation in crocodilians (англ.). ufdc.ufl.edu. Дата обращения: 14 сентября 2017.
- Целлариус А. Ю. Динозавры. — «Астрель», 2001. — Т. 2. — С. 304—305. — 397,[3] с. — (Я познаю мир). — ISBN 5-271-01948-9.
- Wayback Machine (21 октября 2013). Дата обращения: 14 сентября 2017.
- Lang, J. W. (2002). «Crocodilians». In Halliday, T.; Adler, K. The Firefly Encyclopedia of Reptiles and Amphibians. Firefly Books. pp. 212—221. ISBN 1-55297-613-0.
- Reilly, S. M.; Elias, J. A. (1998). «Locomotion in Alligator mississippiensis: kinematic effects of speed and posture and their relevance to the sprawling-to-erect paradigm» (PDF). The Journal of Experimental Biology. 201(18): 2559—2574. PMID 9716509.
- Thermal relations of large crocodiles, Crocodylus porosus, free-ranging in a naturalistic situation (англ.) (22 сентября 1998). doi:10.1098/rspb.1998.0504. Дата обращения: 23 января 2013.
- Jeffrey Lang. Crocodilian Behaviour: Implications for Management, in G.W.J. Webb, S.C. Manolis, and P.J. Whitehead (eds.), Wildlife Management: Crocodiles and Alligators, pp. 273-294. Surrey Beatty and Sons, Sydney, Australia. (англ.).
- C. A. W. Guggisberg. Crocodiles: Their Natural History, Folklore and Conservation. — 1st ed edition. — Newton Abbot: David & Charles, 1972-03-16. — 200 с. — ISBN 9780715352724.
- William E. Magnusson. Crocodilia Diet (англ.) // Encyclopedia of Animal Cognition and Behavior / Jennifer Vonk, Todd Shackelford. — Cham: Springer International Publishing, 2017. — P. 1–5. — ISBN 978-3-319-47829-6. — doi:10.1007/978-3-319-47829-6_973-1.
- Kelly, Lynne (2007). Crocodile: Evolution’s greatest survivor. Orion. ISBN 978-1-74114-498-7.
- William E. Magnusson, Eduardo Vieira da Silva, Albertina P. Lima. Diets of Amazonian Crocodilians // Journal of Herpetology. — 1987. — Т. 21, вып. 2. — С. 85–95. — ISSN 0022-1511. — doi:10.2307/1564468.
- terrestrial hunting by crocodilians. dinets.info. Дата обращения: 1 мая 2020.
- Steven G. Platt, Thomas R. Rainwater, Scott Snider, Anthony Garel, Todd A. Anderson. [310:COLMBC2.0.CO;2/CONSUMPTION-OF-LARGE-MAMMALS-BY-CROCODYLUS-MORELETII--FIELD-OBSERVATIONS/10.1894/0038-4909(2007)52[310:COLMBC]2.0.CO;2.full CONSUMPTION OF LARGE MAMMALS BY CROCODYLUS MORELETII: FIELD OBSERVATIONS OF NECROPHAGY AND INTERSPECIFIC KLEPTOPARASITISM] // The Southwestern Naturalist. — 2007/06. — Т. 52, вып. 2. — С. 310–317. — ISSN 1943-6262 0038-4909, 1943-6262. — doi:10.1894/0038-4909(2007)52[310:COLMBC]2.0.CO;2.
- Knox, David. Airdate: Reptilian Battleground. TVTonight. Дата обращения: 29 декабря 2014.
- Welcome | CrocBITE (англ.). www.crocodile-attack.info. Дата обращения: 9 июля 2017.
- Ruchira Somaweera, Matthew Brien, Richard Shine. The Role of Predation in Shaping Crocodilian Natural History // Herpetological Monographs. — 2013-12-01. — Т. 27, вып. 1. — С. 23–51. — ISSN 0733-1347. — doi:10.1655/HERPMONOGRAPHS-D-11-00001.
- Garnett, S. T. (1986). «Metabolism and survival of fasting estuarine crocodiles». Journal of Zoology. 208(4): 493—502. doi: 10.1111/j.1469-7998.1986.tb01518.x.
- Крокодилам 120 млн. лет. НОВОСТИ СЕГОДНЯ (5 ноября 2010). Дата обращения: 25 мая 2011. Архивировано 25 августа 2011 года.
- Акимушкин, И. И. Пресмыкающиеся // Птицы. Рыбы, земноводные и пресмыкающиеся. — 3-е изд. — М.: «Мысль», 1995. — С. 415. — 462[1] с. — (Мир животных). — 25 000 экз. — ISBN 5-244-00803-X.
- Самые опасные животные: Сильнейшие хищники. Часть 2. Дата обращения: 26 ноября 2011.
- Tomistoma schlegelii (MÜLLER, 1838), crocodilian.com.
- Kornsorn Srikulnath, Watcharaporn Thapana, Narongrit Muangmai. Role of Chromosome Changes in Crocodylus Evolution and Diversity // Genomics & Informatics. — 2015-12. — Т. 13, вып. 4. — С. 102–111. — ISSN 1598-866X. — doi:10.5808/GI.2015.13.4.102.
- Животный мир Африки = Cuccioliin Afrika / Перевод с англ. А. М. Головы, под ред. И. Б. Шустовой. — М.: «Росмэн», 1997. — С. 12. — (Обитатели Земли). — ISBN 5-257-00174-8.
- Постановление Правительства Российской Федерации от 22 июня 2019 года № 795 "Об утверждении перечня животных, запрещенных к содержанию
- caroline seawright. Sobek, Ancient Egyptian God of Crocodiles (англ.) (недоступная ссылка). www.thekeep.org. Дата обращения: 19 июля 2017. Архивировано 25 июля 2017 года.
- Б. Сапунов. Легенда о том, как в реке Великой жили крокодилы (недоступная ссылка). Россия начинается здесь! Псковская земля. Дата обращения: 25 мая 2011. Архивировано 25 августа 2011 года.
- Как в Пскове охотились на «лютого зверя коркодела». Из книги Александры Барковой «Славянские мифы. От Велеса и Мокоши до птицы Сирин и Ивана Купалы». М.: Манн, Иванов и Фербер, 2022.
- Photius. Bibliotheque. Tome VIII… / Texte etabli et traduit par R. Henry. Paris (Collection Byzantine /.. / de 1’association Guillaume Bude), 1977. P. 93 (271. 503.а. 35).
- Brochu, C.A. (2003). «Phylogenetic approaches toward crocodylian history». Annual Review of Earth and Planetary Sciences. 31: 357—397. Bibcode:2003AREPS..31..357B. doi: 10.1146/annurev.earth.31.100901.141308.
- News in Science - Secret of the crocodile heart - 25/08/2000. www.abc.net.au. Дата обращения: 19 июля 2017.
- Summers, A.P. (2005). «Evolution: Warm-hearted crocs». Nature. 434 (7035): 833—834. Bibcode:2005Natur.434..833S. PMID 15829945. doi: 10.1038/434833a.
- Cenozoic dinosaurs in South America – Revisited (PDF Download Available) (англ.). ResearchGate. Дата обращения: 19 июля 2017.
- Nicole Klein, Christian Foth, Rainer R. Schoch. Preliminary observations on the bone histology of the Middle Triassic pseudosuchian archosaur Batrachotomus kupferzellensis reveal fast growth with laminar fibrolamellar bone tissue // Journal of Vertebrate Paleontology. — 2017-07-04. — Т. 37, вып. 4. — С. e1333121. — ISSN 0272-4634. — doi:10.1080/02724634.2017.1333121.
- Susan Drymala, Kate Dzikiewicz, Lindsay Zanno, Vince Schneider. Ontogeny and growth in basal crocodylomorphs and their close relatives. — 2014-11-06.
- Armand de Ricqlès, Kevin Padian, Fabien Knoll, John R. Horner. On the origin of high growth rates in archosaurs and their ancient relatives: Complementary histological studies on Triassic archosauriforms and the problem of a “phylogenetic signal” in bone histology // Annales de Paléontologie. — 2008-04-01. — Т. 94, вып. 2. — С. 57–76. — doi:10.1016/j.annpal.2008.03.002.
- Jorge Cubo, Jennifer Botha-Brink, Guillaume Guénard, Lucas J. Legendre. Palaeohistological Evidence for Ancestral High Metabolic Rate in Archosaurs (англ.) // Systematic Biology. — 2016-11-01. — Vol. 65, iss. 6. — P. 989–996. — ISSN 1063-5157. — doi:10.1093/sysbio/syw033.
- Русские названия по книге Ананьева Н. Б., Боркин Л. Я., Даревский И. С., Орлов Н. Л. Пятиязычный словарь названий животных. Амфибии и рептилии. Латинский, русский, английский, немецкий, французский. / под общей редакцией акад. В. Е. Соколова. — М.: Рус. яз., 1988. — С. 138—139. — 10 500 экз. — ISBN 5-200-00232-X.
- Crocodylus suchus, The Reptile Database. Дата обращения 19 июля 2017.
- The Reptile Database. Дата обращения: 12 мая 2016.
Литература
- А. Богданов. О крокодилах в России. // Очерки из истории заимствований и экзотизмов. — М.: НЛО, 2006, с. 146—181 (Дата обращения: 25 мая 2011)
- Островский А. Удивительная живучесть крокодильего племени // «Вокруг света», апрель 2011, № 4 (2847), рубрика «Спираль времени».
| Таксономия | |
|---|---|
| В библиографических каталогах |
