Стрижиные
Стрижи́ные[1][2], или настоящие стрижи[3][4], или стрижи[5] (лат. Apodidae), — семейство птиц из отряда стрижеобразных. Для них характерно плотное обтекаемое тело, длинные саблевидные крылья, уплощённая голова с коротким плоским клювом и короткие слабые ноги. Окрашены с преобладанием чёрных или серых тонов, брюшная сторона обычно светлее. Половой диморфизм отсутствует. Стрижи распространены повсеместно, кроме северных и южных полярных районов, большей части Австралии, некоторых океанических островов и центральных частей Сахары и Тибета.
| Стрижиные | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Чёрный стриж | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Подкласс: Инфракласс: Клада: Отряд: Подотряд: Семейство: Стрижиные |
||||||||||||
| Международное научное название | ||||||||||||
| Apodidae Hartert, 1897 | ||||||||||||
| ||||||||||||
Стрижи больше других птиц приспособлены к полёту. На лету они питаются, пьют воду, купаются, возможно также спят и спариваются. Основу рациона составляют насекомые, которых птицы добывают исключительно на лету, осуществляя «воздушное траление». Рацион стрижей не имеет точного соответствия по составу насекомых в конкретной местности, и он может существенно различаться у разных видов в зависимости от высоты, на которой птицы охотятся, размеров птиц, времени года и способности широко открывать рот. Также возможны существенные географические различия по составу рациона внутри одного вида. Стрижи сооружают гнёзда в дуплах деревьев, норах, глубоко в пещерах, на скалах или на искусственных сооружениях (под крышами домов, в щелях зданий и т. п.). Чтобы добраться до гнезда, некоторые виды стрижей используют эхолокацию, другие пролетают сквозь толщу воды в водопадах. Многие виды скрепляют гнездо с помощью слюны, а саланганы-водорослееды строят гнездо исключительно из неё, что является деликатесом восточной кухни. Стрижи моногамны, откладывают от одного до семи яиц, которые насиживают оба родителя. Птенцы вылупляются голыми и слепыми и проводят в гнезде до 1,5 месяцев. В поисках пищи родители могут покидать гнездо на несколько дней, при этом птенцы впадают в оцепенение.
В десятое издание «Системы природы» (1758) вошло четыре вида стрижей; Карл Линней отнёс их к ласточкам, немного напоминающим стрижей в полёте. В настоящее время общепризнанным является выделение двух подсемейств стрижей — Cypseloidinae и Apodinae, включающих 113 видов в составе 19 родов. Многие учёные разделяют подсемейство Apodinae на три трибы: cаланганы (Collocaliini), колючехвосты (Chaeturini) и типичные стрижи (Apodini). Самой примитивной группой стрижей является ограниченная Новым Светом Cypseloidinae, за ней следуют Collocaliini и Chaeturini, а наиболее развитой в эволюционном плане является группа Apodini.
Этимология
В «Системе природы» Карла Линнея чёрный стриж (Apus apus) был описан под названием Hirunso apus[6][7] (греч. a- — без, греч. pous — нога)[6][8][9]. В 1777 году итальяно-австрийский естествоиспытатель Джованни Антонио Скополи использовал название Apos[6][10], отделив стрижей от ласточек[10]; в той же работе встречается и Apus, под таким именем были описаны ракообразные (Crustacea). В 1810 году немецкие орнитологи Бернхард Мейер и Иоганн Вольф назвали стрижей Micropus (греч. mikros- — маленький, греч. pous — нога[11]), что было отвергнуто английским орнитологом Ричардом Боудлером Шарпом в 1900 году, так как такое название использовалось Линнеем для ботанического описания[6].
В 1811 году увидел свет «Систематический очерк млекопитающих и птиц» германского зоолога Иоганна Карла Вильгельма Иллигера. В этой работе выделенные в отдельный род стрижи получили латинское название Cypselus[6][12] (греч. kupselos — ласточка[12][13]). Такое название встречается в трудах Аристотеля[12]. В конце XIX века в ряде работ для обозначения семейства использовалось название Macropterigidae[14].
Русское название птицы получили за пронзительный голос[8].
Описание
Внешний вид
_(6040756987).jpg.webp)
Стрижи — мелкие птицы с массой от 10 до 170—200 г, при этом длина тела и размах крыльев у лёгких саланган составляет 9 см и 18 см, а у фиолетового колючехвоста[4] (Hirundapus celebensis) — 26 см и 55 см, соответственно. Птицы довольно схожи внешне: у них плотное обтекаемое тело, длинные саблевидные крылья, уплощённая голова с коротким плоским клювом. Кожа толстая. Половой диморфизм за редким исключением отсутствует, самцы и самки имеют схожие размеры и обычно не отличаются друг от друга окраской[4].
Стрижи характеризуются дальнозоркостью и способны фокусировать своё зрение как моноптически (одним глазом), так и биоптически (двумя глазами)[15]. На сетчатке глаза у них имеются как центральные, так и боковые ямки, в которых повышенная плотность фоторецепторов обеспечивает повышенную зрительную разрешающую способность. Центральная ямка (fovea centralis) представляет собой небольшое углубление, характеризующееся наибольшей концентрацией и поверхностным расположением фоторецепторов. Боковые ямки располагаются в боковом направлении от центральной. Боковая ямка (fovea lateralis), как полагают, важна для бинокулярного зрения птиц, а центральная ямка — для монокулярного[16]. У одного вида, пятнистолобого американского стрижа[3] (Cypseloides cherriei), глаза особенно крупные и составляют 5,5 % от общей массы тела птицы[15]. Радужина тёмная[4]. Непосредственно перед глазами расположены чёрные грубые щетинки, наклон которых может изменяться при помощи мышц. Они служат для защиты глаз и уменьшения солнечных бликов, чему также способствует глубоко утопленное положение глаз[15].
Клюв короткий[15], тёмный[4]. Разрез рта заходит за глаза. Несмотря на то, что клюв у стрижей очень маленький, рот может широко открываться[15][17], что связано с тем, что челюстной сустав находится далеко за глазом. Вибриссы отсутствуют[18]. Под маленьким треугольным языком[18][17] находится подъязычный мешок — в этой полости добыча для выкармливания птенцов обволакивается слюной[18].
У стрижей маленькие, покрытые мозолями ноги с острыми и изогнутыми когтями. Особенности строения ног и жёсткие перья хвоста позволяют стрижам закрепляться на вертикальных поверхностях[15], при этом они не могут сидеть на ветках, так как неспособны обхватить их[8][15]. Не имея возможности лазить и ходить на ровной поверхности, стрижи могут только ползать и испытывают трудности при взлёте с неё[8]. На ногах стрижей четыре пальца, единственное исключение составляет Aerodramus papuensis, утративший один палец в ходе эволюции[15]. У саланган, колючехвостов и представителей подсемейства Cypseloidinae лапы анизодактильные — большой палец направлен назад, а остальные вперёд[15][19]. Исследования американского орнитолога Чарльза Коллинза (Charles T. Collins) показали, что у остальных стрижей большой и внутренний пальцы направлены медиально или слегка друг от друга и противостоят центральному и внешнему пальцам, направленным вбок. Похожее расположение пальцев встречается у хамелеонов (Chamaeleonidae) и коал (Phascolarctos cinereus), хотя у них по пять пальцев[15][19].
Оперение
Оперение стрижей плотное, в основном в чёрной или коричневой цветовой гамме, может сочетать чёрные и белые цвета. Исключение составляют близкородственные тропические виды красношейный американский стриж[3] (Streptoprocne rutila) и венесуэльский стрижик (Streptoprocne phelpsi) с ярко-рыжим горлом, подбородком и ошейником у всех взрослых самцов и некоторых взрослых самок[4][15]. В середине XX века яркое оперение считалось признаком полового диморфизма, однако позднее было показано, что оно является возрастной характеристикой, так как самки приобретают его позднее[20]. У стрижей отсутствуют украшающие перья. Спинная птерилия обычно раздваивается, иногда может раздваиваться и брюшная птерилия[4].
Свежее, сразу после линьки, оперение многих стрижей обязательно блестит синим, зелёным или фиолетовым металлическим блеском[4]. Наибольшим блеском обладают перья филиппинского иглохвоста[3] (Mearnsia picina), фиолетового колючехвоста, колючехвостого стрижа (Hirundapus caudacutus) и белобрюхой саланганы[3] (Collocalia esculenta)[15]. Причины этой особенности остаются неясными: блеск может быть связан с защитой от постоянного воздействия прямых солнечных лучей во время полёта, а может оказывать влияние на терморегуляцию или аэродинамику[15].

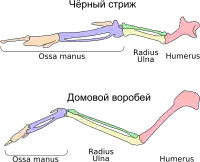
По сравнению с другими птицами, стрижи легко узнаваемы по характерному длинному крылу саблевидной формы. Оно состоит из девяти или десяти длинных первостепенных маховых перьев, за которыми следуют от восьми до одиннадцати очень коротких второстепенных маховых перьев[15][17] и два или три пера крылышка[17]. Длина самого длинного первостепенного пера может в три раза превышать длину самого короткого второстепенного[15][17]. Для сравнения, у ласточек (Hirundinidae) это соотношение обычно не превышает двух. Такое строение крыла связано с тем, что длина пряжки (части кистевого сустава, к которой прикреплены первостепенные маховые перья) значительно превышает длину лучевой и локтевой костей[15]. Роговой ноготок на первом пальце крыла, вероятно, достался от предков[18].
Хвост стрижей обычно довольно короткий, вильчатый или обрезанный прямо[4]. Он состоит из десяти рулевых перьев[15][17], за исключением двенадцати рулевых перьев у чёрного стрижа (Apus apus). Для многих типичных стрижей (Apodini) характерна глубокая вилочка на хвосте, в то время как у саланган имеется только небольшой разрез. Некоторые виды обладают широкой хвостовой вилкой за счёт расширения внешних перьев, это относится к вилохвостым стрижам (Panyptila), карликовым стрижам (Tachornis) и пальмовым стрижам (Cypsiurus). У колючехвостов оси рулевых перьев выступают за их опахала, образуя характерные шипы, из-за которых они получили своё название. Строением хвоста эти стрижи напоминают дятловых (Picidae) и пищуховых (Certhiidae), которые используют его для поддержки на вертикальных поверхностях. В меньшей степени такая особенность свойственна представителям подсемейства Cypseloidinae. У некоторых колючехвостов, в частности короткохвостого иглохвоста[3] (Chaetura brachyura) и иглохвоста — летучей мыши[3] (Neafrapus boehmi), хвост может быть очень коротким, их манёвренность, вероятно, достигается за счёт более широких второстепенных маховых перьев[15].
У многих перелётных стрижей линька начинается до начала миграции на зимние квартиры, приостанавливается во время неё и завершается после её окончания[15][21]. Исключение составляет дымчатый иглохвост (Chaetura pelagica), линька которого заканчивается в начале октября, в самом начале миграции. Молодые особи чёрного стрижа сохраняют часть своего оперения до второй зимовки, используя некоторые перья до полутора лет, взрослые же особи не всегда меняют внешнее первостепенное маховое перо, срок службы которого может достигать двух с половиной лет, при этом последующая линька начинается именно с этого пера. Некоторые стрижи могут линять в период размножения, особенно это относится к иглохвостам (Chaetura), а также к белобрюхому мохноногому стрижу[3] (Aeronautes saxatalis). Оседлые стрижи, как правило, начинают линьку уже после завершения сезона размножения[15]. Во время линьки у стрижей может меняться форма крыла, в частности заметна разница в длине девятого и десятого первостепенных маховых перьев, а также форма хвоста, особенно у птиц с глубоко вильчатым хвостом[21].
Поведение
Вокализация и эхолокация
_hunting_by_echolocation.JPG.webp)
Стрижи — очень шумные, крикливые птицы, демонстрирующие наиболее широкое разнообразие звуковых сигналов во время сезона размножения, при этом вокализации представителей семейства часто схожи между собой[22]. Громкие крики стрижей, летающих в стаях около гнездовых колоний, являются одной из их отличительных особенностей. Сигналом к такому поведению служат повторяющиеся несколько раз позывки приближающегося к колонии стрижа. Крики быстро охватывают всю колонию, и многие птицы оставляют гнёзда, чтобы присоединиться к стае. Стрижи особенно шумны по вечерам[23].
.jpg.webp)
Пары чёрных стрижей издают «swee-ree», при этом исследования австрийского биолога Эриха Кайзера (Erich Kaiser) показали, что самцы издают звук «ree», а самки — «swee». По мнению учёного, полный крик позволяет другим птицам понять, что около гнезда присутствуют оба родителя, а значит потенциальный хищник, желающий добраться до него, будет атакован. Кайзер также предположил, что подобное разделение свойственно и другим представителям семейства. Звуковые сигналы, видимо, важны для определения пола птиц, особенно строящих гнёзда в темноте, а также для формирования пары[22].
Многие стрижи способны к эхолокации[8][15][22]. Не обладая высокой эхолокационной остротой летучих мышей (Chiroptera), необходимой им для ловли насекомых, стрижи используют данную способность для того, чтобы строить гнёзда в тёмных пещерах, возвращаться в них после наступления темноты[15], а также питаться позже многих других видов птиц. Последнее, видимо, позволяет стрижам конкурировать с бульдоговыми летучими мышами (Molossidae), так как представители этого семейства плохо представлены в ареале способных к эхолокации стрижей[22].
Обычно эхолокационные сигналы стрижей расположены в пределах спектра, доступного для человеческого уха, и напоминают звук от проведения предметом по расчёске[22]. Согласно исследованиям лорда Медуэйя и Джона Дэвида Пайя, большинство способных к эхолокации стрижей для ориентации совершают двойные щелчки, в то время как Aerodramus sawtelli и по крайней мере Aerodramus maximus lowi совершают одиночные щелчки. При этом Чарльз Коллинз и Роберт Кашмен Мерфи предположили, что необходимым дополнением к эхолокационным способностям стрижей является хорошо развитая пространственная память[15].
Большинство способных к эхолокации стрижей включены в трибу саланган. Учёные предложили выделить их внутри трибы в отдельный род Aerodramus[15], но возможно, что эта способность распространяется за её пределы. Звуковые сигналы, напоминающие эхолокационные, издают белогорлый американский стриж[3] (Cypseloides cryptus) и пятнистолобый американский стриж, а также плохо изученный мышиный стриж[3] (Schoutedenapus myoptilus). Среди птиц, кроме стрижей, эхолокация встречается только у гуахаро (Steatornis caripensis)[22].
Различные звуки может издавать оперение стрижей. Звуки, напоминающие барабанную дробь, возникают из-за вибрации перьев, V-образный полёт чёрных стрижей, во время которого оба крыла подняты высоко над телом, создаёт короткое «trrr…t», а хвостовые перья чешуйчатого стрижа[3] (Tachymarptis aequatorialis) так широко раскрываются по горизонтали, что внешние рулевые перья почти касаются крыла, и во время вибрации слышен звук «prrrpt-prrrpt-prrrpt»[22].
Полёт

Среди всех птиц стрижи больше всех приспособлены к полёту[18][23][24], в воздухе они проводят очень много времени[18]: питаются, пьют воду, купаются, а некоторые виды также спят и совокупляются[25]. Так, согласно последним исследованиям, чёрный стриж может оставаться в воздухе не приземляясь на протяжении большей части года (за исключением двух месяцев брачного периода)[26].
Стрижи уступают ласточкам в манёвренности и не способны летать с малой скоростью. Техника полёта стрижей зависит от их массы тела и длины крыла, она может различаться в зависимости от целей полёта, из-за чего птиц трудно идентифицировать в воздухе. При этом, если в небе одновременно наблюдается несколько видов стрижей, то можно отличить более тяжёлого и ширококрылого бледного стрижа (Apus pallidus) с коротким хвостом от лёгкого тонкокрылого чёрного стрижа со значительно более глубокой вилкой на хвосте. Также в полёте можно отличить прямолетящую салангану Hydrochous gigas от значительно более мелких саланган, которые в полёте двигаются из стороны в сторону[24].
Сильная горизонтальная тяга, которой объясняется высокая скорость стрижей, создаётся за счёт первостепенных и второстепенных маховых перьев, при этом длинные первостепенные маховые перья обеспечивают также нисходящую силу, а второстепенные и внутренние первостепенные маховые перья — подъёмную силу[15][24]. Взмахи крыльев в полёте довольно жёсткие и быстрые, от четырёх до восьми ударов в секунду[25]. За ними следует скольжение на распростёртых неподвижных крыльях[18]. У стрижей зафиксированы низкие показатели метаболизма в полёте, им не требуются мощные грудные мышцы, соотношение веса груди к общей массе тела составляет 15,5 % у белобрюхого стрижа (Tachymarptis melba) и 19,5 % у бледного стрижа, в то время как у голубиных (Columbidae) — типичных птиц, хлопающих крыльями, — 26,5 %. Стрижи хорошо адаптированы к жизни на больших высотах. Оптимальная доставка кислорода в условиях его низкой концентрации осуществляется за счёт сенсибилизации гемоглобина. Кислородному обмену способствуют бо́льшие, чем у других птиц, размеры эритроцитов[15].
Стрижи считаются самыми быстрыми из маленьких птиц, они могут регулярно летать со скоростью 110 км/час[25], такие показатели были зафиксированы с помощью доплеровского радара у дымчатого иглохвоста во время захода в пещеры в сезон размножения. Более крупные виды стрижей ещё быстрее, скорость колючехвостого стрижа достигала 170 км/час[18][15]. Информация о троекратно превышающей скорости — до 300 км/ч в пике[18] — не была подтверждена. Единственными хищниками, способными поймать стрижей в полёте, являются некоторые крупные соколы (Falco)[25].
Утверждение, что стрижи спят в воздухе, было подтверждено только для чёрного стрижа. Предположительно, оно может быть верно и для бледного и белобрюхого стрижей, так как эти виды обладают исключительной способностью собирать кислород. Концентрация гемоглобина в крови у них очень высока, и сродни показателям, которые были зафиксированы у колибри, обитающих на высоте более 2500 метров. Часто такое поведение наблюдается у молодых птиц, которые занимают воздушное пространство на высоте 1000—2000 м, иногда до 3000 м, и поддерживают высоту редкими взмахами крыльев и скольжением по ветру, обеспечивающему подъёмную силу[23]. Скорость полёта при этом составляет 40—60 км/час[18].
Отдых
Ночью стрижи отдыхают. Птицы спят некрепко, в некоторых колониях всю ночь раздаётся стук. Было замечено, что в плохую погоду птицы обычно устраиваются на ночлег в потенциальных местах гнездования. Чёрный стриж присаживается на стены, в дупла и на стволы деревьев, а также на листья[23]. Чтобы сохранить тепло, птицы могут распушать перья, изгибать спину и вставать друг на друга[23][27], белобрюхий стриж собирается в небольшие плотные группы, а иглохвостый стриж (Hirundapus caucacutus) прячется в дуплах деревьев. Птицы могут только касаться друг друга, как это делает дымчатый иглохвост, или забираться на спину, что свойственно серобрюхому иглохвосту[3] (Chaetura vauxi)[23]. Дымчатый иглохвост в Северной Америке может отдыхать в гнёздах обыкновенного скворца (Sturnus vulgaris) и обыкновенного гракла (Quiscalus quiscula). Иногда птицы формируют довольно крупные группы, в частности во время заморозков в Марокко было зафиксировано три плотных скопления белобрюхих стрижей размером до 350 особей, уцепившихся за часть городской стены[23][27]. Одни виды стрижей находятся рядом с гнёздами круглый год, другие формируют группы около гнезда уже после сезона размножения. В целом для стрижей продолжительность времени, проведённого в гнезде, зависит от климатических и географических условий, возраста птиц, поведения во время сезона размножения, а также необходимости миграции[23].

Из-за суточного распределения плотности насекомых стрижи обычно просыпаются довольно поздно, позже восхода Солнца, при этом северные популяции стрижей поднимаются намного позже, чем южные. Более раннее начало дня может быть связано с хорошей погодой и с отсутствием молодняка в колонии. Малый стриж (Apus affinis) обычно питается утром и вечером, укрываясь во время жаркого дня в гнёздах, при этом птицы остаются в плохо освещённых колониях в среднем на 10—13 минут дольше, чем в хорошо освещённых. В холодные или дождливые дни, а также в дни со слабым ветром, дымчатые иглохвосты дольше остаются в укрытии и раньше возвращаются в него. В укрытии может собираться до 300 птиц[23].
Около водопадов встречаются смешанные гнездовые скопления, характерные в основном для представителей подсемейства Cypseloidinae[23][27]. Пары белогорлого американского стрижа и ошейникового американского стрижа[3] (Streptoprocne zonaris) могут купаться под брызгами воды возле гнезда, при этом последние не дают приземлиться рядом другим птицам, поднимая одно или оба крыла. Пятнистолобый американский стриж может цепляться за выступ скалы со свободно свивающими крыльями, при этом вторая птица цепляется за плечи и верхнюю часть спины и связывается с первой хвостом. Такое поведение наблюдали Мануэль Марин (Manuel Marin) и Фрэнк Гарфилд Стайлс[23].
Белобрюхий мохноногий стриж и, видимо, колючехвостый стриж могут впадать в оцепенение. В течение двух ночных часов температура последних падала с 38,5 ºC до 28 ºC, оставаясь всего на 3 градуса выше температуры окружающей среды, и за несколько утренних минут возвращалась к прежним показателям. Их состояние при этом схоже с настоящими козодоями (Caprimulgidae), в частности американским белогорлым козодоем (Phalaenoptilus nuttallii). Сильные колебания температуры тела зафиксированы и для других стрижей[23][27].
Распространение
Ареал
Стрижи обитают практически повсеместно, в любых регионах, которые могут предоставить необходимые условия для их пропитания[25][28]. Исключение составляют полярные районы (Арктика, Антарктика, Гренландия, Исландия и север Сибири[29]), южная часть Чили и Аргентины, Новая Зеландия, почти вся Австралия[25], некоторые группы островов[28], а также центральные районы Сахары и Тибета[29]. Высокому распространению стрижей способствует их чрезвычайная мобильность, некоторые стрижи обитают на океанических островах, которые никогда не были частью континентальной суши[28].
Стрижи могут расширять ареал, совершая кочевые перемещения. Некоторые кочёвки приводят к быстрой колонизации новых больших регионов, в частности белогузый стриж[3] (Apus caffer) в Испании, бледный стриж на юге Европы и пальмовый стриж[3] (Cypsiurus parvus) на юге Африки. Другие — остаются предметом сложных региональных теорий: чёрный стриж и его предки осуществляли миграцию через Канарские острова ещё в четвертичном периоде, при этом некоторые особи оставались на островах достаточно долго, чтобы вывести потомство. И уже естественный отбор на изолированном участке объясняет морфологические различия между одноцветным стрижом (Apus unicolor), размножающимся на Канарских островах и на Мадейре, и чёрным стрижом. Подобная эволюция происходила у гавайской казарки (Branta sandvicensis). В последние годы на островах стали появляться и гнёзда чёрного стрижа[28].
Для многих видов стрижей доступны кочёвки через Атлантический океан как в западном направлении, так и в восточном. В Карибском бассейне несколько раз был зафиксирован белобрюхий стриж, в Европе, Гренландии и на Тенерифе — дымчатый иглохвост, а в Европе, на Алеутских и Сейшельских Островах — колючехвостый стриж, на островах Прибылова около Аляски и на острове Принца Эдуарда — континентальный чёрный стриж, а на Аляске и в Великобритании — белопоясный стриж (Apus pacificus), обитающий в восточной Азии. Серобрюхий иглохвост, обитающий на западе Северной Америки, несколько раз был замечен на восточном побережье, пепельнохвостый иглохвост[3] (Chaetura andrei) — на Фолклендских островах, чёрный американский стриж[3] (Cypseloides niger) — в Массачусетсе, белогузый стриж с южного Средиземноморья был отмечен в Скандинавии, а белобрюхий стриж — на Британских островах. Кочёвки могут осуществлять и островные стрижи. Антильский карликовый стриж[3] (Tachornis phoenicobia), в частности, был отмечен во Флориде[28].
Среда обитания

Среда обитания стрижей самая разнообразная. Они населяют горы и равнины, открытые пространства и леса, гнездятся на скалах, в пещерах, в норах или в дуплах. Освоили ниши зданий, чердаки и дымоходы[8].
Стрижи обитают в местах с достаточной концентрацией насекомых в воздухе и доступными местами для ночлега и размножения. Зачастую такие условия требуют продолжительных ежедневных перемещений. Птицы, которые откладывают много яиц, добывают корм ближе к гнезду, а птицы с меньшими кладками могут улетать за кормом на большие расстояния. Широкие возможности в этом плане демонстрирует ошейниковый американский стриж. В окрестностях национального парка Анри Питтье на севере Венесуэлы эти птицы могут кормиться в нескольких километрах от высокогорных районов, в которых они строят гнёзда. В случае дождей они могут добывать пищу над улицами близлежащих городов[30]. В районе озёр Рифтовой долины в Кении десятки тысяч особей бурого[3] (Apus niansae) и чешуйчатого стрижа кормятся днём над озёрами, а вечером возвращаются на ночлег в относительно сухую саванну около ущелья Ворота Ада[28].
Скопления насекомых, а за ними и стрижей, обычно встречаются около воды. Особенно зависимы от неё представители подсемейства Cypseloidinae и Hydrochous gigas. При этом бледный стриж может иногда проникать в засушливые районы, встречаясь в оазисах в центре Сахары, а белобрюхий мохноногий стриж размножается в засушливых районах на юге США и на севере Мексики, но добывает пищу как правило на влажных луговых угодьях неподалёку[30].
Более специфические требования к среде обитания предъявляют пальмовые и карликовые стрижи, которые живут на пальмах и редко улетают далеко от них, или иглохвост — летучая мышь, который встречается около баобабов (Adansonia digitata). Некоторые стрижи, включая болотного иглохвоста (Rhaphidura sabini), обитают исключительно в реликтовых лесах[30].
Миграция и перемещения
Большинство стрижей, размножающихся за пределами тропиков, мигрируют на значительные расстояния[28][29]. Некоторые виды стрижей, в частности чёрный стриж, образуют большие колонии перед миграцией, часто смешанные. В Портленде в штате Орегон была зафиксирована осенняя колония серобрюхого иглохвоста размером 20—30 тысяч особей. Такое скопление помогает сохранить запасы жира перед миграцией и способствует линьке. Во время подготовки к осенней миграции дымчатый иглохвост может увеличить свою массу на 51 %. Вместе с тем, фактическая миграция осуществляется в небольших группах[23].
Время миграции стрижей зависит от количества корма, а значит и от погоды. Ярким примером является миграция из Финляндии осенью 1986 года, когда в середине сентября около двух тысяч взрослых птиц вернулись обратно, а затем погибли от голода или с приходом ноябрьских морозов. Предположительно, плохая погода не позволила птицам набрать достаточное количества жира чтобы совершить перелёт на юг. Аналогично, на островах Сисне у побережья Гондураса на протяжении октябрьской недели погибло от голода 200—300 дымчатых иглохвостов[28].
.jpg.webp)
Птицы обычно мигрируют очень быстро. Зафиксированы перелёты чёрного стрижа на 1275 км за четыре дня и белобрюхого стрижа на 1620 км за три дня. При этом птицы могут мигрировать широким фронтом, используя миграционные коридоры только над морем. В частности, чёрный американский стриж во время миграции с Северной Америки в Южную срезает расстояния, пролетая над восточной частью Тихого океана, а дымчатый иглохвост — над Карибским морем. Во время миграции птицы летят очень высоко, с земли их можно увидеть только если плохие погодные условия заставляют стаи снизить высоту. Некоторые птицы могут лететь и ночью. Чёрный стриж, достигнув районов к югу от Сахары, не останавливается на зимовку, а продолжает свои кочёвки по континенту[28].
Помимо ежегодных миграций, птицы могут перемещаться от неблагоприятных погодных условий. Стаи чёрного стрижа летят впереди штормовых фронтов, потом пересекают их и возвращаются на прежние места уже за фронтом. При этом птицы пользуются изобилием воздушного планктона, вызванным конвекцией воздуха во время шторма. Стрижи также могут реагировать на встречный холодный фронт, даже на большом расстоянии. В северном полушарии они кружат по часовой стрелке в тёплых районах к югу или юго-западу от зоны низкого давления и покидают территорию с сокращением числа насекомых. Десятки тысяч чёрных стрижей, следуя зонам низкого давления, могут преодолевать расстояния до двух тысяч километров, так что птицы из южной Англии кормятся в Германии. Во время сезона размножения такие стаи состоят в основном из птиц, которые не сделали кладки, но в них могут участвовать и родители, которые оставляют птенцов на некоторое время без внимания и пищи. Аналогичные полёты в Северной Америке осуществляет чёрный американский стриж[28].
В тропиках после сезона размножения следуют высотные перемещения и распределение птиц. Ошейниковый американский стриж и африканские виды путешествуют по саванне во время засухи, следуя за насекомыми, потревоженными пожарами[28].
Питание
Рацион
Основу питания всех стрижей составляют насекомые и пауки, которых птицы добывают исключительно на лету[31]. В основном это «воздушный планктон» — плохо летающие мелкие насекомые, пассивно дрейфующие в воздушных потоках, или мелкие паучки, использующие паутинки для расселения[18]. Рацион стрижей не имеет точного соответствия составу насекомых в местности — в разных условиях основную часть корма составляют представители разных отрядов. Анализ содержимого желудков и болюсов показал, что его основу составляют перепончатокрылые (Hymenoptera), в частности пчёлы, осы, муравьи, двукрылые, полужесткокрылые, жесткокрылые. Обычно птицы избегают жалящих насекомых, но питаются насекомыми, которые мимикрируют им. Известно, что чёрный стриж в Демократической Республике Конго мог питаться в основном безжальными пчёлами Meliponini. Однако, в ходе другого исследования были обнаружены особи белобрюхого стрижа с укусами на горле[31].
Согласно исследованиям чёрного стрижа, его добычей главным образом становятся равнокрылые хоботные (Homoptera) и относительно реже двукрылые[32]. По наблюдениям в корме чёрного стрижа преобладают тли (Aphididae) и двукрылые[33][34]. Имеются сведения о погадках, собранных около гнёзда чёрного стрижа, которые состояли на 40 % из перепончатокрылых и на 40 % из жуков[35].
Распределение пищевых ресурсов связано с размерами птиц и, возможно, специфическими предпочтениями в рационе[23]. У мелких стрижей рацион более разнообразен, чем у крупных. В количественном выражении крупные стрижи ловят меньшее количество насекомых, как правило более крупного размера. Например, болюс чёрного стрижа в среднем содержит остатки 300—500 насекомых, в то время как болюс белобрюхого — 156—220 насекомых. Крупные стрижи легче добывают роящихся насекомых в местах концентрации отдельных видов на более обширных территориях, что привело к их более узкой специализации. Например, в желудке ошейникового американского стрижа в Венесуэле было обнаружено 800 крылатых муравьёв, а в желудке белоголового американского стрижа[3] (Streptoprocne semicollaris) — 681 огненный муравей Solenopsis geminata. Мелкие стрижи также могут охотиться на роящихся насекомых, но имеют ограниченные кормовые территории. Обычно их кладки превосходят по размерам кладки крупных стрижей и требуют частого кормления быстро растущего потомства, что не даёт птицам отлетать далеко от гнёзд[31]. Согласно исследованиям американских стрижей (Cypseloides), маленький вид — пятнистолобый американский стриж — в основном ловит насекомых длиной 1—5 мм (87,3 % рациона), средний вид — белогорлый американский стриж — 3—5 мм (41,3 %) и 10—12 мм (38,4 %), самый крупный — чёрный американский стриж — 8—11 мм (82,3 %)[36].

Стрижам свойственны сильные географические различия в рационе. В частности, на северо-западе Италии бледный стриж питается в основном мелкими двукрылыми и перепончатокрылыми, в то время как чёрный стриж — крупной тлёй и жесткокрылыми; в Гибралтаре рацион бледного стрижа в основном включает крупных насекомых; а исследование в Марокко выявило значительную долю пауков, что, впрочем, может быть связано с коротким периодом, во время которого были получены образцы[31]. Некоторые виды стрижей демонстрируют значительные сезонные изменения в рационе. Сокращение массовых скоплений питательных видов насекомых в конце сезона размножения птиц приводит к увеличению доли жуков в рационе чёрного стрижа в это время[31][36]. В связи с тем, что большую часть массы жука составляет твёрдый экзоскелет, он считается менее питательным, чем, например, тля. В частности, болюсы, взятые 30 июля, содержали 13 жуков из 898 насекомых, а 30 августа — 106 жуков из 348 насекомых[36]. Вместе с тем, анализ типов добычи полосатохвостого иглохвоста[3] (Chaetura spinicaudus) в Коста-Рике во влажный и засушливый сезоны не показал значительных различий[31].
Стрижи пьют, собирая воду в подклювье, когда пролетают над поверхностью водоёмов[18][31]. При этом размеры водоёма могут быть очень маленькие, а вместе с водой птицы могут собирать и насекомых[31].
Особенности добычи пропитания
Помимо размеров насекомых, стратификация различных видов стрижей также происходит за счёт выбора различных высот, на которых ловят добычу птицы[31][37]. Во время охоты за насекомыми стрижи используют большое вертикальное пространство — на высотах от 2 м до 5800 м, чаще всего на высоте до 50 м[38][39]. В основном стрижи ловят добычу намного выше полога леса. В Европе верхним пределом является высота около 100 метров, за которой количество насекомых сокращается. Согласно исследованиям, проведённым в Эквадоре, в одном столбе воздуха крупные стрижи охотятся выше, чем мелкие. Аналогичные результаты были получены около пещер Ниах в Малайзии и у водохранилища Кариба в Зимбабве. Некоторые стрижи, в частности папуанский иглохвост[3] (Mearnsia novaeguineae), серогузый иглохвост[3] (Chaetura cinereiventris) и дымчатый иглохвост, ловят насекомых непосредственно над куполом леса, а иногда опускаются ниже и даже снимают насекомых с листьев, хотя такое поведение больше характерно для манёвренных ласточек. Также около купола леса питается белобрюхая салангана, в рацион которой входит много насекомых, недоступных для остальных стрижей[31].
Обычно стрижи кормятся в определённое время. Hydrochous gigas питается в сумерках, малый стриж — утром и вечером, устраивая перерыв в жаркие дневные часы, обитающие в пещерах виды питаются поздно вечером, благодаря способности к эхолокации, а белобрюхий стриж и Aerodramus unicolor добывают насекомых ночью на освещённых участках или в полной темноте. Возможно, ночными охотниками является больше представителей семейства, чем считалось ранее. Известно, что белобрюхий мохноногий стриж в поисках добычи следует за сельскохозяйственной техникой, которая тревожит насекомых, а ошейниковый американский стриж в Южной Америке может охотиться на насекомых, потревоженных сезонными пожарами[31].
Основным методом добычи пропитания является «воздушное траление», когда птицы пролетают через скопления воздушного планктона с открытым ртом[18]. Межвидовая конкуренция стрижей зависит от того, насколько широко они могут открывать рот[31][36]. Датский орнитолог Финн Саломонсен, исследовав эту характеристику у четырёх видов стрижей, обитающих на архипелаге Бисмарка, выяснил, что каждый из них имеет свою кормовую нишу[37]. Вероятно, на ранней стадии эволюции стрижи бросались за добычей с насеста, как мухоловки (Muscicapidae) и древесные стрижи (Hemiprocne), затем стали охотиться в воздухе за отдельными насекомыми, как ласточки, а потом освоили «траление»[18]. Стрижи часто образуют крупные смешанные стаи с другими быстрыми птицами, в том числе ласточками, пользуясь кратковременной концентрацией пищи[23].
Размножение
Сезон размножения стрижей зависит от количества насекомых. В тропиках он обычно совпадает с сезоном дождей, в умеренных широтах, как правило, преобладает летнее размножение[40][41]. Исследования в известняковых пещерах Ниах в Малайзии показали, что большинство особей черногнёздных саланган (Aerodramus maximus) и Aerodramus salangana гнездятся с сентября по апрель. При этом черногнёздные саланганы питаются высоко в небе и чаще испытывают проблемы с регулярным питанием. Яйца и птенцы этого вида долгое время могут обходиться без наседки, несмотря на понижение температуры, а сами птицы, включая птенцов, могут переносить периоды голодания. У белобрюхой саланганы выделяют три сезона размножения[40]. Американские стрижи откладывают яйца через один—три месяца после начала сезона дождей, который приносит материал для гнезда и подходящие условия для его склеивания[40][41]. Представители рода Streptoprocne, напротив, откладывают яйца в конце сухого сезона, чтобы обилие насекомых в начале сезона дождей пришлось на период, когда птенцы уже вылупились[40]. Салангана-водорослеед (Aerodramus fuciphagus) в Пенанге у западного побережья полуострова Малакка, по-видимому, имеет достаточное количество насекомых для пропитания, её птенцы появляются в сухой сезон[40][41].
С наличием насекомых связана и продолжительность сезона размножения. В частности, для чёрного стрижа характерна одна кладка, в то время как обитающий южнее бледный стриж может сделать две кладки. В экваториальных широтах некоторые стрижи могут размножаться круглый год, однако даже в таком климате сроки размножения других видов ограничены[40].
Развитие птенцов также сильно связано с условиями питания и количеством добычи. От них зависят темпы роста и сроки вылета птенцов из гнезда. Мягкий средиземноморский климат подходит для бледных стрижей, которые испытывают проблемы со становлением птенцов на крыло в очень сухую или очень влажную погоду, в то время как птенцы более северного чёрного стрижа более устойчивы к капризам погоды. Исследования в Ниме на юге Франции, включающие анализ корреляции успеха размножения с метеорологическими данными на протяжении 13 лет, показали, что северо-северо-западные ветры оказали намного более сильное негативное влияние на количество корма и на успех размножения чёрного стрижа, чем осадки и температура. На северо-западе Италии ненастная погода привела к тому, что самки бледного стрижа отложили яйца на месяц позже обычного, при этом размер кладки не изменился, такая продолжительная задержка невозможна в более северных районах. У чёрного стрижа, напротив, холодная погода привела к уменьшению размера кладки[40].
До настоящего времени не удаётся искусственно вырастить птенцов стрижей, что говорит о сложности процесса. По всей видимости, он дорого обходится взрослым птицам. Обычно гнездовые обязанности выполняют и самцы, и самки[40].
Брачное поведение
В 1950-х годах Ричард Бобби Фишер (Richard Bobby Fischer) отметил, что брачное поведение стрижей всё ещё остаётся слабо изученным. Согласно его исследованиям дымчатого иглохвоста, брачное поведение можно разделить на две фазы. Когда стрижи только возвращаются на летние квартиры, они собираются группами по 4—7 особей, но не для кормления. Часто эти группы разбиваются на летающие вместе пары, по всей видимости, возможность синхронного полёта в таких парах служит индикатором дальнейшего успеха при размножении. В следующую фазу в течение пяти минут два самца гоняются между препятствиями за самкой, иногда улетая на 150 метров от ближайших гнездовий, изредка один из самцов физически атакует другого. Все птицы при этом громко и пронзительно кричат (ранее полагали, что звуки издаёт только самка). Ошейниковый, белогорлый и красношейный американские стрижи также устраивают погони, при этом иногда, прежде чем присоединиться к стае, самку преследуют три самца. Во время погони при глубоком нырке может формироваться V-образная поза, когда оба крыла птицы высоко подняты над телом. Такая поза особенно выражена в уже сформированных парах, обычно она встречается у догоняющих птиц, но может быть и у лидирующих, реже встречается, если в погоне принимают участие три птицы. У чёрного стрижа крылья встречаются за спиной с заметным хлопком, быстро опускаются, а потом хлопают за спиной снова. К другим особенностям брачного поведения стрижей относятся параллельные полёты пар короткохвостого иглохвоста, которые они совершают с крайне медленными взмахами крыльев и тикающими звуками[23].
Считается, что стрижи спариваются в воздухе, однако до сих пор это не было убедительно доказано. Поведение, которое принимается за воздушное спаривание, было зафиксировано у чёрного стрижа, белобрюхого мохноногого стрижа и некоторых других представителей семейства. Самка чёрного стрижа падает вперёд с высоко поднятыми вверх крыльями, а самец, быстро следуя за ней, пытается подняться на неё в тот момент, когда самка выравнивается в горизонтальном полёте. При этом крылья самца оказываются почти в вертикальном положении, а крылья самки — в горизонтальном с небольшим спуском[40][42]. Пара белобрюхого мохноногого стрижа летит навстречу друг другу, а после объединения несколько секунд «крутит колесо», падая к земле. Пальмовые стрижи выпадают с дерева и демонстрируют спиралевидное вращение, наподобие крылатки, повторяя это несколько раз, однако нет доказательств, что в полёте осуществляется спаривание[40]. Согласно ещё одному описанию, во время спаривания в полёте самец подлетает к самке снизу, переворачивается в воздухе вниз спиной и прикладывается клоакой к клоаке[18]. При этом у многих видов стрижей, включая чёрного, зарегистрировано спаривание в гнезде, что ставит под сомнение гипотезу спаривания в воздухе[40][42]. По мнению Марина и Стайлса, воздушное спаривание влечёт за собой большой риск неудачи и бесплодия яиц, что невыгодно видам с небольшими кладками[40].
Стрижи моногамны[8], пары стрижей сохраняются в течение длительного промежутка времени, при этом наличие пары и связанная с этим возможность раньше отложить яйца не оказывает влияние на успех кладки[40]. Пары формируются во время первого лета птиц[40][42], они могут строить вместе гнездо, но не откладывают яйца. Исследования британского орнитолога Дэвида Лака показали, что пары чёрного стрижа остаются вместе, так как они возвращаются в одно гнездо, при этом около четверти пар делают это в один день, а остальные прилетают с интервалом до 21 дня. На гнезде птицы приветствуют друг друга слегка поднятыми крыльями и короткими вскриками[42]. В тропиках пары стрижей могут оставаться вместе, в том числе на гнезде, в течение всего года. Мигрирующие стрижи ежегодно обновляют гнездо[40].
Гнёзда
_nesting_in_Bukidnon%252C_Mindanao%252C_Philippines_01.jpg.webp)
_on_the_nest.jpg.webp)
Большинство стрижей строят гнёзда в тёмных местах, зачастую в полной темноте, для чего им нужны способности к эхолокации — птицы пролетают значительные расстояния в системах пещер. Обычно у стрижей очень строгие требования к месту для строительства гнезда. У чёрного стрижа они включают наличие воды, недоступность для хищников, темноту, возможность беспрепятственного пролёта непосредственно над гнездом, наличие каменных ниш, на которых можно строить гнездо. У пальмового стрижа дистанция подлёта к гнезду может составлять 40 метров[40]. Колючехвостый стриж, чтобы забраться в гнездо, садится на ствол ниже его, а затем карабкается вверх, опираясь на хвост, словно дятел. У многих видов требования касаются окружающих птиц и растений. Карликовые и пальмовые стрижи зависят от различных видов пальм. Эфиопский стриж[3] (Apus horus) гнездится в колониях щурковых (Meropidae), зимородковых (Alcedinidae) и ласточковых (Hirundinidae). Белогузый стриж и, иногда, домовой стриж[1] (Apus nipalensis) используют старые гнёзда рыжепоясничной ласточки (Hirundo daurica). Чтобы избежать такого захвата, другая ласточка — рыжегрудая (Cecropis semirufa) — редко строит гнёзда на высоте, превышающей 1 метр. Серобрюхий иглохвост в Калифорнии часто строит гнёзда в дуплах хохлатой желны (Dryocopus pileatus)[40]. Чтобы занять подходящее дупло или нишу, стрижи могут разорять гнёзда других птиц, проглатывая яйца и птенцов[18]. При выборе мест гнездования большинство стрижей не избегают соседства других пар своего вида, в результате чего образуются небольшие колонии[43]. В литературе приводятся сведения о том, что многие стрижи постоянно гнездятся на одной и той же территории и по возможности в одном и том же месте[44]. При нехватке подходящих для гнездования мест стрижи Apodinae могут занимать дуплянки или скворечники, в ряде случаев даже с уже находящимися в них гнёздами других птиц. Так имеются сведения о том, как пара чёрных стрижей заняла гнездо домовых воробьёв (Passer domesticus), выбросив насиженную кладку последних[45].
Использование слюны при строительстве гнезда является отличительной чертой большинства представителей подсемейства Apodinae. Стрижи рода Aerodramus используют слюну больше остальных стрижей, при этом салангана-водорослеед делает гнёзда исключительно из слюны. Выработке большого количества слюны способствует значительное увеличение подъязычной железы у птиц обоих полов во время сезона размножения[15][40]. Крепление гнезда к вертикальной стенке также осуществляется при помощи слюны, однако есть и исключения. Большой[3] (Panyptila sanctihieronymi) и малый[3] (Panyptila cayennensis) вилохвостые стрижи из слюны и растений формируют материал, похожий на войлок, из которого создают трубчатые конструкции, свисающие с основания, либо прикреплённые по всей длине к вертикальной поверхности, длина конструкции при этом может достигать 66 см[40]. Яйца они откладывают в карманчиках на внутренней стенке гнезда, которое используют несколько лет, обустраивая каждый раз новый карманчик[46]. Антильский, центральноамериканский[3] (Tachornis furcata) и вилохвостый[3] (Tachornis squamata) карликовые стрижи в Новом Свете строят гнёзда на высушенных пальмовых листьях, покрывая внутреннюю стенку пушистыми семенными коробками, а внешнюю — перьями[46][40], аналогичным образом строят гнёзда пальмовые стрижи[40].
В качестве строительных материалов стрижи могут использовать переносимый по воздуху детрит, который они доставляют к месту строительства гнезда в клюве. Они собирают лапами кору с деревьев, лишайники и печёночные мхи с влажных каменных стен[40], используют веточки, травинки, растительные волокна и перья. Птицы хватают материалы на лету[8], часто крадут у других, в том числе у своих сородичей[40]. У некоторых стрижей размеры гнёзд рекордно малые относительно размеров самих птиц[46].
Кардинально отличаются гнёзда представителей подсемейства Cypseloidinae и, возможно, колючехвостых стрижей (Hirundapus), которые не используют слюну при строительстве гнезда. Ошейниковый американский стриж, белоголовый американский стриж, большой колючехвост (Hirundapus giganteus) и колючехвостый стриж, как и, возможно, менее изученные виды колючехвостых стрижей, обычно выскребают гнездо[40]. Американские стрижи прикрепляют гнёзда в скалах у воды, и часто, чтобы добраться до ниши с гнездом, они должны пролететь под водопад сквозь толщу воды[46].
Стрижи обычно защищают свои гнёзда. Птицы издают сигнал перед входом в углубление для гнезда. Ответ позволяет определить не только занято ли оно, но и то, сформировалась ли пара. Конфликты с нарушителями границ могут продолжаться часами и иметь фатальные последствия. Одна птица приближается к другой с поднятыми крыльями, слегка наклоняясь в сторону, чтобы обнажить сильные лапы. Слабый, а следовательно малоэффективный клюв в такой борьбе используется редко[40][42]. В столкновении с представителями других видов, чёрный стриж обычно одерживает победу над домовым воробьём (Passer domesticus), но проигрывает в борьбе с обыкновенным скворцом (Sturnus vulgaris). Во время борьбы между белобрюхим стрижом и сизым голубем (Columba livia) два стрижа прижались к голубю сверху и снизу, и вели его к столкновению, вызвав таким образом серьёзные травмы у последнего. Несколько особей небольшой городской ласточки (Delichon urbica) могут объединяться, чтобы прогнать более крупных одноцветных или белобрюхих стрижей[40].
Яйца

Представители семейства откладывают однородные тускло-белые яйца небольшого размера с высоким содержанием желтка, при этом размеры яиц могут составлять от 15,5 × 10 мм у вилохвостого карликового стрижа до 43 × 28,5 мм у ошейникового американского стрижа. Значительно варьирует и размер кладки: от одного яйца у некоторых саланган до семи у иглохвостов и колючехвостых стрижей[40]. Средние размеры первой и второй кладок саланганы-водорослееда не отличаются друг от друга, первая кладка бледного стрижа в Тунисе в среднем состоит из 2,89 яиц, а вторая — 1,95[41]. Размер яиц в кладке хорошо коррелирует с развитием птенцов, птенцы из более крупных яиц быстрее встают на крыло. Возможно, это связано с оптимизацией конкуренции между птенцами в случае непогоды[40].
Некоторые стрижи приклеивают яйца к гнезду. Самка пальмового стрижа прижимает яйцо к стороне гнезда, и наносит слюну на гнездо рядом с яйцом чтобы скрепить их, поскольку яйца откладываются довольно открыто[40]. При насиживании птицы удерживаются, крепко вцепившись в пальмовый лист, качающийся на ветру[46][47]. Возможно, приклеивает яйца к гнезду и вилохвостый карликовый стриж[40].
Стрижи обычно откладывают яйца с интервалом в 2—3 дня. Инкубационный период составляет 17—25 дней и начинается с первого яйца[46], у некоторых видов превышает 30 дней[47]. Насиживают яйца оба родителя[46][47]. Промежутки времени могут составлять от двух минут до почти шести часов, за десять часов родители могут смениться два раза или двенадцать. Иногда яйца остаются без присмотра до шести с половиной часов, причём такое поведение наблюдалось не только в тропическом климате[47]. Aerodramus terraereginae выработал уникальную стратегию инкубации, связанную с ограниченными и изменчивыми запасами продовольствия в окрестностях Чиллаго в Квинсленде, Австралия. Самка откладывает два яйца за сезон, при этом второе яйцо откладывается через 50 дней после первого и инкубируется птенцом первой кладки. Первый птенец использует инкубационную позу, аналогичную взрослой птице, возможно, из-за тесного гнезда. Если первый птенец вылетал из гнезда раньше появления второго, насиживание продолжали уже взрослые птицы, но время второй кладки выбрано так, чтобы этого почти не случалось. При этом не было замечено значительной разницы между периодами инкубации первого и второго яйца. Таким образом, общее время инкубации сокращается на три недели[40].
У некоторых иглохвостов и у белогузой саланганы[3] (Aerodramus spodiopygius), которые делают большие кладки, часто в одном гнезде могут быть яйца разных самок. Было показано, что у дымчатого иглохвоста в насиживании и кормлении участвует третий взрослый, причём делает это чаще родителей. Обычно у гнезда обнаруживают трёх взрослых птиц, но в исследовании американского орнитолога Ральфа Уоррена Декстера (1912—1991) в 6 из 22 случаев было зафиксировано четыре взрослые птицы, обслуживающие гнездо. Помощниками являются молодые или очень старые птицы, чаще самцы, чем самки. Молодые птицы ещё не умеют защищать гнездо и им требуется безопасное место для ночлега[40].
Птенцы


Вылупившиеся птенцы стрижей обычно слепые и голые[40][46], но вскоре они покрываются густым белым или серым пухом[46]. Представители подсемейства Cypseloidinae и трибы типичных стрижей покрыты пухом с рождения[40]. Наличие пуха у птенцов может быть связано с открытыми гнёздами[46][40], когда температура колеблется от 15 °C до 41 °C[40]. Чтобы уменьшить риск падения из открытого гнезда, птенцы крепко вцепляются в субстрат когтями[46]. Птенцы пальмовых стрижей сразу после появления ведут себя очень беспокойно, а на четвёртый день они выбираются из гнездовой камеры на пальмовую ветвь[40].
Взрослые птицы кормят птенцов насекомыми, которых в большом количестве приносят в подъязычном мешке. Кормление происходит редко[46], иногда родители покидают гнездо на несколько дней, а птенцы впадают в оцепенение[47]. Чаще всего большое количество птенцов в гнезде стимулирует более частое питание, однако при этом уменьшается количество пищи. Эксперименты с размерами приплода у чёрных стрижей показали, что несмотря на увеличившуюся из-за большего количества птенцов частоту кормления, общее количество пищи на одного птенца уменьшалось, как и масса взрослых птиц и птенцов. При достижении опасно низкого собственного веса родители переставали кормить птенцов, что позволяло им выжить самим и накормить оставшихся в живых птенцов. В среднем у стрижей частые кормления сокращают период гнездования. Методы кормления птенцов отличаются разнообразием даже в рамках одного вида. Взрослые стрижи отрыгивают болюс с насекомыми непосредственно в горло птенцов или делят между ними[40]. В болюсе чёрного стрижа может содержаться до 2000 насекомых, обычно — 300—500, в то время как в болюсе белобрюхого стрижа — 156—220 насекомых, хотя эта птица крупнее. По мере роста птенцов родители приносят в болюсе более крупных насекомых[36]. Непереваренные остатки насекомых птенцы выбрасывают через рот в виде погадок[46]. Птенцы стрижей часто обладают большим количеством жира, что помогает им выжить при редком кормлении и плохой погоде. Птицы могут испытывать периоды длительного голодания, когда на протяжении нескольких недель получают очень мало пищи. При этом вес птенцов может уменьшаться в два раза[36][40]. В исследовании Лака птенцы чёрного стрижа набирали вес в среднем с 2,75 г до 56 г за четыре недели, после чего теряли вес вместе с ростом перьев. Изменение массы птенцов сильно зависит от погодных условий, десятидневные птенцы одного вида могут весить от 5 до 50 граммов. Такая вариация реже наблюдается в тропическом климате, где колебания температур обычно меньше, хотя было отмечено, что ей чаще подвержены вторые птенцы или птенцы, страдающие от паразитов[41].
Период гнездования стрижей длиннее, чем у других птиц схожего размера, и не связан с размерами самих стрижей. Птенцы чёрного стрижа остаются в гнезде в среднем 42 дня, в плохую погоду — до 56 дней, при этом случаются и преждевременные вылеты из гнезда. Дольше всего в гнезде остаются птенцы пятнистолобого американского стрижа — 65—70 дней в особенно плохую погоду[40][41]. За неделю до вылета из гнезда птицы теряют вес, что помогает им достичь оптимальной нагрузки на крыло, необходимой для быстрого и манёвренного полёта[40]. Перед тем как покинуть гнездо, молодые птицы делают различные упражнения для крыла. Птенцы иглохвоста — летучей мыши, гнёзда которого расположены в глубоких ямах, карабкаются по стене вверх и быстро бьют крыльями, поддерживая себя с помощью хвоста во время частых периодов отдыха. Птенцы могут подниматься над полом с помощью вытянутых вниз крыльев. Предполагается, что птенцы, готовые покинуть гнездо, могут удерживать такую позицию[40].
Вылетевшие из гнезда птенцы полностью независимы[40][46], они умеют летать и не нуждаются в родительской опеке[46], с чем также может быть связана такая продолжительность времени, проводимого птенцами в гнезде[40]. Обычно молодые стрижи не возвращаются в гнездо после того, как первый раз вылетят из него. Тем не менее птенцы некоторых видов ещё неделю после первого полёта отдыхают днём около гнезда, а другие птицы возвращаются в гнездо вместе со взрослыми[23].
Стрижи становятся половозрелыми на второй — третий год[48].
Экология и выживаемость

Стрижи сильно подвержены паразитизму, что скорее всего связано с трудностями очистки в связи с анатомическими особенностями птиц. Многие беспозвоночные паразитируют как на отдельных птицах, так и на гнёздах стрижей. В Африке основными паразитами являются кровососки (Crataerina, Pseudolynchia, Ornithomya), пухопероеды (Dennyus, Eureum) и Ixodida. Некоторые беспозвоночные эндемичны для определённых групп стрижей, что свидетельствует об их коэволюции[23]. Заражение белогузой саланганы вшами Myophthiria fijiarum на Фиджи связано с высокой плотностью гнёзд[40]. В северной части Калимантана яйца и молодые птенцы саланган часто заражены Rhapidophora oophaga[49]. Эксперимент с Crataerina pallida и Dennyus hirundinis в Оксфордском университете показал, что успех кладки не зависит от числа паразитов[40].
Вместе с тем, многие стрижи поддерживают гнёзда в чистоте. Птенцы белогузой саланганы уже с первого дня испражняются с края гнезда, аналогичным образом поступают многие другие птенцы стрижей. Взрослые особи дымчатого иглохвоста могут выбрасывать из гнезда фекалии совсем маленьких птенцов, или глотать их. Яичная скорлупа и мёртвые яйца обычно также выкидываются из гнезда, пальмовые стрижи при этом разбивает скорлупу на мелкие кусочки. Гнёзда чёрных стрижей загрязнены сильнее[40].
На птенцов Aerodramus sawtelli охотятся крабы, которые иногда забираются в гнёзда по стенам пещер, а яйца и птенцы пальмовых стрижей могут стать добычей различных птиц от краснокрылого длиннохвостого скворца (Onychognathus morio) до африканского филина (Bubo africanus)[40][49]. Опасность для стрижей представляют быстрые хищные птицы: сапсан (Falco peregrinus) часто охотится на белобрюхого мохноногого стрижа, чеглок (Falco subbuteo) — на малого стрижа, серебристый чеглок (Falco concolor) — на белобрюхого стрижа, а широкоротый коршун (Macheiramphus alcinus) — на саланган[23][49]. При этом стрижи могут нападать на хищников, быстро маневрируя и издавая тревожные звуковые сигналы[23].
Выживаемость птенцов пальмовых стрижей крайне мала и составляет 26 %, в то время как выживаемость птенцов дымчатых иглохвостов — 96,1 %[40]. После первого года уровень смертности стрижей очень низок. Например, в колонии бледного стрижа на Гибралтаре уровень смертности в первый год составил 67,3 %, а в последующие — 26 %. Показатели смертности чёрного стрижа, дымчатого иглохвоста и красношейного американского стрижа составляют 15—19, 15—17 и 19—29 %, соответственно[40]. Высокая смертность часто связана с плохой погодой, при этом птицы умирают как от холода и дождя, так и от связанного с ними отсутствия корма[49].
Средняя продолжительность жизни в природе составляет 7—10 лет[29]. Рекордный возраст зафиксирован у белобрюхого стрижа — 26 лет[40], чёрного стрижа — 21 год[29] и дымчатого иглохвоста — 14 лет[40].
Взаимоотношение с человеком

Отношения стрижей с человеком крайне противоречивы. С одной стороны, люди разрушают естественную среду обитания, сокращают количество насекомых, используя инсектициды и гербициды, готовят деликатесы из птичьих гнёзд. В то же время, построенные людьми сооружения и высаживаемые пальмы создают дополнительные места для гнездования. Такое гнездование не только увеличивает успех размножения, но и способствует расширению ареала. Некоторые виды стрижей, включая дымчатого иглохвоста, полностью перешли на строительство гнёзд на искусственных сооружениях, за последние 100 лет было обнаружено только десять гнёзд на клёне серебристом (Acer saccharinum). При этом обычно ремонт кровли, установление сеток против насекомых и прочие работы на крышах ведутся без учёта благополучия стрижей и могут служить причиной их гибели[50].
Стрижи, как и прочие насекомоядные птицы, являются естественными регуляторами насекомых. Согласно исследованиям домового стрижа в провинции Юньнань на юге Китая, в диете этого вида 67,5 % насекомых являются сельскохозяйственными вредителями, а 13 % насекомых — полезны. Аналогичные результаты дало исследование белопоясного стрижа в Китае. Малый стриж и азиатский пальмовый стриж[1] (Cypsiurus balasiensis) на юге Индии питаются определёнными видами насекомых-вредителей, но их популяция существенно сократилась из-за большого количества пестицидов[50].
Ласточкины гнёзда


Сделанные полностью из слюны, гнёзда саланганы-водорослееда являются деликатесом в китайской кухне[50]. Ещё три вида саланган производят съедобные гнёзда — съедобная салангана (Aerodramus germani), черногнёздная салангана (Aerodramus maximus) и Aerodramus unicolor. Особенно ценными считаются гнёзда чистого белого цвета без примесей, собранные во время сезона размножения. Основными элементами съедобного птичьего гнезда являются сиаловая кислота — 9 %, галактозамин — 7,2 %, глюкозамин — 5,3 %, галактоза — 16,9 %), фруктоза — 0,7 %, аминокислоты (аспарагиновая, глутаминовая, пролин, треонин, валин) и минеральные соли (в основном натрия и кальция)'[51]. Было высказано предположение, что они способствуют клеточному делению и могут применяться как один из компонентов борьбы с иммунодефицитом при СПИДе[50].
Торговля съедобными гнёздами осуществлялась ещё в XVII веке в конце правления династии Мин и в начале государства Чжэн, высказываются предположения о более ранней традиции — с VI или VIII века. В настоящее время торговля гнёздами является многомиллионным бизнесом. В начале 1990-х годов цена гнёзд составляла 367—611 долларов США за килограмм, гнёзда хорошего качества могли стоить более 1225 долларов. Позднее цена поднялась до 36 тысяч долларов за килограмм. Основная торговля сосредоточена в Гонконге. Согласно некоторым оценкам, Индонезия экспортирует гнёзда на сумму около миллиарда долларов США в год[50].
Один килограмм включает 80—120 гнёзд, только в Индонезии собирают до 200 тонн гнёзд в год. Во время сезона гнёзда собираются дважды, так что для размножения птицы вынуждены их строить в третий раз, при этом из-за сильного истощения слюнных желёз гнёзда содержат кровь. Всё вместе это приводит к сокращению популяции стрижей со съедобными гнёздами. По некоторым оценкам в 1959 году в пещерах Гомантонг в штате Сабах в Малайзии во время сбора гнёзд из них было выкинуто около 190 тысяч яиц и птенцов. В пещерах Ниах численность птиц одно время достигала 4,5 млн, однако в конце 1980 — начале 1990 годов оценивалась в 150—298 тысяч. Несмотря на существенное сокращение гнёзд, в 1995 году доля экспорта съедобных гнёзд в экономике Саравака составила 10 400 000 долларов США[50].
Такой рынок окружает большое количество незаконных операций, включая «узаконенные убийства», которые впоследствии выдаются за самооборону[50]. Сбор съедобных гнёзд в Юго-Восточной Азии часто ведётся с нарушением закона. Даже такие известные и охраняемые места гнездования, как пещеры Ниах, в которых официальный сбор гнёзд был полностью запрещён в 1989—1991 и 1993—1997 годы, привлекают браконьеров, при этом мониторинг меньших пещер, в частности на островах Таиланда в Андаманском море, невозможен. Помимо опасностей, связанных с методикой сбора (люди забираются высоко под свод пещеры по бамбуковым конструкциям), в этих регионах происходят конфликты между сборщиками гнёзд по лицензии и браконьерами[52].
Учёные отмечают, что на смену птицам со съедобными гнёздами приходят белобрюхие саланганы и летучие мыши. В странах Азии начали сооружать конструкции, на которых стрижи могут строить гнёзда. Обычно гнёзда строят белобрюхие саланганы, люди подкладывают в них яйца саланган-водорослеедов, птенцы которых, когда вырастают, создают колонии на этих искусственных сооружениях[52].
Охранный статус

В зависимости от классификации стрижей к охраняемым относят разное количество видов. Международный союз охраны природы к вымирающим относит один вид стрижей, Aerodramus bartschi; к уязвимым — шесть видов, а именно Aerodramus elaphrus, Aerodramus sawtelli, Schoutedenapus schoutedeni (рассматривается Международным союзом орнитологов как подвид Schoutedenapus myoptilus[53]), блестящий стриж[3] (Apus acuticauda), дымчатый иглохвост, чёрный американский стриж. Близки к уязвимому положению пять видов стрижей: маврикийская салангана[3] (Aerodramus francicus), Cypseloides rothschildi, филиппинский иглохвост, Hydrochous gigas, Aerodramus vulcanorum. Ещё по шести видам биологических данных недостаточно: пятнистолобый американский стриж, Cypseloides storeri, Apus sladeniae, филиппинская салангана (Aerodramus whiteheadi), Aerodramus papuensis, Aerodramus orientalis[54].
Многие виды стрижей защищены национальными законами и международными договорами. В частности, одноцветный стриж включён в четвёртую категорию списка «Species of European Conservation Concern», к включению в который также планируются бледный и белобрюхий стрижи. С 1984 года Aerodramus bartschi включён в Красную книгу США. Однако, как показывает ситуация с Aerodramus unicolor на Шри-Ланке, одних законодательных инициатив бывает недостаточно. Ни один из видов, производящих съедобные гнёзда, не является объектом международного права[52].
Человек редко охотится на стрижей. Птиц Hirundapus celebensis убивают на Филиппинах, так как они охотятся около пчелиных ульев, а чёрных стрижей ловят и едят в Малави и Либерии. Кроме того, известно только несколько записей из Демократической Республики Конго и Таиланда об убитых человеком стрижах. Чаще стрижи умирают в результате человеческой деятельности случайно, сталкиваясь с проводами и самолётами, употребляя хлорированную воду из бассейнов, задыхаясь в дымоходах и падая в камины[52][49]. Как и различные хищники, стрижи страдают от высокого уровня пестицидов, который у хищных птиц снижает успех размножения. У Aerodramus bartschi, обитающих только на тихоокеанском острове Гуам, концентрация ДДТ в тканях составила 0,27 на миллион[52].
По всей видимости, сокращаются ареалы некоторых лесных видов стрижей, для строительства гнёзд которым необходимы реликтовые леса, хотя они могут питаться и во вторичном лесу. Капские стрижи[3](Apus barbatus), например, для гнездования и ночлега использует только большие деревья. Кроме того, было высказано предположение, что полная вырубка старого леса в некоторых районах Северной Америки может привести к исчезновению в них серобрюхих иглохвостов[52].
Большое влияние на стрижиных может оказать глобальное изменение климата. Адекватное количество насекомых является обязательным условием успешного размножения и зависит от температуры и осадков. В случае глобального потепления в северные и южные районы могут распространиться традиционно тропические виды: домовой стриж, малый стриж и белогузый стриж. Вместе с тем, отсутствие подробных знаний об изменении численности стрижей во время четвертичного периода не позволяет в полной мере оценить антропогенное влияние на проблему[52].
Эволюция
| Кладограмма стрижеобразных по Ksepka et al.[55] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Самые ранние ископаемые остатки представителей семейства стрижей были найдены в Дании и Германии в 1980-е годы и датируются средним эоценом. Они были описаны как вымерший род Scaniacypselus Harrison, 1984[56][57]. Харрисон также относил к стрижам род Procypseloides Harrison, 1984[56]. Стрижеподобных птиц нижнего эоцена учёные относят к предкам древесных стрижей или совиных козодоев[57]. Уже с верхнего эоцена известны представители современных родов Cypseloides и Chaetura[4].
Самой примитивной группой стрижей, по мнению южноафриканского орнитолога Ричарда Кендалла Брука, который рассмотрел недавнюю эволюцию стрижиных, а также анатомические и морфологические различия и особенности гнездования, является ограниченная Новым светом Cypseloidinae. За ней следуют Collocaliini и Chaeturini, а вершиной эволюции в настоящий момент является группа Apodini. Брук обратил внимание на то, что представители Chaeturini в Старом Свете сильно отличаются друг от друга, имеют меньшее число подвидов и непересекающиеся ареалы, в то время как в Новом Свете птицы сильно похожи друг на друга, обладают большим количеством подвидов и обширными ареалами. По его мнению, это показывает, что птицы распространились со Старого Света в Новый[9][58][59]. Харрисон расположил таксоны по порядку уменьшения длины и усиления крепости локтевой кости следующим образом: Eocypselus — Hemiprocnidae — Cypseloidinae — Apodinae — Chaeturini (в оригинале, Chaeturinae). При этом он отметил, что другие ископаемые остатки не укладываются в эту цепочку[56]. В некоторых исследованиях род Eocypselus считается общим предком как стрижей, так и колибри[55].
Исследования немецкого палеонтолога Геральда Майра подтвердили монофилию современных стрижей на основе сильно сокращённой проксимальной фаланги пальцев и из-за отсутствия плечевой ямки. Он также показал монофилию стрижей вместе с вымершим родом Scaniacypselus[57].
Систематика
Исторический очерк
В десятое издание «Системы природы» Карла Линнея, увидевшее свет в 1758 году и давшее начало современной биологической систематике, было включено четыре вида стрижей, которые были отнесены к ласточкам: Hirundo esculenta (Collocalia esculenta), Hirundo Apus (Apus apus), Hirundo pelagica (Chaetura pelagica) и Hirundo melba (Tachymarptis melba)[53][7]. Уже в работе Скополи 1777 года стрижи выделены в отдельный род Apos[10][12], той же точки зрения придерживался Иллигер (1811)[12]. В 1827 году французский зоолог Фердинанд Жозеф Л’Эрминье расположил отдельное семейство стрижей, включающее также древесных стрижей, между колибри и козодоями. К середине XIX века такая классификация поддерживалась большинством учёных[60], а британский зоолог Филип Склейтер в 1865 году предложил деление стрижей на подсемейства Chaeturinae и Apodinae (в оригинале Cypselinae), основываясь на строении ноги у птиц[59][60]. Подсемейство Cypselinae включало два рода и 19 видов, при этом учёный говорил о возможности дальнейшего разделения, а подсемейство Chaeturinae — четыре рода, включая древесных стрижей и род Collocalia, перечисление видов которого не вошло в описание[60].
В 1892 году немецкий орнитолог Эрнст Хартерт разделил Chaeturinae на три группы: саланганы (Collacalia) с обычными перьями хвоста, колючехвосты (Chaetura) с очень твёрдыми рулевыми перьями с острыми концами и Cypseloides с немного твёрдыми, но не острыми рулевыми перьями[61]. Хартерт полагал, что семейство впервые было описано в работе Склейтера[14], однако в современных источниках описание приписывается ему[62]. В классификации, опубликованной американским орнитологом Джеймсом Ли Питерсом в 1940 году, учёный определил девять родов стрижей подсемейства Chaeturinae. Род Collocalia оставался неизменным, в то время как из рода Chaetura были выделены Hirund-apus, Streptoprocne, Zoonavena, Mearnsia, а из рода Cypseloides — Nephoecetes. Он рассматривал род Aerornis, в который включал два вида — A. senex и A. semicollaris — связующим звеном между родами Cypseloides и Streptoprocne, а стрижей S. rutila относил к Chaetura (Хартерт включал их в Cypseloides). В целом он продолжил классификацию Склейтера[61]. Со временем учёные снова объединили вместе роды Chaetura, Hirund-apus, Zoonavena, Mearnsia, возвращаясь, с небольшими исключениями касающимися отдельных видов, к классификации Хартерта. Также произошло объединение родов Cypseloides, Streptoprocne, Aerornis и Nephoecetes. При этом Лак полагал, что в целом такая классификация продолжает базироваться на признаках, введённых ранее Хартертом. Он считал, что представители рода Chaetura развили свои хвостовые перья в результате специализации, в то время как стрижи рода Cypseloides потеряли бородки на перьях из-за трения[61].
Питерс разделил подсемейство Apodinae на семь родов, включающих 10 видов. При этом, как и у Хартерта, птицы родов Apus, Aeronautes, Panyptila обладали пампродактильным строением ноги, а у родов Tachornis, Cypsiurus, Reinarda пальцы направлены попарно в разных направлениях. К последней группе также относился описанный позже в классификации Хартерта род Micropanyptila. Дальнейшее деление включало противопоставление голых ног и ног, покрытых перьями[61]. Лак полностью отказался от такой классификации, определив в составе подсемейства пять родов, включающих 19 видов, два рода при этом были ограничены Старым Светом, а три — Новым[61].

Критический анализ систематики стрижей опубликовал в 1956 году Лак, который работал над исследованием особенностей гнездования птиц. Вслед за ним бельгийский натуралист Антон Де Ро в 1968 году подставил под сомнение деление стрижей на подсемейства Chaeturinae и Apodinae, выделив род Scoutedenapus, представители которого, в частности S. myoplitus и S. schoutedeni, внешним видом напоминают типичных стрижей, а анизодактильным строением ног — иглохвостов (строение ног было основной характеристикой, различающей роды Chaetura и Apus). На основании подробной работы Лака, которая включает и другие характеристики стрижей, Брук предложил разделить стрижей на подсемейство Cypseloidinae, которое включало объединённых Лаком в один род птиц, и подсемейство Apodinae, которое включало всех остальных стрижей. Брук отнёс род Schoutedenapus к подсемейству Apodinae. Учёный не делал подробный анализ представителей этого рода, но высказал предположение на основе строения хвоста с характерной глубокой вилкой — особенность, присущая также Apus, Cypsiurus, Panyphila и Tachornis. Он же предложил разделить подсемейство Apodinae на три трибы[59].
Многие характеристики, используемые Бруком для определения подсемейств и триб, в дальнейшем были поставлены под сомнение, однако само деление на подсемейства Cypseloidinae и Apodinae сохранилось[20].
Современная систематика
Международный союз орнитологов относит стрижей к отряду стрижеобразных, который традиционно включает настоящих стрижей, древесных стрижей и колибри. Некоторые учёные относят к стрижеобразным совиных козодоев, сохраняя при этом монофилию отряда[57][63], другие включают все указанные выше семейства к обширному отряду козодоеобразных[64].
Общепризнанным является выделение двух подсемейств настоящих стрижей: Cypseloidinae и Apodinae[17][29], включающих 19 родов[29][53]. К подсемейству Cypseloidinae относят роды Cypseloides и Streptoprocne, включающие наиболее примитивные американские виды[9][29][58], представители которых обладают двумя сонными артериями, примитивным нёбом[58][9] и острым и твёрдым хвостом[25]. Кроме того, они не используют слюну при строительстве гнезда. Представители подсемейства Apodinae, напротив, используют слюну при строительстве гнезда (исключение составляет Hirundapus), а также обладают одной сонной артерией, хорошо развитым нёбом[9] и мягким хвостом[25].
Многие учёные делят подсемейство Apodinae на три трибы[9][17][29]:
- Саланганы (Collocaliini) включает роды Hydrochous, Collocalia (саланганы), Aerodramus и Schoutedenapus;
- Колючехвосты[29] (Chaeturini) включает роды Mearnsia, Zoonavena, Telacanthura, Rhaphidura, Neafrapus, Hirundapus (колючехвостые стрижи) и Chaetura (иглохвосты, или иглохвостые стрижи);
- Типичные стрижи[29] (Apodini) включает роды Aeronautes (мохноногие стрижи), Tachornis (карликовые стрижи[3]), Panyptila (вилохвостые, или кайенские, стрижи), Cypsiurus (пальмовые стрижи), Tachymarptis и Apus (стрижи, или обыкновенные стрижи).
Трибу колючехвостов иногда выделяют в отдельное подсемейство Chaeturinae[17]. Положение Schoutedenapus также остаётся неопределённым[9][58], американский орнитолог Марк Холмгрен (Mark Holmgren) отнёс его к трибе типичных стрижей. Он также считал, что древесные стрижи не должны рассматриваться как отдельное семейство, их следует считать трибой Hemiprocnini семейства стрижиных[9].
Проблемы систематики стрижей связаны также с тем, что роль морфологического анализа сильно преувеличена. В частности, изучение морфологии гнёзд стрижей не способствует их классификации[9]. Наибольшие сложности возникают во время морфологического анализа саланган. Питерс назвал их одной из самых сложных групп птиц для классификации[9][58]. В 1970 году Брук разделил род на три части: Hydrochous, Collocalia, Aerodramus. По Бруку Hydrochous включает Hydrochous gigas, Collocalia — блестящих стрижей не способных к эхолокации, а Aerodramus — неблестящих стрижей способных к эхолокации. При этом согласованность между молекулярной филогенией и эхолокацией выше, чем между ней и гнездовым поведением. Кроме того, на всех трёх группах и только на них паразитирует один род вшей. Вместе с тем, сравнение цитохром b показывает, что Collocaliini не является монофилетической трибой[9]. В то же время, хвостовые шипы Cypseloidinae, ранее считавшиеся филогенетической особенностью, являются функцией размера тела и особенностей присаживания птиц на поверхность[9].
Учёные сходятся во мнении, что для достижения реального консенсуса по вопросам систематики стрижеобразных все таксоны должны быть полностью проанализированы с использованием всех доступных методов[9].
Примечания
- Глущенко Ю. Н., Коблик Е. А., Архипов В. Ю., Глущенко В. П., Елисеев С. Л., Коробов Д. В., Коробова И. Н., Логинов Н. Г., Малых И. М., Семёнов Г. А., Симонов В. А., Хайдаров Д. Р., Чернышёв О. Г. Орнитологические наблюдения в Таиланде в 2006—2018 годах // Русский орнитологический журнал. — 2018. — Т. 27, вып. 1627. — С. 2960.
- база данных «Птицы Российской федерации» Архивная копия от 25 ноября 2020 на Wayback Machine на сайте Института проблем экологии и эволюции имени А. Н. Северцова
- Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Русский язык, РУССО, 1994. — С. 151—154. — 2030 экз. — ISBN 5-200-00643-0.
- Коблик, 2001, с. 54.
- Галушин В. М., Дроздов Н. Н., Ильичёв В. Д., Константинов В. М., Курочкин Е. Н., Полозов С. А., Потапов Р. Л., Флинт В. Е., Фомин В. Е. Фауна Мира: Птицы: Справочник. — М.: Агропромиздат, 1991. — С. 178—180. — 311 с. — ISBN 5-10-001229-3.
- Jobling J. A. The Helm Dictionary of Scientific Bird Names. — London: A&C Black Publishers Ltd, 2010. — P. 51—52. — 432 p. — ISBN 978-1-4081-2501-4.
- Linnaeus C. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. — Editio decima, reformata. — Stockholm: Holmiae. (Laurentii Salvii), 1758. — Vol. 1. — P. 192. — 824 p.
- Коблик, 2001, с. 56.
- HBW Alive: Family Apodidae, Systematics.
- Scopoli C., Wolfgang G. Introductio ad historiam naturalem sistens genera lapidum, plantarum, et animalium :hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae. — Pragae: Apud Wolfgangum Gerle, 1777. — P. 483. — 540 p.
- Jobling J. A. The Helm Dictionary of Scientific Bird Names. — London: A&C Black Publishers Ltd, 2010. — P. 254. — 432 p. — ISBN 978-1-4081-2501-4.
- Illiger C. Prodromus systematis mammalium et avium. — Berolini : Sumptibus C. Salfeld, 1811. — P. 229. — 301 p.
- Jobling J. A. The Helm Dictionary of Scientific Bird Names. — London: A&C Black Publishers Ltd, 2010. — P. 129. — 432 p. — ISBN 978-1-4081-2501-4.
- Hartert E. Fam. Macropterygidae (англ.) // Podargidae, Caprimulgidae und Macropterygidae. — Berlin: R. Friedländer und sohn, 1897. — P. 62—90.
- HBW Alive: Family Apodidae, Morphological Aspects.
- Соломахина Л.А. Нормальные вариации глазного дна птиц // VetPharma. — 2016. — № 3. — С. 56—65.
- Chantler, Driessens, 2000, p. 19.
- Коблик, 2001, с. 55.
- Collins C. T. A Reinterpretation of Pamprodactyly in Swifts: a Convergent Grasping Mechanism in Vertebrate // Auk. — 1983. — Vol. 100. — P. 735—737.
- Holmgren J. A parsimonious phylogenetic tree for the swifts, Apodi, compared with DNA-analysis phylogenies // Bull. B.O.C.. — 1998. — Vol. 118. — P. 238—249.
- Chantler, Driessens, 2000, p. 38.
- HBW Alive: Family Apodidae, Voice.
- HBW Alive: Family Apodidae, General Habits.
- Chantler, Driessens, 2000, p. 41.
- Swifts (англ.). Encyclopaedia Britannica. Дата обращения: 7 марта 2019.
- Anders Hedenström, Gabriel Norevik, Kajsa Warfvinge, Arne Andersson, Johan Bäckman, Susanne Åkesson. Annual 10-Month Aerial Life Phase in the Common Swift Apus apus.
- Chantler, Driessens, 2000, p. 31.
- HBW Alive: Family Apodidae, Movements.
- Коблик, 2001, с. 58.
- HBW Alive: Family Apodidae, Habitat.
- HBW Alive: Family Apodidae, Food and Feeding.
- Прокофьева И. В. Наблюдения за выкармливанием птенцов чёрного стрижа // Биология питания, развитие и поведение птиц. — Русский орнитологический журнал. — Ленинград, 1976. — С. 127—137.
- Lack D., Owen D.E. The food of the swift // Journal of Animal Ecology. — 1955. — № 24. — P. 239—252.
- Люлеева Д. С. Суточное распределение активности и ночные полёты чёрных стрижей Apus apus // Русский орнитологический журнал. — 2006. — № 15 (306). — С. 7—9.
- Tischmacher J. Etude du regime alimentaire du martinet noir (Apus apus) dans les Basses-Pyrénées // Bull. Centre etudes at reech scient. — 1961. — № 3. — С. 399—401.
- Chantler, Driessens, 2000, p. 33.
- Chantler, Driessens, 2000, p. 34.
- Резанов А. Г. О кормовом поведении стрижей и ласточек во время продолжительных дождей и похолоданий // Русский орнитологический журнал. — 2003. — № 12 (226). — С. 660—663.
- Булюк В. Н., Чернецов Н. С. Сравнительный анализ пространственно-временного распределения совместно обитающих чёрного стрижа, деревенской и городской ласточек при использовании пищевых ресурсов // Русский орнитологический журнал. — 1993. — № 2. — С. 239—252.
- HBW Alive: Family Apodidae, Breeding.
- Chantler, Driessens, 2000, p. 29.
- Chantler, Driessens, 2000, p. 27.
- Прокофьева И. В. Особенности поведения чёрных стрижей Apus apus в период гнездования // Русский орнитологический журнал. — 2006. — Т. 15, № 333. — С. 951—956.
- Люлеева Д.С. Стрижи: Миграции и гнездование пяти видов стрижей (чёрного, белопоясного, малого, белобрюхого и иглохвостого) на территории России и сопредельных стран // Труды Зоологического института РАН. — 1993. — № 254. — С. 1—176.
- Литун В. И. К гнездовой биологии чёрного стрижа // Экология гнездования птиц и методы её изучения. — Самарканд, 1979. — С. 131—132.
- Коблик, 2001, с. 57.
- Chantler, Driessens, 2000, p. 28.
- Коблик, 2001, с. 57—58.
- Chantler, Driessens, 2000, p. 36.
- HBW Alive: Family Apodidae, Relationship with Man.
- Shun Wan Chan. Review of Scientific Research on Edible Bird's Nest (англ.) (2006). Дата обращения: 13 августа 2019.
- HBW Alive: Family Apodidae, Status and Conservation.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Owlet-nightjars, treeswifts, swifts (англ.). IOC World Bird List (v11.2) (15 июля 2021). doi:10.14344/IOC.ML.11.2. Дата обращения: 16 августа 2021.
- Apodidae (англ.). The IUCN Red List of Threatened Species.
- Ksepka D. T., Clarke J. A., Nesbitt S. J., Kulp F. B., Grande L. Fossil evidence of wing shape in a stem relative of swifts and hummingbirds (Aves, Pan-Apodiformes) (англ.) // Proceedings: Biological Sciences. — Royal Society, 2013. — Vol. 280, no. 1761. — P. 1—8. — doi:10.1098/rspb.2013.0580.
- Harrison C. J. O. A revision of the fossil swifts Vertebrata, Aves, Suborder, Apodi),with descriptions of three new genera and two new species (англ.) // Mededelingen van de werkgroep voor tertiaire en kwartaire geologie. — 1984. — Vol. 21, iss. 4. — P. 157—177.
- Mayr G. Phylogeny of early tetriary swifts and hummingbirds (Aves: Apodiformes) // Auk. — 2003. — Vol. 120. — P. 145—151.
- Chantler, Driessens, 2000, p. 20.
- Brooke R. K. Taxonomic and evolutionaty notes on the subfamilies, tribes, genera and subgenera of the swifts (Aves: Apodidae) // Durban Museum Novitates. — 1970. — Vol. IX. — P. 13—24.
- Sclater P. L. Notes on Genera and Species of Cypselidae // Proceedings of the Zoological Society of London. — 1865. — P. 593—617.
- Lack D. A review of the genera and nesting havits of swifts // Auk. — 1956. — Vol. 73. — P. 13—32.
- Apodidae (англ.) информация на сайте Paleobiology Database. (Дата обращения: 17 августа 2019)
- Hackett S. J. et al. A Phylogenomic Study of Birds Reveals Their Evolutionary History (англ.) // Science. — 2008-07. — P. 1763—1768. — doi:10.1126/science.1157704.
- Caprimulgiformes (англ.). Handbook of the Birds of the World Alive. Дата обращения: 18 февраля 2019.
Литература
- Family Apodidae (Swifts) / Chantler P. // Barn-owls to Hummingbirds. — Barcelona : Lynx Edicions, 1999. — (Handbook of the Birds of the World : [16 v.] / In: J. del Hoyo, A. Elliott, J. Sargatal ; 1992—2013, vol. 5). — ISBN 978-84-87334-25-2.
- Коблик Е. А. Семейство настоящие стрижи — Apodidae // Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). — М.: Издательство МГУ, 2001. — Т. 3. — С. 54—62. — 360 с. — 400 экз. — ISBN 5-211-04072-4.
- Chantler P., Driessens G. Swifts: A Guide to the Swifts and Treeswifts of the World. — Second edition. — Sussex: Pica Press, 2000. — 272 с. — ISBN 978-1-8734-0383-9.
Ссылки
- Chantler P. Family Apodidae (Swifts) (англ.). Handbook of the Birds of the World Alive (10 сентября 2014). Дата обращения: 18 апреля 2019.
| Таксономия |
|---|