Вокализация птиц
Вокализация птиц — набор голосовых звуков, издаваемых птицами. В орнитологии разделяется на длинное пение и короткие позывы, различные по функциям — пение используется преимущественно в половом поведении и для защиты территории, позывы предназначены преимущественно для коммуникации, например призыва или сообщения об опасности.

Большинство звуков, издаваемых птицами, мелодично для человеческого уха и оказало влияние на развитие музыки.
Общая характеристика
В вокализации выделяют песни (пение) и позывы[1][2]. Песни длинные, более сложные, видоспецифичные и обычно в некоторой степени выученные; они связаны с брачным и территориальным поведением. Позывы короткие, простые, меньше отличаются у разных видов и в основном врождённые; они выполняют определённые узкие функции (например, предупреждения или удержание стаи вместе)[3][1][2].
Пение наиболее развито у птиц отряда воробьинообразных, особенно подгруппы певчие воробьиные. Большей частью пение свойственно самцам, а не самкам, хотя встречаются исключения. Чаще всего птицы поют, сидя на каком-либо субстрате, хотя некоторые виды способны петь в полёте. Некоторые группы птиц практически молчаливы, издают лишь механические звуки, например аист, лишь щелкают клювами. У некоторых манакинов (Pіprіdae) самцы развили несколько механизмов образования таких звуков, включая стрекотание перьями, подобное стрекотанию насекомых[4].
Образование звуков другими, кроме сиринкса, частями тела (клювом, крыльями, хвостом, перьями и т. д.) и механическими средствами носит название инструментальной музыки (по определению Чарльза Дарвина), инструментальных или механических звуков[5][2] или, в работах некоторых авторов, сонации[6].
Анатомия
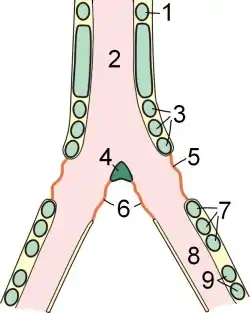
1 — последнее свободное хрящевое кольцо трахеи; 2 — тимпанальный орган; 3 — первая группа колец сиринкса; 4 — козелок; 5 — боковая тимпанальная мембрана; 6 — средняя тимпанальная мембрана; 7 — вторая группа колец сиринкса; 8 — бронх; 9 — хрящевые кольца бронха
Голосовым органом птиц является сиринкс. Это костная структура в месте бифуркации трахеи. В отличие от млекопитающих, у птиц отсутствуют голосовые складки. Звук издаётся вследствие вибраций тимпанальных мембран (стенок сиринкса) и козелка, вызываемых продуванием воздуха через сиринкс[7]. Специальные мышцы способны изменять натяжение мембран и диаметр просвета бронхов, что приводит к изменению издаваемого звука.
Сиринкс и иногда воздушные мешки, которые окружают его, резонируют в ответ на вибрации, которые создаются мембранами, мимо которых проходит воздух при дыхании. Птица контролирует частоту звука изменением натяжения мембран. Так птицы контролирует как частоту, так и громкость, изменяя скорость выдоха. Птицы способны независимо контролировать обе стороны трахеи, поэтому некоторые виды образовывают две главные частоты одновременно.
Социальные аспекты и этология
Функция
Принято считать, что пение птиц развилось прежде всего в результате полового отбора как элемент полового поведения, в частности ухаживания и привлечения самок самцами. Кроме того, важной функцией пения является обозначение территории. По данным экспериментов, качество вокализации является индикатором приспособленности к сложным условиям окружающей среды[8]. Также, по данным экспериментов, на характеристики и частоту пения могут влиять паразиты и заболевания, таким образом, вокализация является прямым индикатором здоровья[9][10]. Репертуар пения также является важным индикатором приспособленности[11][12], способности самцов предрасполагать самок и обозначать территорию. Часто разные по функции типы пения производятся лишь на протяжении определённого сезона или в разные времена года, когда необходимо выполнение определённой функции, и лишь в это время они воспринимаются другими птицами. Например, самец западного соловья (Luscіnіa megarhynchos) издает пение, предназначенное для привлечения самок, лишь ночью (когда поют лишь неспаренные самцы), а пение, предназначенное для обозначения территории, — почти исключительно на протяжении утреннего хора (когда поют все самцы)[13].
Позывы используются прежде всего для коммуникации. Такое общение осуществляется как в пределах одного вида, так и между видами. Общие сигналы часто используются для привлечения индивидуальных птиц к стае. Эти позывы характеризуются широким спектром и резким началом и окончанием, а их повторение, обычное среди многих видов, как считается, полезно для определения расположения стаи. Сигналы предупреждения об опасности, в отличие от них, у большинства видов характеризуются высокой частотой звука, который затрудняет определение положения птицы, выдающей такой сигнал[14].
Часто птицы могут очень хорошо различать позывы, что позволяет им распознавать друг друга по голосу. В частности, многие птицы, гнездящиеся в колониях, таким образом распознают своих птенцов[15].
Много птиц способны подавать сигналы дуэтом. Порой такие дуэты настолько синхронизированы, что звучат как один голосовой сигнал. Такие сигналы называют антифонными[16]. Дуэтные сигналы отмечены во многих семействах птиц, включая фазановых[17], саванных сорокопутов (Malaconotidae)[18], тимелиевых и некоторых совообразных[19] и попугаеобразных[20]. Сухопутные певчие птицы чаще всего производят подобные сигналы в случае вторжения чужаков на их территорию[21], предлагая роль таких сигналов в межвидовой конкуренции.
Некоторые птицы способны очень хорошо имитировать позывы. У некоторых птиц, таких как дронговые, такое имитирование может служить для формирования многовидовых стай[22].
Некоторые пещерные виды, такие как гуахаро и саланганы (рода Collocalia и Aerodramus), используют звуки в диапазоне преимущественно от 2 до 5 кГц для эхолокации в темноте пещер. [23][24][25].
Язык и характеристики вокализации
Язык птиц долго был темой мифов и легенд. Очень давно было известно, что позывы имеют определённое значение, которое соответствующим образом интерпретируется слушателями. Домашние куры, например, издают различные сигналы в ответ на приближение воздушных и наземных хищников, и соответствующим образом отвечают на них[26][27]. Но язык, кроме отдельных слов, должен иметь и определённые грамматические структуры и правила. Исследования подобных структур у птиц довольно трудны из-за большого количества возможных интерпретаций. В одном исследовании, тем не менее, исследователям удалось продемонстрировать способность попугаев к образованию грамматических структур, включая наличие таких понятий, как существительное, глагол и прилагательное[28]. Исследование позывов скворцов также показали наличие рекурсивных структур[29].
Обычно при описании языка птиц охотниками и натуралистами выделяют 5 главных типов звуков: призыв, песня, территориальный сигнал, ухаживание и тревога. Первые четыре представляют «базовое» поведение и подаются при относительной безопасности и покое, тогда как последний означает присутствие хищника или другой угрозы. В пределах каждой категории значения звуков зависят от модуляции голоса, движения тела и контекста[30].
Слух птиц может выходить за частотные пределы слуха человека, опускаясь у некоторых видов как ниже 50 Гц, так и выше 20 кГц, с максимальной чувствительностью между 1 и 5 кГц[31][12].
Диапазон частот голосовых сигналов зависит от условий окружающей среды, прежде всего имеющихся шумов. По обыкновению узкие частотные диапазоны, низкие частоты, низкочастотная модуляция и длинная продолжительность звуков и интервалов между ними характерны для пространств с плотной растительностью (где происходит поглощение и отражение звуков), тогда как высокие частоты, широкие диапазоны, высокочастотная модуляция и короткие элементы сигналов характерные для открытых пространств[32][33]. Была выдвинута гипотеза, что разные птицы и их виды делят между собой время и частотный ресурс, в результате чего у каждого отдельного вида длина и диапазон частот звуковых сигналов уменьшаются (то есть вид занимает свою «акустическую нишу»)[34]. Птицы поют громче и на более высоких частотах в городских районах, где имеется значительный низкочастотный шум[35][36].
Диалекты

Вокализация птиц даже одного вида часто довольно сильно отличается, образовывая «диалекты». Эти диалекты могут возникать как из-за разнообразия окружающей среды, так и вследствие генетического дрейфа, хотя явление мало исследовано, влияние отдельных факторов остается неизвестным даже для хорошо исследованных видов[37]. Лучше всего эти отличия исследованы для пения во время брачного периода. Тем не менее, следствия этого явления не являются одинаковыми и существенным образом отличаются в зависимости от вида птиц.
Самки, которые выросли под влиянием одного диалекта, не отвечают или отвечают хуже на пение самца того же вида, который владеет другим диалектом, что было продемонстрировано, например, для белоголовой зонотрихии (Zonotrichia leucophrys)[38][39]. С другой стороны, самки, которые происходят из районов, где распространены несколько диалектов или диалекты разных подвидов, не проявляют такого предпочтения к одному диалекту[40][41].
Также был исследован и ответ территориальных самцов на пение чужаков. Так, обычно самцы сильнее всего отвечают на пение представителей собственного диалекта, слабее на представителей своего вида из других районов, и ещё слабее на пение родственных видов[42], а самцы, которые разделяют больше песен с соседствующими, лучше охраняют свою территорию[43].
В связи с возникновением диалектов часто рассматривается вопрос их влияния на видообразование. Например, это явление было продемонстрировано в работах по исследованию вьюрков Дарвина[44]. Другие работы, тем не менее, указывают на противоречивость данных по этому вопросу[45][46][37].
Обучение и контроль
Общие характеристики
Пение птиц разных видов довольно сильно отличается и часто является характерным признаком вида. Именно пение часто препятствует смешению родственных видов, генетически достаточно близких для создания жизнеспособного потомства[47]. В современных исследованиях пение характеризуется с помощью акустической спектроскопии. Виды сильно отличаются по сложности пения и по числу типов песен, которое может достигать 3000 у бурого пересмешника; у некоторых видов даже отдельные особи отличаются этой характеристикой. У нескольких видов, таких как скворцы и пересмешники, пение включает случайные элементы, запоминавшиеся на протяжении жизни птицы в форме мимикрии или «присвоение» (из-за того, что птица использует звуки, характерные для других видов). Ещё в 1773 году было установлено, что в экспериментах по выращиванию птенцов птицами других видов коноплянка (Acanthіs cannabіna) была способна выучить пение жаворонка (Alauda arvensis)[48]. У многих видов кажется, что хотя основная песня и одинаковая для всех представителей вида, молодые птицы изучают некоторые детали пения от своих родителей, при этом вариации накапливаются, формируя «диалекты»[49].
Обычно птицы изучают песни на протяжении начала жизни, хотя определённые особенности продолжают накапливаться и позднее, формируя пение взрослой птицы. Зебровая амадина, популярнейший модельный организм для исследования пения птиц, издаёт пение, которое напоминает взрослое, приблизительно после 20 суток после вылупления. К возрасту 35 дней птенец уже полностью изучает взрослое пение. Наиболее ранние песни довольно «пластичны», подвержены изменениям, и птице надо примерно 2—3 месяца для доведения пения до окончательной формы, которую оно имеет у половозрелых особей[50].
Исследования также указывают на то, что обучение пению является формой обучения, в которой принимают участие участки базальных ганглиев. Часто модели обучения птиц используются в качестве моделей изучения языка человеком[51]. У некоторых видов (например, зебровая амадина) обучение ограничено первым годом жизни, эти виды называют «ограниченными по возрасту» или «закрытыми». Другие виды, такие как канарейка, способны изучать новые песни даже в половозрелом возрасте, такие виды называются «открытыми» или «неограниченными по возрасту»[52][53].
Исследователи высказывали предположение, что обучение песням через широкое культурное общение позволяет образование межвидовых диалектов, которые помогают птицам адаптироваться к разнообразному акустическому окружению[54].
Обучение птиц родителями было впервые продемонстрировано в экспериментах Уильяма Торпи 1954 года. Выращенные в изоляции от самцов своего вида птицы способны петь, а их пение за общими чертами по обыкновению напоминает пение взрослых птиц, тем не менее, не имеет сложных элементов и часто существенным образом отличается[55]. Такое пение часто не способно предрасполагать самок. Кроме пения родителей, птенцам также важно слышать своё собственное пение на протяжении сенсоромоторного периода. Птицы, которые потеряли слух к кристаллизации пения, производят пение, которое существенным образом отличается от характерного для данного вида[56].
Присвоение и имитация
Многие птицы способны перенимать пение не только своего вида, но и других, более или менее родственных видов. Так, птенцы многих видов, выращенные родителями родственных видов, часто способны развивать пение, которое напоминает пение приёмных родителей, и в некоторых случаях даже предрасполагать самок этого вида[44]. Другие птицы способны перенимать пение птиц других видов, даже когда выращиваются собственными родителями. Несколько сотен видов по всему миру способны к такой имитации. Например, название пересмешник (Mіmus) было дано этой птице именно за способность копировать звуки других птиц и воссоздавать их. Другим известным видом, способным к копированию, является обыкновенный скворец (Sturnus vulgarіs), в частности в Северной Америке, куда эта птица была завезена из Европы, он «передразнивает» даже пересмешника. В Европе и Британии обыкновенный скворец является известнейшим имитатором пения других птиц, который часто воссоздает звуки таких птиц, как обыкновенный канюк (Buteo buteo), Oriolus oriolus, Numenius arquata, серая неясыть (Strіx aluco), утиные и гуси. В некоторых случаях эти птицы способны имитировать голос грудного ребёнка или даже звуки падающих бомб во время Второй мировой войны. По некоторым данным, один скворец сымитировал свисток футбольного судьи, чем вызвал недоразумение во время матча[57].
Наиболее впечатляющим и популярным среди людей примером имитации звуков птицами является имитация человеческого языка. Известно несколько волнистых попугайчиков, выросших в неволе, чей репертуар достигал 550 слов. Также хорошими способностями для имитации языка человека владеют попугай жако (Psіttacus erіthacus), австралийские попугаи, такие как какаду (Cacatua galerita) и южноамериканские амазоны (Amazona). Александр фон Гумбольдт во время исследования Южной Америки описал случай, когда ему удалось услышать от попугая «мертвый язык» исчезнувшего племени атуров. В Европе случаи способности имитировать голос человека были известны среди некоторых представителей семейства врановых, такие как галка (Corvus monedula), сорока (Pіca pіca) и ворон (Corvus corax)[58].
Тем не менее, точные причины такой имитации неизвестны. Вероятно, они являются усложнениями собственного пения, однако польза такого явления для птицы все ещё остается предметом исследований.
Также существуют случаи присвоения позывов, а не пения птиц. Например, толстоклювая эуфония (Euphonia laniirostris) часто издаёт сигнал угрозы других видов во время приближения потенциального хищника к её гнезду, оставаясь в безопасности. Подобное поведение характерно и для сойки (Garrulus glandarіus) и красноголовой горихвостки (Cossypha natalensis). В других случаях имитация используется для заманивания жертвы в ловушку, так, дымчатый лесной сокол (Micrastur mirandollei) способен имитировать призывы о помощи своих жертв, а потом ловит птиц, которые прилетели в ответ на призыв[59].
Нейрофизиология
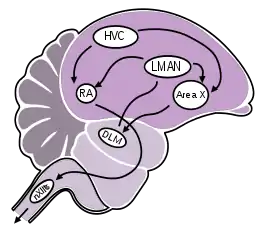
В контроле голосовых сигналов принимают участие следующие отделы головного мозга:
- Путь формирования пения: состоит из верхнего вокального центра (hіgh vocal center или hyperstrіatum ventralіs pars caudalіs, HVC), ядра аркопиллиума (rubust nucleus of arcopіllіum, RA) и части подъязычного ядра, которое идёт к трахее и сиринксу (tracheosyrіngeal nerve)[60][61];
- Передняя часть переднего мозга, который отвечает за обучение: состоит из латеральной части магноцеллюлярного ядра переднего нового полосатого тела (lateral part of the magnocellular nucleus of anterіor neostrіatum, LMAN, гомолога базальных ганглиев млекопитающих), области X (части базальных ганглиев) и дорсально-латерального отдела среднего таламуса (DLM).
Пение птиц и музыка
Примечания
- И. Р. Бёме. Формирование вокализации воробьиных птиц (Passeriformes) в онтогенезе. Современное состояние проблемы // Журнал общей биологии. — 2006. — Т. 67, № 4. — С. 268—279. — PMID 17022487. синопсис
- Ильичёв В. Д., Карташёв Н. Н., Шилов И. А. Общая орнитология. — М.: Высшая школа, 1982. — С. 225, 238. — 464 с.
- Ehrlich, Paul R., David S. Dobkin, and Darryl Wheye. "Vocal Functions", "Bird Voices" and "Vocal Development" from Birds of Stanford essays. Дата обращения: 9 сентября 2008. Архивировано 25 июля 2018 года.
- Bostwick, Kimberly S. and Richard O. Prum. Courting Bird Sings with Stridulating Wing Feathers (англ.) // Science : journal. — 2005. — Vol. 309, no. 5735. — P. 736. — doi:10.1126/science.1111701. — PMID 16051789.
- Manson-Barr P., Pye J. D. Mechanical sounds // A Dictionary of Birds / B. Campbell, E. Lack. — Staffordshire: Poyser, 1985. — P. 342—344. — ISBN 978-1-4081-3840-3.
- Bostwick, Kimberly S. and Richard O. Prum. High-speed video analysis of wing-snapping in two manakin clades (Pipridae: Aves) (англ.) // The Journal of Experimental Biology. — The Company of Biologists, 2003. — Vol. 206. — P. 3693—3706. — doi:10.1242/jeb.00598. — PMID 12966061.
- Direct observation of syringeal muscle function in songbirds and a parrot Ole Næsbye Larsen and Franz Goller, The Journal of Experimental Biology 205, 25-35 (2002)
- Read, A. W. and D. M. Weary. Sexual selection and the evolution of bird song: A test of the Hamilton-Zuk hypothesis (англ.) // Behavioral Ecology and Sociobiology. — 1990. — Vol. 26, no. 1. — P. 47—56. — doi:10.1007/BF00174024.
- Garamszegi, L. Z., A. P. Møller, János Török, Gábor Michl, Péter Péczely and Murielle Richard. Immune challenge mediates vocal communication in a passerine bird: an experiment (англ.) // Behavioral Ecology. — 2004. — Vol. 15, no. 1. — P. 148—157. — doi:10.1093/beheco/arg108.
- Redpath, S. M., Bridget M Appleby, Steve J Petty. Do male hoots betray parasite loads in Tawny Owls? (англ.) // Journal of Avian Biology : journal. — 2000. — Vol. 31, no. 4. — P. 457—462. — doi:10.1034/j.1600-048X.2000.310404.x.
- Reid, J. M., Peter Arcese, Alice L. E. V. Cassidy, Sara M. Hiebert, James N. M. Smith, Philip K. Stoddard, Amy B. Marr, and Lukas F. Keller. Fitness Correlates of Song Repertoire Size in Free-Living Song Sparrows (Melospiza melodia) (англ.) // The American Naturalist : journal. — University of Chicago Press, 2005. — Vol. 165. — P. 299—310. — doi:10.1086/428299.
- A. P. Møller, J. Erritzøe, L. Z. Garamszegi. Covariation between brain size and immunity in birds: implications for brain size evolution (англ.) // Journal of Evolutionary Biology. — 2005. — Vol. 18, no. 1. — P. 223—237. — doi:10.1111/j.1420-9101.2004.00805.x. — PMID 15669979.
- Roth T., Sprau P., Schmidt R., Naguib M., Amrhein V. Sex-specific timing of mate searching and territory prospecting in the nightingale: nocturnal life of females (англ.) // Proc Biol Sci. : journal. — 2009. — PMID 19324798.
- Marler, P. Characteristics of some animal calls (англ.) // Nature. — 1955. — Vol. 176. — P. 6—8. — doi:10.1038/176006a0.
- Lengagne, T., J. Lauga and T. Aubin. Intra-syllabic acoustic signatures used by the King Penguin in parent-chick recognition: an experimental approach (англ.) // The Journal of Experimental Biology. — The Company of Biologists, 2001. — Vol. 204. — P. 663—672.
- Thorpe, W. H. Antiphonal Singing in Birds as Evidence for Avian Auditory Reaction Time (англ.) // Nature : journal. — Antiphonal singing in birds as evidence for avian auditory reaction time. — Vol. 197. — P. 774—776. — doi:10.1038/197774a0.
- Stokes, A., W. and H. W. Williams. Antiphonal calling in quail (англ.) // American Ornithological Society. — 1968. — Vol. 85. — P. 83—89.
- Harris, Tony; Franklin, Kim. Shrikes and Bush-Shrikes (англ.). — Princeton University Press, 2000. — P. 257—260. — ISBN 0-691-07036-9.
- Osmaston, B. B. "Duetting" in birds (англ.) // Ibis. — Wiley-Blackwell, 1941. — Vol. 5. — P. 310—311. — doi:10.1111/j.1474-919X.1941.tb00620.x.
- Power, D. M. Antiphonal duetting and evidence for auditory reaction time in the Orange-chinned Parakeet (англ.) // American Ornithological Society : journal. — 1966. — Vol. 83. — P. 314—319.
- Hyman, Jeremy. Countersinging as a signal of aggression in a territorial songbird (англ.) // Animal Behaviour : journal. — Elsevier, 2003. — Vol. 65. — P. 1179—1185. — doi:10.1006/anbe.2003.2175.
- Goodale, E. and Kotagama, S. W. Testing the roles of species in mixed-species bird flocks of a Sri Lankan rain forest (англ.) // Journal of Tropical Ecology : journal. — 2005. — Vol. 21. — P. 669—676. — doi:10.1017/S0266467405002609.
- Suthers RA and Hector D. H. The physiology of vocalization by the echolocating Oilbird, Steatornis caripensis (англ.) // J. Comp. Physiol. : journal. — 1985. — Vol. 156, no. 2. — P. 243—266. — doi:10.1007/BF00610867.
- Suthers RA and Hector D. H. Mechanism for the production of echolocating clicks by the Grey Swiftlet, Collocalia spodiopygia (англ.) // J. Comp. Physiol. A : journal. — 1982. — Vol. 148. — P. 457—470. — doi:10.1007/BF00619784.
- Coles R. B., Konishi M and Pettigrew J. D. Hearing and echolocation in the Australian Grey Swiftlet, Collocalia spodiopygia (англ.) // The Journal of Experimental Biology : journal. — The Company of Biologists, 1987. — Vol. 129. — P. 365—371.
- Collias, N. E. The vocal repertoire of the Red Junglefowl: A spectrographic classification and the code of communication (англ.) // American Ornithological Society : journal. — 1987. — Vol. 89. — P. 510—524. — doi:10.2307/1368641.
- Evans, C. S., Macedonia, J. M., and Marler, P. Effects of apparent size and speed on the response of chickens, Gallus gallus, to computer-generated simulations of aerial predators (англ.) // Animal Behaviour : journal. — Elsevier, 1993. — Vol. 46. — P. 1—11. — doi:10.1006/anbe.1993.1156.
- Pepperberg, I.M. The Alex Studies: Cognitive and Communicative Abilities of Grey Parrots (англ.). — Harvard University Press, 2000.
- Marcus, Gary F. Language: Startling starlings (англ.) // Nature. — 2006. — 27 April (vol. 440, no. 7088). — P. 1117—1118. — ISSN 0028-0836. — doi:10.1038/4401117a.
- Young, Jon, and Gardoqui, Dan. Bird Language Intensive Workshop Presentation. Regenerative Design Institute (2008). Архивировано 7 апреля 2012 года.
- Dooling, R.J. Auditory perception in birds. Acoustic Communication in Birds, Vol. 1 (eds D.E. Kroodsma & E.H. Miller) (англ.). — 1982. — P. 95—130.
- Boncoraglio, G. and Nicola Saino. Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis (англ.) // Functional Ecology : journal. — 2007. — Vol. 21. — P. 134—142. — doi:10.1111/j.1365-2435.2006.01207.x.
- Morton, E.S. Ecological sources of selection on avian sounds (англ.) // The American Naturalist : journal. — University of Chicago Press, 1975. — Vol. 109. — P. 17—34. — doi:10.1086/282971.
- Krause, Bernard L. The Niche Hypothesis (англ.) // The Soundscape Newsletter. — 1993. — Vol. 06.
- Henrik Brumm. The impact of environmental noise on song amplitude in a territorial bird (англ.) // Journal of Animal Ecology. — 2004. — Vol. 73, no. 3. — P. 434—440. — doi:10.1111/j.0021-8790.2004.00814.x.
- Slabbekoorn, H. and Peet, M. Birds sing at a higher pitch in urban noise (англ.) // Nature. — 2003. — Vol. 424. — P. 267. — doi:10.1038/424267a.
- Slabbekoorn H., Smith T. B. Bird song, ecology and speciation (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences. — 2002. — Vol. 357, no. 1420. — P. 493—503. — PMID 12028787.
- Baker, M. C. Vocal dialect recognition and population genetic consequences (англ.) // Integrative and Comparative Biology : journal. — Oxford University Press. — Vol. 22. — P. 561—569.
- Baker, M. C. The behavioural responses of female Nuttall’s white-crowned sparrows to male song of natal and alien dialects (англ.) // Behavioral Ecology and Sociobiology. — 1983. — Vol. 12. — P. 309—315. — doi:10.1007/BF00302898.
- Chilton, G. & Lein, M. R. Song and sexual responses of female white-crowned sparrows Zonotrichia leucophrys from a mixed-dialect population (англ.) // Behaviour : journal. — 1996. — Vol. 133. — P. 173—198. — doi:10.1163/156853996X00107.
- Chilton, G., Lein, M. R. & Baptista, L. F. Mate choice by female white-crowned sparrows in a mixed-dialect population (англ.) // Behavioral Ecology and Sociobiology. — 1990. — Vol. 27. — P. 223—227. — doi:10.1007/BF00180307.
- Milligan, M. M. & Verner, J. Inter-population song dialect discrimination in the white-crowned sparrow (англ.) // Condor : journal. — 1971. — Vol. 73. — P. 208—213.
- Beecher, M. D., Campbell, S. E. & Nordby, J. C. Territory tenure in song sparrows is related to song sharing with neighbours, but not to repertoire size (англ.) // Animal Behaviour. — Elsevier, 2000. — Vol. 59. — P. 29—37. — doi:10.1006/anbe.1999.1304. — PMID 10640364.
- Ratcliffe, L. M. & Grant, P. R. Species recognition in Darwin’s finches (Geospiza, Gould). III. Male responses to playback of different song types, dialects and heterospecific songs (англ.) // Animal Behaviour : journal. — Elsevier, 1985. — Vol. 33. — P. 290—307. — doi:10.1016/S0003-3472(85)80143-3).
- Zink, R. M. Genetic population structure and song dialects in birds (англ.) // Behav. Brain Sci. : journal. — 1985. — Vol. 8. — P. 118—119.
- Petrinovich, L. An unbalanced survey of bird-song research: smoke gets in your eyes (англ.) // Behav. Brain Sci. : journal. — 1985. — Vol. 8. — P. 113—114.
- Jeffrey Podos. Correlated evolution of morphology and vocal signal structure in Darwin's finches (англ.) // Nature : journal. — 2001. — Vol. 409. — P. 185—188. — doi:10.1038/35051570.
- Barrington, D. Experiments and observations on the singing of birds (англ.) // Philosophical Transactions of the Royal Society of London : journal. — 1773. — Vol. 63. — P. 249—291. — doi:10.1098/rstl.1773.0031.
- Marler, P., & M. Tamura. Song dialects in three populations of the white-crowned sparrow (англ.) // Condor : journal. — 1962. — Vol. 64. — P. 368—377. — doi:10.2307/1365545.
- Nottebohm, F. The Neural Basis of Birdsong (англ.) // PLoS Biol : journal. — 2005. — Vol. 3, no. 5. — P. 163. — doi:10.1371/journal.pbio.0030164.
- Teramitsu I., Kudo L. C., London S. E., Geschwind D. H., White S. A. Parallel FoxP1 and FoxP2 expression in songbird and human brain predicts functional interaction (англ.) // J. Neurosci.. — 2004. — Vol. 24, no. 13. — P. 3152—3163. — doi:10.1523/JNEUROSCI.5589-03.2004. — PMID 15056695.
- Nottebohm, F. The road we travelled: discovery, choreography, and significance of brain replaceable neurons (англ.) // Ann. N. Y. Acad. Sci. : journal. — 2004. — Vol. 1016. — P. 628—658. — doi:10.1196/annals.1298.027.
- Brenowitz, Eliot A. and Michael D. Beecher. Song learning in birds: diversity and plasticity, opportunities and challenges (англ.) // Trends in Neurosciences : journal. — Cell Press, 2005. — Vol. 28, no. 3. — P. 127—132. — doi:10.1016/j.tins.2005.01.004.
- Slater, P. J. B. Bird song learning: causes and consequences (англ.) // Ethol. Ecol. Evol. : journal. — 1989. — Vol. 1. — P. 19—46.
- Thorpe, W. The process of song-learning in the chaffinch as studied by means of the sound spectograph (англ.) // Nature : journal. — 1954. — Vol. 173. — P. 465—469. — doi:10.1038/173465a0.
- Konishi, M. The role of auditory feedback on the control of vocalization in the white-crowned sparrow (англ.) // Zeitschrift fur Tierpsychologie : journal. — 1965. — Vol. 22. — P. 770—783.
- The language of birds: Vocal appropriation. Listen to Nature. British Library. Архивировано 20 января 2012 года.
- The language of birds: "Talking" birds. Listen to Nature. British Library. Архивировано 17 июля 2012 года.
- The language of birds: Calls as deceitful mimicry. Listen to Nature. British Library. Архивировано 20 июня 2012 года.
- Brainard, M. S. and Doupe, A. J. Auditory feedback in learning and maintenance of vocal behavior (англ.) // Nature Rev. Neurosci. : journal. — 2000. — Vol. 1. — P. 31—40. — doi:10.1038/35036205.
- Carew, Thomas J. Behavioral Neurobiology: The Cellular Organization of Natural Behavior (англ.). — Sinauer Associates, Inc., 2000.
| В библиографических каталогах |
|---|