Дрейф генов
Дрейф ге́нов, или гене́тико-автомати́ческие проце́ссы, или эффект Райта (англ. Sewall Wright effect) — случайные изменения частот аллелей и генотипов, происходящие в небольшой полиморфной популяции при смене поколений. Такие случайные изменения приводят к двум характерным последствиям: сначала к флуктуациям частоты аллеля из поколения в поколение, а в конце концов к полному закреплению или элиминации данного аллеля. Влияние дрейфа генов на изменение частот аллелей в данной популяции зависит прежде всего от её размеров[1][2][3][4][5].
Первые работы по изучению случайных процессов в популяциях были проведены в начале 1930-х годов Сьюэлом Райтом в США, Роналдом Фишером в Англии, а также В. В. Лисовским, М. А. Кузнецовым, Н. П. Дубининым и Д. Д. Ромашовым в СССР. Понятие «дрейф генов» (англ. genetic drift) было введено в оборот Райтом (1931), а синонимичное понятие «генетико-автоматические процессы в популяциях» — Дубининым и Ромашовым (1932). Впоследствии в мировой литературе, в том числе и в русскоязычной, закрепился термин С. Райта[6]. В 1942 году Дж. Хаксли в своей книге «Эволюция: современный синтез» предложил называть случайный дрейф генов «эффектом Сьюэла Райта»[1].
В начале 20-го века велись активные дискуссии об относительной важности естественного отбора по сравнению с нейтральными процессами, включая генетический дрейф. Рональд Фишер, объяснивший естественный отбор с использованием менделевской генетики[7], придерживался мнения, что генетический дрейф играет в эволюции самую незначительную роль, и это оставалось доминирующим взглядом в течение нескольких десятилетий. В 1968 году популяционный генетик Мотоо Кимура возродил дебаты своей нейтральной теорией молекулярной эволюции, которая утверждает, что большинство случаев, когда генетические изменения распространяются по популяции (хотя не обязательно изменения в фенотипе), вызваны генетическим дрейфом, действующим на нейтральные мутации[8][9].
Пояснение
Один из механизмов дрейфа генов заключается в следующем. В процессе размножения в популяции образуется большое число половых клеток — гамет. Большая часть этих гамет не формирует зигот. Тогда новое поколение в популяции формируется из выборки гамет, которым удалось образовать зиготы. При этом возможно смещение частот аллелей относительно предыдущего поколения.
Дрейф генов на примере
Механизм дрейфа генов может быть продемонстрирован на небольшом примере. Представим очень большую колонию бактерий, находящуюся изолированно в капле раствора. Бактерии генетически идентичны за исключением одного гена с двумя аллелями A и B. Аллель A присутствует у одной половины бактерий, аллель B — у другой. Поэтому частота аллелей A и B равна 1/2. A и B — нейтральные аллели, они не влияют на выживаемость или размножение бактерий. Таким образом, все бактерии в колонии имеют одинаковые шансы на выживание и размножение.
Затем размер капли уменьшаем таким образом, чтобы питания хватало лишь для 4 бактерий. Все остальные умирают без размножения. Среди четырёх выживших возможно 16 комбинаций для аллелей A и B:
(A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A),
(A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A),
(A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B),
(A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
Вероятность каждой из комбинаций

1/2 (вероятность аллеля A или B для каждой выжившей бактерии) возводится в 4 степень (общий размер результирующей популяции выживших бактерий).
Если сгруппировать варианты по числу аллелей, то получится следующая таблица:
| A | B | Количество вариантов | Вероятность |
| 4 | 0 | 1 | 1/16 |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 0 | 4 | 1 | 1/16 |
Как видно из таблицы, в шести вариантах из 16 в колонии будет одинаковое количество аллелей A и B. Вероятность такого события 6/16. Вероятность всех прочих вариантов, где количество аллелей A и B неодинаково несколько выше и составляет 10/16.
Дрейф генов происходит при изменении частот аллелей в популяции из-за случайных событий. В данном примере популяция бактерий сократилась до 4 выживших (эффект бутылочного горлышка). Сначала колония имела одинаковые частоты аллелей A и B, но шансы, что частоты изменятся (колония подвергнется дрейфу генов) выше, чем шансы на сохранение оригинальной частоты аллелей. Также существует высокая вероятность (2/16), что в результате дрейфа генов один аллель будет утрачен полностью.
Математическая модель
Математические модели генетического дрейфа могут быть разработаны с использованием либо ветвящихся процессов, либо диффузионного уравнения, описывающего изменения частоты аллелей в идеализированной популяции[10].
Модель Райта — Фишера
Рассмотрим ген с двумя аллелями, A или B. В диплоидных популяциях, состоящих из N особей, имеется 2N копий каждого гена. Особь может иметь две копии одного и того же аллеля или двух разных аллелей. Мы можем назвать частоту одного аллеля p и частоту другого q. Модель Райта–Фишера (названная в честь Сьюолла Райта и Рональда Фишера) предполагает, что поколения не совпадают (например, однолетние растения имеют ровно одно поколение в год) и что каждая копия гена, обнаруженная в новом поколении, образуется независимо и случайным образом из всех копий гена в старом поколении. Формула для расчета вероятности получения «k» копий аллеля с частотой «p» в последнем поколении[11][12]

где символ «!» означает факториальную функцию. Это выражение также может быть сформулировано с использованием биномиального коэффициента,

Модель Морана
Модель Морана предполагает перекрывающиеся поколения. На каждом временном шаге выбирается один человек для воспроизведения, а один человек выбирается для смерти. Таким образом, на каждом временном шаге количество копий данного аллеля может увеличиваться на единицу, уменьшаться на единицу или может оставаться неизменным. Это означает, что стохастическая матрица является трехдиагональной, что означает, что математические решения легче для модели Морана, чем для модели Райта — Фишера. С другой стороны, компьютерное моделирование, как правило, легче выполнить с использованием модели Райта-Фишера, поскольку необходимо рассчитать меньшее количество временных шагов. В модели Морана для прохождения одного поколения требуется N временных шагов, где N — эффективный размер популяции. В модели Райта-Фишера требуется всего один[13].
На практике модели Морана и Райта-Фишера дают качественно сходные результаты, но генетический дрейф в модели Морана в два раза быстрее.
Дрейф и фиксация
Закон Харди — Вайнберга гласит, что в достаточно больших популяциях частоты аллелей остаются постоянными от поколения к поколению, если только равновесие не нарушено миграцией, генетическими мутациями или отбором[14].
Однако в конечных популяциях новые аллели не получены в результате случайной выборки аллелей, переданной следующему поколению, но выборка может привести к исчезновению существующего аллеля. Поскольку случайная выборка может удалить, но не заменить аллель, и поскольку случайное снижение или увеличение частоты аллелей влияет на ожидаемое распределение аллелей для следующего поколения, генетический дрейф ведет популяцию к генетической однородности с течением времени. Когда аллель достигает частоты 1 (100 %), она считается «фиксированной» в популяции, а когда аллель достигает частоты 0 (0 %), она исчезает. Меньшие популяции достигают фиксации быстрее, тогда как в пределе бесконечной популяции фиксация не достигается. Как только аллель становится фиксированной, генетический дрейф останавливается, и частота аллеля не может измениться, если новый аллель не введен в популяцию посредством мутации или потока генов. Таким образом, даже несмотря на то, что генетический дрейф является случайным, бесцельным процессом, он устраняет генетические изменения во времени[15].
Скорость изменения частоты аллелей из-за дрейфа
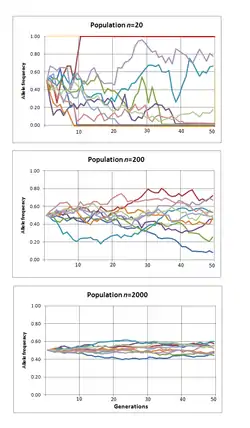
Предполагая, что генетический дрейф является единственной эволюционной силой, действующей на аллель, после t поколений во многих реплицируемых популяциях, начиная с частот аллелей p и q, дисперсия частоты аллелей между этими популяциями

Экспериментальное доказательство С. Райта
С. Райт экспериментально доказал, что в маленьких популяциях частота мутантного аллеля меняется быстро и случайным образом. Его опыт был прост: в пробирки с кормом он посадил по две самки и по два самца мух дрозофил, гетерозиготных по гену А (их генотип можно записать Аа). В этих искусственно созданных популяциях концентрация нормального (А) и мутационного (а) аллелей составила 50 %. Через несколько поколений оказалось, что в некоторых популяциях все особи стали гомозиготными по мутантному аллелю (а), в других популяциях он был вовсе утрачен, и, наконец, часть популяций содержала как нормальный, так и мутантный аллель. Важно подчеркнуть, что, несмотря на снижение жизнеспособности мутантных особей и, следовательно, вопреки естественному отбору, в некоторых популяциях мутантный аллель полностью вытеснил нормальный. Это и есть результат случайного процесса — дрейфа генов.
Против естественного отбора
В природных популяциях генетический дрейф и естественный отбор не действуют изолированно, оба явления всегда играют роль, вместе с мутацией и миграцией. Нейтральная эволюция является продуктом как мутации, так и дрейфа, а не только дрейфа. Точно так же, даже когда отбор преодолевает генетический дрейф, он может действовать только на не нейтральные вариации.
В то время как естественный отбор имеет направление в сторону наследственных адаптаций к текущей среде, генетический дрейф не имеет направления и зависит только от математической случайности[17]. В результате дрейф воздействует на частоты генотипов (числа особей с одинаковым набором ДНК) в популяции, независимо от их фенотипических признаков. В отличие от этого, отбор способствует распространению аллелей, фенотипические эффекты которых увеличивают выживаемость и / или плодовитость их носителей, снижает частоту аллелей, вызывающих неблагоприятные признаки, и игнорирует нейтральные[18].
Закон больших чисел предсказывает, что, когда абсолютное число копий аллеля мало (например, в маленьких популяциях), величина дрейфа на частотах аллеля на поколение больше. Величина дрейфа достаточно велика, чтобы подавить отбор при любой частоте аллеля, когда коэффициент отбора меньше 1, деленного на эффективный размер популяции. Поэтому считается, что не адаптивная эволюция, возникающая в результате мутации и генетического дрейфа, является последующим механизмом эволюционных изменений, главным образом в небольших изолированных популяциях[19]. Математика генетического дрейфа зависит от эффективного размера популяции, но неясно, как это связано с фактическим числом особей в популяции[20]. Генетическая связь с другими генами, которые находятся в процессе отбора, может уменьшить эффективный размер популяции, испытываемый нейтральным аллелем. При более высокой частоте рекомбинации уменьшается связь, и вместе с этим это локальное влияние на эффективный размер популяции[21][22]. Этот эффект виден в молекулярных данных как корреляция между локальной скоростью рекомбинации и генетическим разнообразием[23] и отрицательной корреляцией между плотностью и разнообразием генов в некодирующих областях ДНК[24]. Стохастичность, связанная со связью с другими генами, находящимися в процессе отбора, не является той же ошибкой, что и выборка, и иногда её называют генетической тягой, чтобы отличить её от генетического автостопа[20].
Когда частота аллелей очень мала, дрейф также может усиливать отбор даже в больших популяциях. Например, в то время как неблагоприятные мутации обычно быстро устраняются в больших популяциях, новые полезные мутации почти так же уязвимы для потери из-за генетического дрейфа, как и нейтральные мутации. Только когда частота аллеля для преимущественной мутации достигнет определённого порога, генетический дрейф не будет иметь никакого эффекта[18].
Эффект бутылочного горлышка
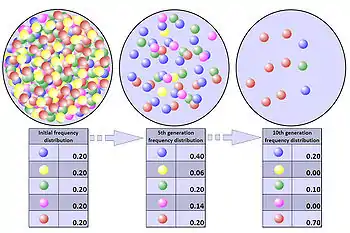
Эффект бутылочного горлышка или «эффект основателя»[25] — сокращение генофонда (то есть генетического разнообразия) популяции вследствие прохождения периода, во время которого по различным причинам происходит критическое уменьшение её численности, в дальнейшем восстановленное[25]. В истинном бутылочном горлышке в популяции шансы на выживание любого организма являются чисто случайными и не улучшаются никаким конкретным врожденным генетическим преимуществом. Эффект может привести к радикальным изменениям частоты аллелей, совершенно не зависящим от отбора[26].
Генетическая вариативность популяции может быть значительно уменьшена эффектом бутылочного горлышка, и даже полезные приспособления могут быть навсегда устранены[27].
См. также
Примечания
- Ермолаев А. И. Роль Сьюэла Райта в создании популяционной генетики // Историко-биологические исследования. — 2012. — Т. 4, № 4. — С. 61—95.
- Genetic drift (англ.) // Current Biology : journal. — Cell Press, 2011. — October (vol. 21, no. 20). — P. R837—8. — doi:10.1016/j.cub.2011.08.007. — PMID 22032182.
- Stephen Jay Gould. Chapter 7, section "Synthesis as Hardening" // The Structure of Evolutionary Theory (неопр.). — 2002.
- Futuyma, 1998, Glossary
- Star B., Spencer H. G. Effects of genetic drift and gene flow on the selective maintenance of genetic variation (англ.) // Genetics : journal. — 2013. — May (vol. 194, no. 1). — P. 235—244. — doi:10.1534/genetics.113.149781. — PMID 23457235.
- Кайданов Л. З. Популяционно-генетические исследования факторов эволюции. В кн.: Развитие эволюционной теории в СССР: (1917—1970-е годы). Под ред. С. Р. Микулинского и Ю. И. Полянского. Л.: «Наука», 1983. С. 143
- Miller, 2000, p. 54
- Evolutionary rate at the molecular level (англ.) // Nature. — Nature Publishing Group, 1968. — February (vol. 217, no. 5129). — P. 624—626. — doi:10.1038/217624a0. — . — PMID 5637732.
- Futuyma, 1998, p. 320
- Wahl L. M. Fixation when N and s vary: classic approaches give elegant new results (англ.) // Genetics : journal. — Genetics Society of America, 2011. — August (vol. 188, no. 4). — P. 783—785. — doi:10.1534/genetics.111.131748. — PMID 21828279.
- Hartl, 2007, p. 112.
- Tian, 2008, p. 11.
- Moran, P. A. P. Random processes in genetics (англ.) // Mathematical Proceedings of the Cambridge Philosophical Society : journal. — 1958. — Vol. 54, no. 1. — P. 60—71. — doi:10.1017/S0305004100033193. — .
- Ewens, 2004
- Li & Graur, 1991, p. 29.
- Barton, 2007, p. 417.
- Natural Selection: How Evolution Works. Actionbioscience. Washington, D.C.: American Institute of Biological Sciences. Дата обращения: 24 ноября 2009. Архивировано 6 января 2010 года. Смотрите ответ на вопрос: Является ли естественный отбор единственным механизмом эволюции?
- Cavalli-Sforza, 1996.
- Zimmer, 2001
- Is the population size of a species relevant to its evolution? (англ.) // Evolution; International Journal of Organic Evolution : journal. — John Wiley & Sons for the Society for the Study of Evolution, 2001. — November (vol. 55, no. 11). — P. 2161—2169. — doi:10.1111/j.0014-3820.2001.tb00732.x. — PMID 11794777.
- Non-Neutral Evolution: Theories and Molecular Data (англ.) / Golding, Brian. — New York: Chapman & Hall, 1994. — ISBN 0-412-05391-8. «Материалы семинара, спонсируемого Канадским институтом перспективных исследований»
- Charlesworth B., Morgan M. T., Charlesworth D. The effect of deleterious mutations on neutral molecular variation (англ.) // Genetics : journal. — Genetics Society of America, 1993. — August (vol. 134, no. 4). — P. 1289—1303. — PMID 8375663.
- Recombination enhances protein adaptation in Drosophila melanogaster (англ.) // Current Biology : journal. — Cell Press, 2005. — September (vol. 15, no. 18). — P. 1651—1656. — doi:10.1016/j.cub.2005.07.065. — PMID 16169487.
- Nordborg M., Hu T. T., Ishino Y., Jhaveri J., Toomajian C., Zheng H., Bakker E., Calabrese P., Gladstone J., Goyal R., Jakobsson M., Kim S., Morozov Y., Padhukasahasram B., Plagnol V., Rosenberg N. A., Shah C., Wall J. D., Wang J., Zhao K., Kalbfleisch T., Schulz V., Kreitman M., Bergelson J. The pattern of polymorphism in Arabidopsis thaliana (англ.) // PLoS Biology : journal. — PLOS, 2005. — July (vol. 3, no. 7). — P. e196. — doi:10.1371/journal.pbio.0030196. — PMID 15907155.

- Большая российская энциклопедия.
- Robinson, Richard, ed. (2003), Population Bottleneck, Genetics, vol. 3, New York: Gale (publisher), ISBN 0-02-865609-1, OCLC 614996575, <http://www.bookrags.com/research/population-bottleneck-gen-03/>. Проверено 14 декабря 2015.
- Futuyma, 1998, pp. 303–304
Литература
- Михеев В. С. ДРЕЙФ ГЕНОВ. Большая российская энциклопедия (2007). Дата обращения: 2 декабря 2019.
- Воронцов Н.Н., Сухорукова Л.Н. Эволюция органического мира. — М.: Наука, 1996. — С. 93—96. — ISBN 5-02-006043-7.
- Грин Н., Стаут У., Тейлор Д. Биология. В 3 томах. Том 2. — М.: Мир, 1996. — С. 287—288. — ISBN 5-03-001602-3.
- Tian, Jianjun Paul. Evolution Algebras and their Applications (англ.). — Berlin; New York: Springer, 2008. — Vol. 1921. — (Lecture Notes in Mathematics). — ISBN 978-3-540-74283-8. — doi:10.1007/978-3-540-74284-5.
- Hartl, Daniel L.; Clark, Andrew G. Principles of Population Genetics (неопр.). — 4th. — Sunderland, MA: Sinauer Associates, 2007. — ISBN 978-0-87893-308-2.
- Futuyma, Douglas Evolutionary Biology (неопр.). — 3rd. — Sunderland, MA: Sinauer Associates, 1998. — ISBN 0-87893-189-9.
- Miller, Geoffrey The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature (англ.). — New York: Doubleday (publisher), 2000. — ISBN 0-385-49516-1.
- Ewens, Warren J. Mathematical Population Genetics I. Theoretical Introduction (англ.). — 2nd. — New York: Springer Science+Business Media, 2004. — Vol. 27. — (Interdisciplinary Applied Mathematics). — ISBN 0-387-20191-2.
- Li, Wen-Hsiung; Graur, Dan Fundamentals of Molecular Evolution (неопр.). — Sunderland, MA: Sinauer Associates, 1991. — ISBN 0-87893-452-9.
- Barton, Nicholas H.; Briggs, Derek E.G.; Eisen, Jonathan A.; Goldstein, David B.; Patel, Nipam H. Evolution (неопр.). — Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 2007. — ISBN 978-0-87969-684-9.
- Cavalli-Sforza, L. Luca; Menozzi, Paolo; Piazza, Alberto The History and Geography of Human Genes (неопр.). — Abridged paperback. — Princeton, N.J.: Princeton University Press, 1996. — ISBN 0-691-02905-9.
- Zimmer, Carl Evolution: The Triumph of an Idea (неопр.). — 1st. — New York: HarperCollins, 2001. — ISBN 0-06-019906-7.
- Non-Neutral Evolution: Theories and Molecular Data (англ.) / Golding, Brian. — New York: Chapman & Hall, 1994. — ISBN 0-412-05391-8. «Материалы семинара, спонсируемого Канадским институтом перспективных исследований».
Ссылки
| Эволюционные процессы | |
|---|---|
| Факторы эволюции | |
| Генетика популяций | |
| Происхождение жизни | |
| Исторические концепции | |
| Современные теории | |
| Эволюция таксонов | |