Эволюционная биология
Эволюцио́нная биоло́гия — раздел биологии, изучающий происхождение видов от общих предков, наследственность и изменчивость их признаков, размножение и разнообразие форм в ходе эволюционного развития. Развитие отдельных видов обычно рассматривается в контексте глобальных преобразований флор и фаун, как компонентов биосферы. Эволюционная биология начала оформляться в качестве раздела биологии с широким признанием идей об изменчивости видов во второй половине XIX века.
.jpg.webp)
Эволюционная биология — междисциплинарная область исследований, поскольку она включает в себя как полевые, так и лабораторные направления различных наук. Вклад в эволюционную биологию вносят исследования в таких узкоспециальных областях, как териология, орнитология или герпетология, которые обобщаются для получения ясной картины развития всего органического мира. Палеонтологи и геологи анализируют окаменелости, чтобы получить сведения о темпах и формах эволюции, а популяционная генетика исследует эти же вопросы теоретически. Экспериментаторы используют селекцию дрозофил для лучшего понимания многих проблем эволюционной биологии, например эволюции старения. В 1990-х годах биология развития вернулась в эволюционную биологию после длительного забвения в виде новой синтетической дисциплины — эволюционной биологии развития.
История
Эволюционная биология как академическая дисциплина стала общепризнанной в результате синтеза дарвиновской теории и генетики в 1930-х и 1940-х годах. Основу новой теории заложили работы Четверикова[1], Фишера[2], Райта[3] и Холдейна[4], в которых рассматривалось воздействие естественного отбора на частоту аллелей в популяциях. Характер этих работ был скорее теоретическим, чем экспериментальнообоснованным[5]. Ситуацию исправила монография Феодосия Добжанского «Генетика и происхождение видов»[6]. В основу проблемы автор положил экспериментальную популяционную генетику. Теоретическая работа предшествующих авторов сопоставлялась с данными по изменчивости и отбору, полученными в ходе различных экспериментов. Добжанский полагал, что макроэволюционные процессы могут быть объяснены в терминах микроэволюции, которые протекают достаточно быстро, чтобы человек мог наблюдать их в экспериментах или в природе.
Генетические идеи проникли в систематику, палеонтологию, эмбриологию, биогеографию. Из названия книги Джулиана Хаксли «Evolution: The Modern synthesis»[7] в научную литературу проник термин «современный синтез», обозначивший новый подход к эволюционным процессам. Выражение «синтетическая теория эволюции» в точном приложении к данной теории впервые было использовано Джорджем Симпсоном в 1949 году. Эта теория стала основой для развития эволюционной биологии во второй половине XX века. Подавляющее число новых идей в этой области рождалось из дискуссий вокруг синтетической теории, причём как из её защиты, так и из критики.
Методы эволюционной биологии
Эволюционная биология широко использует методы смежных наук. Опыт, накопленный палеонтологией, морфологией, генетикой, биогеографией, систематикой и другими дисциплинами, стал той базой, которая позволила превратить метафизические идеи о развитии живых существ в научный факт. Далее приводится описание различных методов приблизительно в той последовательности, в которой они входили в исследования по эволюции.
Палеонтологические методы
Практически все методы палеонтологии применимы для изучения эволюционных процессов[8]. Наибольшую информацию палеонтологические методы дают о состоянии биосферы на различных этапах развития органического мира вплоть до современности, о последовательности смен флор и фаун. Важнейшие из этих методов: выявление ископаемых промежуточных форм, восстановление филогенетических рядов и обнаружение последовательности ископаемых форм.
Биогеографические методы
Биогеографические методы основаны на анализе распространения ныне существующих видов, что даёт информацию о местонахождении очагов происхождения таксонов, путях их расселения, влиянии климатических условий и изоляции на развитие видов. Особое значение имеет изучение распространения реликтовых форм[8].
Морфологические методы
Морфологические (сравнительно-анатомические, гистологические и др.) методы позволяют на основе сравнения сходств и различий в строении организмов судить о степени их родства. Методы сравнительной анатомии, наряду с палеонтологическими, были одними из первых, позволивших поставить эволюционные представления на рельсы биологической науки.
Молекулярно-генетические методы
Макромолекулярные данные, под которыми имеется в виду последовательности генетического материала и белков, накапливаются всё быстрыми темпами благодаря успехам молекулярной биологии. Для эволюционной биологии быстрое накопление данных последовательностей целых геномов имеет значительную ценность, потому что сама природа ДНК позволяет использовать его как «документ» эволюционной истории. Сравнения последовательности ДНК разных генов у разных организмов могут сказать учёному много нового об эволюционных взаимоотношениях организмов, которые не могут иначе быть обнаружены на основе на морфологии, или внешней форме организмов, и их внутренней структуре. Поскольку геномы эволюционируют через постепенное накопление мутаций, количество отличий последовательности нуклеотидов между парой геномов разных организмов должно указать, как давно эти два генома разделили общего предка. Два генома, которые разделились в недавнем прошлом, должны иметь меньшие отличий, чем два генома, чей общий предок очень давний. Потому, сравнивая разные геномы друг с другом, возможно получить сведения об эволюционном взаимоотношения между ними. Это является главной задачей молекулярной филогенетики.
Теоретическая эволюционная биология
В современной эволюционной биологии сосуществует несколько теорий, описывающих эволюционные процессы. Такое сосуществование, хотя и не всегда мирное, объясняется тем, что каждая из теорий уделяет основное внимание ограниченной группе факторов. Так синтетическая теория делает упор на популяционно-генетические процессы, а эпигенетическая — на онтогенетическое развитие. Проблемы, связанные с эволюцией биоценозов как целого, освещает экосистемная теория эволюции, находящаяся в начальной стадии разработки. В то же время теория прерывистого равновесия даёт представление о сменах режимов эволюционного процесса, хотя мало что может сказать об их причинах.
Синтетическая теория эволюции
Синтетическая теория в её нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX века. После переоткрытия законов Менделя (в 1901 г.), доказательства дискретной природы наследственности и особенно после создания теоретической популяционной генетики трудами Р. Фишера (1918—1930), Дж. Б. С. Холдейна-младшего (1924), С. Райта (1931; 1932), учение Дарвина приобрело прочный генетический фундамент.
Статья С. С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926) по сути стала ядром будущей синтетической теории эволюции и основой для дальнейшего синтеза дарвинизма и генетики. В этой статье Четвериков показал совместимость принципов генетики с теорией естественного отбора и заложил основы эволюционной генетики. Главная эволюционная публикация С. С. Четверикова была переведена на английский язык в лаборатории Дж. Холдейна, но никогда не была опубликована за рубежом. В работах Дж. Холдейна, Н. В. Тимофеева-Ресовского и Ф. Г. Добржанского идеи, выраженные С. С. Четвериковым, распространились на Запад, где почти одновременно Р. Фишер высказал очень сходные взгляды о эволюции доминантности.
Толчок к развитию синтетической теории дала гипотеза о рецессивности новых генов. Говоря языком генетики второй половины XX века, эта гипотеза предполагала, что в каждой воспроизводящейся группе организмов во время созревания гамет в результате ошибок при репликации ДНК постоянно возникают мутации — новые варианты генов.
Нейтральная теория молекулярной эволюции
В конце 1960-х годов Мотоо Кимурой была разработана теория нейтральной эволюции, предполагающая, что в эволюции важную роль играют случайные мутации, не имеющие приспособительного значения. В частности, в небольших популяциях естественный отбор, как правило, не играет решающей роли. Теория нейтральной эволюции хорошо согласуется с фактом постоянной скорости закрепления мутаций на молекулярном уровне, что позволяет, к примеру, оценивать время расхождения видов.
Теория нейтральной эволюции не оспаривает решающей роли естественного отбора в развитии жизни на Земле. Дискуссия ведётся касательно доли мутаций, имеющих приспособительное значение. Большинство биологов признают ряд результатов теории нейтральной эволюции, хотя и не разделяют некоторые сильные утверждения, первоначально высказанные Кимурой. Теория нейтральной эволюции объясняет процессы молекулярной эволюции живых организмов на уровнях не выше организменных. Но для объяснения прогрессивной эволюции она не подходит по математическим соображениям. Исходя из статистики для эволюции, мутации могут как возникать случайно, вызывая приспособления, так и те изменения, которые возникают постепенно. Теория нейтральной эволюции не противоречит теории естественного отбора, она лишь объясняет механизмы проходящие на клеточном, надклеточном и органном уровнях.
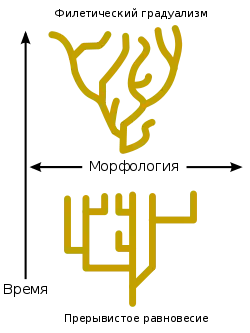
Теория прерывистого равновесия
В 1972 году палеонтологами Нильсом Элдриджем и Стивеном Гулдом была предложена теория прерывистого равновесия, утверждающая, что эволюция существ, размножающихся половым путём, происходит скачками, перемежающимися с длительными периодами, в которых не происходит существенных изменений. Согласно этой теории, фенотипическая эволюция, эволюция свойств, закодированных в геноме, происходит в результате редких периодов образования новых видов (кладогенез), которые протекают относительно быстро по сравнению с периодами устойчивого существования видов. Теория стала своеобразным возрождением сальтационной концепции. Принято противопоставлять теорию прерывистого равновесия теории филетического градуализма, которая утверждает, что бо́льшая часть процессов эволюции протекает равномерно, в результате постепенной трансформации видов.
Эволюционная биология развития
В последние десятилетия эволюционная теория получила импульс от исследований в области биологии развития. Открытие hox-генов и более полное понимание генетического регулирования эмбриогенеза стало основой для глубокого продвижения в теории морфологической эволюции, связи индивидуального и филогенетического развития, эволюции новых форм на основе прежнего набора структурных генов.
Экспериментальная эволюционная биология
-PLoS.jpg.webp)
Опыты Шапошникова
В конце 1950-х — начале 1960-х годов советским биологом Георгием Шапошниковым была проведена серия экспериментов, в процессе которых проводилась смена кормовых растений у различных видов тлей. Во время опытов впервые наблюдалась репродуктивная изоляция использованных в эксперименте особей от исходной популяции, что свидетельствует об образовании нового вида.
Эксперимент по эволюции E. coli
Уникальный эксперимент по эволюции бактерии E. coli в искусственных условиях, проведённый группой под руководством Ричарда Ленски в университете штата Мичиган. В процессе эксперимента прослежены генетические изменения, происходившие в 12 популяциях E. coli на протяжении 60 000 поколений. Эксперимент начался 24 февраля 1988 года и продолжается более 25 лет[9][10]
См. также
Примечания
- Четвериков С. С. О некоторых моментах эволюционного процесса с точки зрения современной генетики // Журн. эксп. биол. — 1926. — Т. 2. — С. 3—54.
- Fisher R. A. The Genetical Theory of Natural Selection. — Oxford: Clarendon Press, 1930.
- Wright S. Evolution in Mendelian populations (англ.) // Genetics. — 1931. — Vol. 16. — P. 97—159.
- Haldane J. B. S. The Causes of Evolution. — London: Longmans, Green & Co., 1932.
- Иорданский Н. Н. Эволюция жизни. — М.: Академия, 2001. — 425 с.
- Dobzhansky Th. Genetics and the Origin of Species. — New York: Columbia University Press, 1937.
- Huxley J. Evolution: The Modern synthesis. — London: Allen & Unwin, 1942.
- Яблоков А.В., Юсуфов А.Г. Эволюционное учение. — М.: Высшая школа, 2006. — 310 с.
- Richard E. Lenski Source of founding strain, 2000. Accessed June 18, 2008.
- Jeffrey E. Barrick, Dong Su Yu, Sung Ho Yoon, Haeyoung Jeong, Tae Kwang Oh, Dominique Schneider, Richard E. Lenski, Jihyun F. Kim. Genome evolution and adaptation in a long-term experiment with Escherichia coli (англ.) // Nature : journal. — 2009. — Vol. 461. — P. 1243—1247.
Литература
Научно-популярная
- Еськов К. Ю. История Земли и жизни на ней: От хаоса до человека. — М.: НЦ ЭНАС, 2004. — ISBN 5-93196-477-0.
- Марков А. В. Рождение сложности. — М.: Астрель, 2010. — 527 с. — 4000 экз. — ISBN 978-5-271-24663-0.
Учебная и научная
- Воронцов Н. Н. История эволюционного учения. — М.: Изд.отдел УНЦ ДО МГУ, 1999. — 640 с.
- Грант В. Эволюционный процесс. Критический обзор эволюционной теории. — М.: Мир, 1991. — 488 с. Архивная копия от 9 апреля 2012 на Wayback Machine
- Иорданский Н. Н. Эволюция жизни. — М.: Академия, 2001. — 425 с.
- Северцов А. С. Теория эволюции. — М.: Владос, 2005. — 380 с. — 10 000 экз.
- Солбриг О., Солбриг Д. Популяционная биология и эволюция. — М.: Мир, 1982. — 244 с.
- Колчинский Э. И. Неокатастрофизм и селекционизм: вечная дилемма или возможность синтеза?. — СПб.: Наука, 2002. — 554 с. — ISBN 5-02-026174-2.
На английском языке
- Futuyma D. J. Evolution. — Sunderland: Sinauer Associates, 2005. — ISBN 0-878-93187-2.
- Gould S. J. The Structure of Evolutionary Theory. — Cambridge: Belknap Press of Harvard University Press, 2002. — 1443 экз. — ISBN 978-0-674-00613-3.
- Mayr E. What Evolution Is. — New York: Basic Books, 2001. — 336 p.
- Ridley M. Evolution. — 3rd ed. — Wiley-Blackwell, 2004. — 751 p. — ISBN 978-1-4051-0345-9.
- Shapiro J. Evolution: A View from the 21st Century. — FT Press Science, 2011. — 253 p. — ISBN 978-0-13-278093-3.
Ссылки
- Сайт кафедры биологической эволюции биологического факультета МГУ
- Сайт «Проблемы эволюции»
- Сайт «Антропогенез.РУ»
| Эволюционные процессы | |
|---|---|
| Факторы эволюции | |
| Генетика популяций | |
| Происхождение жизни | |
| Исторические концепции | |
| Современные теории | |
| Эволюция таксонов | |