Стрижеобразные
Стрижеобра́зные[1][2], или длиннокрылые[1] (лат. Apodiformes), — отряд новонёбных птиц, в состав которого включают семейства стрижей, древесных стрижей и колибри. Стрижеобразные — мелкие или очень мелкие птицы с крупной головой, крепким телом и острыми крыльями. Характерными признаками представителей отряда являются удлинённая дистальная часть крыла (пряжка) и укороченная проксимальная (плечевая, лучевая и локтевая кости). Птицы способны к быстрому и маневренному полёту, но их слабые ноги непригодны для ходьбы по земле, а строение лапы настоящих стрижей не позволяет им даже сидеть на ветках. Подавляющее большинство представителей отряда обитают в тропиках, лишь немногие освоили умеренные широты. Настоящие стрижи распространены по всему свету за исключением полярных районов, колибри населяют Новый Свет, включая острова Карибского моря, древесные стрижи — Юго-Восточную Азию, многочисленные прибрежные острова и Новую Гвинею.
| Стрижеобразные | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Чёрный стриж, иллюстрация 1873 года | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Подкласс: Инфракласс: Клада: Отряд: Стрижеобразные |
||||||||||||
| Международное научное название | ||||||||||||
| Apodiformes Peters, 1940 | ||||||||||||
| Семейства | ||||||||||||
|
||||||||||||
| ||||||||||||
Настоящие стрижи — шумные и общительные птицы, которые часто собираются в большие стаи, устраивают гнездовые колонии. Для колибри в основном характерно территориальное поведение, но представители некоторых видов игнорируют друг друга. Представители отряда могут впадать в оцепенение на ночь или на несколько дней, чтобы понизить скорость обмена веществ и пережить периоды голодания в ненастную погоду. Основу рациона составляют насекомые, у колибри — нектар.
С конца XIX века обнаружено много ископаемых остатков птиц, по анатомическому строению схожих со стрижами. Определению их систематического положения в пределах отряда мешает отсутствие оперения, так как для классификации важны длина перьев и пропорции длины крыла относительно тела: у стрижей первостепенные маховые перья длиннее, а у колибри длина крыла пропорционально короче. Предполагается, что предки стрижеобразных отделились от праракшеобразных в позднемеловой эпохе около 70 млн лет назад.
Систематика
| Кладограмма Caprimulgiformes + Apodiformes по Hackett et al. (2008)[3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ещё до появления молекулярно-филогенетических данных господствовало мнение, что стрижиные наиболее близкородственны колибри, и семейства настоящих и древесных стрижей объединялись с колибри в отряд стрижеобразных (Apodiformes)[4], выделенный американским орнитологом Джеймсом Ли Питерсом в 1940 году[5]. Их близкое родство подтвердил анализ ДНК-ДНК-гибридизации, проведённый Ч. Сибли и Дж. Алквистом[4]. Отряд стрижеобразных традиционно разделяют на два подотряда[6][7]. Подотряд стрижей (Apodes) включает семейство стрижиных (Apodidae) (113 видов[8]) и семейство хохлатых, или древесных, стрижей (Hemiprocnidae) (4 вида[8]), а подотряд колибри (Trochili) — семейство колибри (Trochilidae) (361 вид[9]). Некоторые исследователи относят древесных стрижей к семейству Apodidae[7], другие считают их близкими к совиным козодоям (Aegothelidae)[10]. По классификации Сибли — Монро, отдельные отряды стрижеобразных и колибриобразных (Trochiliformes)[3] формируют надотряд Apodimorphae[7]. Существует также точка зрения, что у колибри и стрижей нет достаточно близкого общего предка[10][11].
По ряду признаков стрижеобразные схожи с козодоеобразными[4][12], в частности с настоящими козодоями, а также с птицами-мышами[12]. Некоторые исследователи обнаруживают сходство стрижей и колибри с воробьинообразными и предлагают включить их в этот отряд как подотряды[12]. Немецкий палеонтолог Геральд Майр показал в 2008 году, что отряд козодоеобразных в классическом объёме является полифилетическим, в нём семейство совиных козодоев отделено от остальной части отряда монофилетическим[13] отрядом стрижеобразных[3]. Ещё в 2002 году он предположил, что это семейство может представлять собой сестринскую группу по отношению к стрижеобразным[13]. На основе этих данных Международным союзом орнитологов было принято решение выделить семейство совиных козодоев (9 видов[8]) в отдельный отряд Aegotheliformes[8]. В справочнике Handbook of the birds of the world настоящие стрижи, древесные стрижи и колибри, наряду с совиными козодоями, а также настоящими козодоями, исполинскими козодоями, лягушкоротами и гуахаро, включены в обширный монофилетический[3] отряд козодоеобразных[14]. По предположению Сибли и Алквиста, совообразные, козодоеобразные и стрижеобразные образуют монофилетическую группу[4].
Определение отряда
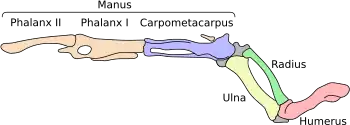

Майр в работах 2002 и 2003 годов сформулировал восемь признаков отряда стрижеобразных, включающего совиных козодоев[15].
- (i) нёбная кость (os palatinum) имеет сильно выпуклый угол между боковой и задней стенкой (angulus caudolateralis);
- (ii) базиптеригоидные сочленения (processus basipterygoidei) редуцированы;
- (iii) на задней поверхности глазного отростка (processus oticus) имеется пневматическое отверстие;
- (iv) у коракоида верхний конец (extremitas omalis) загнут, внешняя часть (processus lateralis) сильно редуцирована;
- (v) у ременной мышцы головы (musculus splenius capitis) крестообразное основание;
- (vi) слепая кишка (caeca) отсутствует;
- (vii) нижняя часть (processus terminalis) седалищной кости (ischiiof pelvis) очень узкая и тонкая, стыкуется с лобковой костью под углом 45—90°, седалищная вырезка (fenestra ischiopu-bica) очень широкая;
- (viii) длинная малоберцовая мышца (musculus fibularis longus) отсутствует.
В более поздних работах признак (v) был назван синапоморфным, а признаки (iii), (iv), (vi) — (viii), хоть и не являются уникальными для представителей отряда, по мнению Джорджа Сангстера (George Sangster), независимо эволюционировали у его общего предка[15].
Описание
Размеры
Стрижеобразные — птицы малых или очень малых размеров[1]. Размеры колибри составляют 5—22 см[16], стрижей — 9—25 см[17], древесных стрижей — 15 — 31 см [18]. Колибри — самые маленькие птицы на Земле, первое место занимает колибри-пчёлка (Mellisuga helenae) с Кубы, расстояние от кончика клюва до кончика хвоста которой составляет 62 мм[10].
Масса птиц в основном составляет от 1,6 до 50 г[6]. Большинство колибри имеют массу тела 2,5—6,5 г (по другим данным, 3—7 г[19]). Самые лёгкие представители колибри — краснобрюхий солнечный колибри (Phaethornis ruber) из Гайаны и Бразилии[20] и колибри-пчёлка с Кубы — весят 1,6—1,9 г[19]. На 12—14 г тяжелее средней массы колибри орлиноклювые колибри (Eutoxeres), колибри-мечеклювы (Ensifera) и голубокрылые колибри (Pterophanes)[20], а самый тяжёлый представитель семейства — исполинский колибри (Patagona gigas) — весит 19—21 г (по другим данным, 18—20 г[19]). Масса стрижей также сильно варьирует: карликовая салангана (Collocalia troglodytes) весит 5 г. Некоторые виды рода стрижи (Apus) в 20 раз тяжелее[10], а масса фиолетового колючехвоста[12] (Hirundapus celebensis) может достигать 200 г[12].

_(8127975211).jpg.webp) Белобрюхая салангана
Белобрюхая салангана
Оперение
Представители отряда характеризуются прежде всего формой и пропорциями крыла[10]. Дистальная часть крыла — кисть и первостепенные маховые перья — у них очень длинная, а проксимальная — плечо и предплечье — напротив, укороченная[1][10]. Плечевая кость толстая, с хорошо развитым отростком для крепления мышц. Первостепенных маховых перьев обычно десять, первое и второе составляют острую вершину крыла[1], при этом у многих колибри их размер последовательно увеличивается от внутренней части крыла к внешней[20]. Второстепенных маховых перьев 6—7 у колибри и 8—11 у стрижей, первостепенные маховые перья намного длиннее второстепенных[1][10]. Длина самого длинного первостепенного пера стрижа может в три раза превышать длину самого короткого второстепенного[21].
Хвост колибри[20] и настоящих стрижей[12] обычно короткий, для древесных стрижей характерен длинный хвост с глубоким вырезом[22]. Рулевых перьев чаще всего десять[1]. Чёрный стриж (Apus apus) обладает двенадцатью рулевыми перьями[21], а колибри лоддигезия (Loddigesia mirabilis) — только четырьмя[20]. У большинства колибри хвостовые перья невелики, но у некоторых сильно вытянуты. У стрижей хвосты существенно различаются: у представителей трибы колючехвосты (Chaeturini) центральный стержень хвостовых перьев выступает за лопасти и выглядит как шипы, у остальных стрижей хвост может быть квадратным, умеренно или глубоко раздвоенным[10]. Легко узнаваемы очертания древесных стрижей на насесте: кончики хвоста у них выступают за глубоко скрещенные кончики крыльев. Длина хвоста составляет от 58 до 85 % длины крыла, а глубина разреза больше, чем у любого из настоящих стрижей[22].
Колибри ярко окрашены, в то время как стрижи — напротив, обладают однообразным, тёмным оперением[6]. Изумрудно-зелёная окраска колибри, их радужные горловые пятна, хохолки и некоторые другие участки тела обусловлены меланиновой матрицей, расположенной под тонким верхним слоем в перьевых бороздочках[20]. Цвет крыльев колибри основан на пигментах, как и у почти всех других птиц, и редко бывает ярким, но может быть матово-охристым, буроватым или чёрным[23]. Оперение стрижей в чёрной или коричневой цветовой гамме, может сочетать чёрные и белые цвета. Исключение составляют близкородственные тропические виды красношейный американский стриж (Streptoprocne rutila) и венесуэльский стрижик (Streptoprocne phelpsi) с ярко-рыжим горлом, подбородком и ошейником[12][21]. Древесные стрижи отличаются от настоящих стрижей хохлом и маской на лице[12].
Контурные перья имеют добавочные стержни. На аптериях у стрижей имеется короткий пух, а у колибри он полностью отсутствует[1].
Анатомия

Обычно у птиц крупная голова и короткая шея. У стрижей череп эгитогнатический — нёбные отростки верхнечелюстных костей тонкие и удлинённые, они подходят под сошник, но не соприкасаются ни с ним, ни друг с другом, у широкого сошника свободный передний конец иногда раздвоён, а задний глубоко расщеплён и охватывает клювовидный отросток. У колибри же череп схизогнатический — отсутствует соединение нёбных отросков правой и левой верхнечелюстных костей, сочленение нёбных и крыловидных костей подвижное, они примыкают к клювовидному отростку, отсутствует соединение сошника, заострённого спереди, с нёбными отростками верхнечелюстных костей. Базиптеригоидные сочленения, связывающие нёбноквадратные хрящи с птеригоидами, в черепе отсутствуют[1]. Ноздри округлые, не сквозные[10][1]. У стрижей ноздри не покрытые, а у колибри покрытые[10]. Клюв различных форм и размеров[10][1]. Так, у колибри он обычно длинный, прямой или в разной степени загнутый вниз, но иногда может быть загнут и вверх. Рот не может широко открываться, язык длинный. Для подотряда стрижей характерны короткие клюв и язык, крупные слюнные железы[10]. Челюстной сустав стрижей находится далеко за глазом, что позволяет им широко открывать рот[24].
У многих стрижеобразных зоб отсутствует, исключение составляют птенцы колибри и некоторые взрослые представители этого семейства. Правая сонная артерия отсутствует[10][1], но имеется у стрижей подсемейства Cypseloidinae[10]; копчиковая железа голая. Желчный пузырь присутствует у стрижей и отсутствует у колибри. Слепая кишка отсутствует или сильно редуцирована. На нижней гортани трахеобронхиального типа расположены одна или две пары голосовых мышц. У стрижей 13 шейных позвонков[1] и 6—7 пар рёбер[10], а у колибри — 14—15[1] и 8, соответственно[10].
Система мускулов на ногах упрощена, цевка сильно укорочена, с частичным или полным оперением[1]. Из-за слабых и маленьких ног стрижи не могут ходить, садиться на ветки и провода, но могут цепляться за вертикальные поверхности[10][1]. Их различают по положению пальцев и оперенности лап[10]. У древесных стрижей лапа анизодактильная — четвёртый палец направлен назад[7]. Древесные стрижи любят сидеть на голых ветках деревьев, именно с них бросаются в погоню за насекомыми. Колибри могут сидеть на ветке спокойно и проводят в воздухе меньше времени, чем стрижи, однако передвигаются по земле они с трудом и предпочитают перелёты даже на короткие расстояния[10].
Поведение

_hunting_by_echolocation.JPG.webp)
Неотъемлемой частью поведения представителей отряда является их полёт[10]. Стрижеобразные — птицы воздуха, на лету они принимают пищу, пьют воду, купаются, собирают материал для гнезда, проводят ритуалы ухаживания, некоторые виды могут спать[10][6] или совокупляться в полёте[25]. Стрижеобразные способны к чрезвычайно быстрому и маневренному машущему полёту[1]. Жёсткие и быстрые взмахи крыльев стрижей чередуются со скольжением по воздуху[24]. Высокая скорость достигается за счёт первостепенных и второстепенных маховых перьев[21]. Некоторые крупные стрижи могут летать со скоростью до 170 км в час[21][24], но в отличие от ласточек они не способны к медленному полёту[25]. Полёт древесного стрижа представляет собой комбинацию сложных манёвров ласточек и высокой скорости стрижей[26]. У усатого древесного стрижа нагрузка на крыло меньше, чем у представителей стрижей (Apus) и колючехвостых стрижей (Hirundapus), а их высокая манёвренность связана с длинным хвостом, похожим на хвост ласточек[22]. Колибри в полёте могут зависать на месте, летать в обратном направлении или вверх ногами[10]. Во время трепещущего полёта колибри их крылья перемещаются главным образом в горизонтальной плоскости и постоянно перекручиваются, описывая концами плоскую восьмёрку. Чем меньше размер колибри, тем чаще птица машет крыльями. Во время брачного полёта частота взмахов может достигать 100 в секунду[27]. В лабораторных условиях скорость птиц составляет 48—85 км в час и даже 150 км в час[20].
Настоящие стрижи часто собираются в стаи, которые могут объединять представителей разных видов. Многие виды устраивают многочисленные гнездовые колонии, но даже гнездящиеся отдельно в другое время года собираются вместе. Некоторые ночные скопления включают сотни птиц. Для колибри, напротив, характерно территориальное поведение, они защищают свои кормовые и гнездовые территории. При этом колибри могут атаковать гораздо более крупных птиц, включая воронов и ястребов. Вместе с тем, у некоторых видов территориальность отсутствует полностью. Они могут кормиться вместе на одном дереве, игнорируя друг друга. На одном дереве могут питаться до шести видов колибри[10].
Древесные стрижи в основном активны в сумерках. Известно, что крупные виды интенсивно кормятся рано утром и поздно вечером, а усатый древесный стриж совершает вылазки и после наступления темноты. Кроме того, все представители семейства особенно активны после дождя[26]. Стрижи ночью отдыхают, но спят некрепко. Обычно они просыпаются позже восхода солнца, при этом северные популяции стрижей поднимаются намного позже, чем южные. Суточная активность стрижей связана с распределением насекомых[28].
Некоторые виды стрижей способны ориентироваться в полёте с помощью эхолокации[6][10], они строят гнёзда в полной темноте глубоких пещер и могут охотиться дольше остальных птиц. Средм всех птиц помимо стрижей эхолокация встречается только у гуахаро (Steatornis caripensis). Обычно эхолокационные сигналы стрижей находятся в пределах спектра, доступного для человеческого слуха[10][29], в отличие от звуков, используемых летучими мышами[29]. Необходимым дополнением к эхолокационным способностям стрижей является хорошо развитая пространственная память[21].
И стрижи, и колибри способны снижать температуру тела и впадать в оцепенение для сохранения энергии[6][10]. Колибри используют такой способ сохранения энергии ночью, а стрижи — во время периодов длительного голодания[10], а также непогоды (похолоданий или затяжных дождей). Снижение высокого уровня обмена веществ позволяет стрижам переживать голодание до 5—12 дней. Особенно это характерно для птенцов в гнезде[7]. Оцепенение колибри может продолжаться 15—20 часов, иногда — до нескольких дней[30]. Во время оцепенения колибри не способны реагировать на внешние раздражители и могут попасться хищникам, их поведение становится летаргическим — колибри могут не проснуться[20]. Оцепенение у северных колибри возникает не регулярно, а только когда уровень внутренней энергии опускается ниже порогового значения. Возможно, это связано с большей продолжительностью светового дня, чем в тропиках, где в оцепенение впадают почти все колибри[20].
Распространение
Ареал

Подавляющее большинство представителей отряда обитают в тропиках, лишь немногие освоили умеренные широты[7].
Настоящие стрижи широко распространены, встречаясь и в Старом, и в Новом Свете[10]. Вне пределов ареала семейства лишь полярные районы (Арктика, Антарктика, Гренландия, Исландия и север Сибири[31]), южная часть Чили и Аргентины, Новая Зеландия, почти вся Австралия[25], некоторые группы островов[32], а также центральные районы Сахары и Тибета[31]. Высокому распространению стрижей способствует их чрезвычайная мобильность. Так, некоторые стрижи обитают на океанических островах, которые никогда не были частью континентальной суши[32]. Единственным ограничением для распространения птиц является достаточное для пропитания количество летающих насекомых. Сильнее всего семейство представлено в тропиках, где встречаются виды с перекрывающимися ареалами (симпатрические), иногда близкородственные. Например, на Тринидаде обитают три очень похожих вида рода иглохвосты (Chaetura). О распределении экологических ресурсов и возможном высотном разделении симпатрических видов стрижей практически ничего не известно[10].
Древесные стрижи обитают только в Южной и Юго-Восточной Азии, включая Зондские и Филиппинские острова, Новую Гвинею и прилегающие острова[7][10]. Ареалы различных видов древесных стрижей в основном не пересекаются, только ушастый и блестящий клехо делят обширную общую территорию в Юго-Восточной Азии[33].
Колибри, напротив, обитают исключительно в Новом Свете. Европейские нектарницевые (Nectariniidae) внешне напоминают колибри, но не находятся с ними в родстве[10]. Колибри встречаются в Центральной, Северной и Южной Америке, а также на островах Карибского моря, на высотах до 5200 метров над уровнем моря[34]. Большинство видов колибри обитают в тропиках, где цветение происходит круглый год, только некоторые живут в умеренном климате или на высокогорье[10]. В Эквадоре зарегистрировано 163 вида колибри, в Колумбии — 135, в Перу — 100, а в Бразилии — 84[35].
Миграция
Северные стрижи, в частности чёрный стриж (Apus apus) и дымчатый иглохвост (Chaetura pelagica), осуществляют продолжительные миграции. Даже тропические виды стрижей могут перемещаться на большие расстояния[10]. Птицы обычно мигрируют очень быстро и летят очень высоко, с поверхности Земли их можно увидеть, только если плохие погодные условия заставят стаю снизить высоту. Некоторые птицы могут лететь и ночью. Птицы могут мигрировать широким фронтом, используя миграционные коридоры только над морем. Стрижи могут лететь впереди штормовых фронтов, пользуясь изобилием воздушного планктона, вызванным конвекцией воздуха. Во время сезона размножения такие стаи состоят в основном из птиц, не сделавших кладки, но в них могут участвовать и родители, которые оставляют птенцов на некоторое время без внимания и пищи[32].
Характер и масштаб миграций колибри мало изучен. Северные колибри, как и северные стрижи, являются мигрирующими птицами. Рубиновогорлый колибри (Archilochus colubris) во время миграции пересекает Мексиканский залив, а чилийский огнешапочный колибри (Sephanoides sephaniodes) достигает Огненной Земли[10]. Некоторые колибри используют разные маршруты для осенней и весенней миграции[36]. Высокогорные колибри могут совершать высотные кочёвки[37], мигрируя на более низкие высоты, часто в прибрежные районы. При этом некоторые виды, в частности колибри — горные звёзды (Oreotrochilus), остаются высоко в горах круглый год и перемещаются ниже только в самых суровых условиях. Как правило, колибри-отшельники мигрируют меньше, чем типичные колибри[36].
Древесные стрижи ведут преимущественно оседлый образ жизни. Сезонные перемещения могут осуществлять хохлатые и блестящие клехо, локальные кочёвки возможны у усатого древесного стрижа. Ушастый клехо остаётся на своей кормовой территории в течение всего года[38].
Среда обитания

Среда обитания стрижей самая разнообразная: они населяют горы и равнины, открытые пространства и леса, гнездятся на скалах, в пещерах, в норах или в дуплах, освоили ниши зданий, чердаки и дымоходы[39]. Стрижи обитают в местах с достаточной концентрацией насекомых в воздухе и доступными местами для ночлега и размножения. Чаще всего они встречаются около воды, особенно зависимы от неё представители подсемейства Cypseloidinae и Hydrochous gigas. При этом бледный стриж может иногда проникать в засушливые районы, встречаясь в оазисах в центре Сахары, а белобрюхий мохноногий стриж размножается в засушливых районах на юге США и на севере Мексики, но добывает пищу как правило на влажных лугах неподалёку. Некоторые стрижи предпочитают селиться на пальмах, баобабах или в реликтовых лесах[40]. Большинство древесных стрижей селятся на деревьях, возвышающихся над пологом леса, обычно там, где за лесом находится открытое пространство — русла рек, расчищенные ветром территории, системы автодорог, сельскохозяйственные площади[41]. Самая большая плотность хохлатых и блестящих клехо, а также, по-видимому, усатых древесных стрижей приходится на зрелые парковые посадки и леса около сельскохозяйственных участков. Однако даже в лесопосадках древесные стрижи предпочитают местные деревья, в частности дуриан[42]. В то время как ушастый клехо в основном обитает в глубине леса, остальные три вида живут на его краю[43].
Колибри широко распространены в тропических лесах, особенно многочисленны в среднегорье, лишь несколько видов обитают в умеренных широтах. Ареалы многих видов колибри ограничены одной долиной или склоном, некоторые тянутся узкой полосой вдоль западного или восточного склона Анд, много островных эндемиков. Предпочитают богатые растительностью биотопы, однако известны виды, характерные для пустынь и полупустынь[34]. На разных высотах таксоны экологически сменяются другими подобными. В регионах большого видового разнообразия колибри существенное значение также имеют специфичные для разных видов стратегии кормления[44]. Многие виды колибри легко адаптируются к изменениям среды обитания, а при благоприятных условиях могут даже расширять свои ареалы[44].
Питание
Рацион
_feeding.jpg.webp)
Основу питания стрижей и древесных стрижей составляют различные членистоногие. Колибри питаются цветочным нектаром, который они высасывают языком, как росу или дождевую воду, а мелких насекомых и пауков ловят клювом[10].
Стрижи питаются в основном «воздушным планктоном» — плохо летающими мелкими насекомыми, пассивно дрейфующими в воздушных потоках, или мелкими паучками, использующими паутинки для расселения[24]. Рацион стрижей не имеет точного соответствия составу насекомых в местности — в разных условиях основную часть корма составляют разные группы членистоногих. Обычно птицы избегают жалящих насекомых, но питаются насекомыми, которые мимикрируют под них. Тем не менее, были обнаружены особи белобрюхого стрижа с укусами насекомых на горле. Анализ содержимого желудков и болюсов показал, что его основу составляют перепончатокрылые (Hymenoptera), в частности пчёлы, осы, муравьи, двукрылые, полужесткокрылые, жесткокрылые. У мелких стрижей рацион более разнообразен, чем у крупных. Крупные стрижи ловят меньшее количество насекомых, как правило, более крупного размера[45]. Особенности питания древесных стрижей изучены очень слабо, какие-либо количественные данные отсутствуют полностью. В их рацион входят пчёлы, муравьи, полужесткокрылые, жесткокрылые, термиты. Неясно, как древесные стрижи справляются с пчелиным ядом и «химической защитой» настоящих щитников (Pentatomidae)[26].
В ходе эволюции возникла тесная взаимосвязь между некоторыми цветковыми растениями и колибри — орнитофилия. Так, многие растения Нового Света опыляются исключительно колибри. Они смещают время цветения, обеспечивая птиц нектаром в любое время года. Как растения, так и птицы — участники таких взаимоотношений представлены множеством видов; по всей видимости, парной коэволюции отдельных видов колибри и отдельных видов растений не происходило[46]. Чаще всего колибри потребляют нектар цветов двудольных многолетних травянистых растений и кустарников, крайне редко — деревьев. Обычно растения — поставщики нектара относятся к родам Zauschneria, живокость (Delphinium), водосбор (Aquilegia), губастик (Mimulus), афеландра (Aphelandra), Centropogon, кавендишия (Cavendishia), колумнея (Columnea), Psittacanthus и геликония (Heliconia). Цветущие деревья, например эритрина (Erythrina), являются большим источником нектара и обычно занимаются территориальными колибри в самом начале цветения. Колибри также потребляют пыльцу, содержащую необходимые аминокислоты, и членистоногих в качестве белковой добавки. В среднем ежедневное соотношение потребления нектара и белковой пищи по массе составляет девять к одному[46]. Вместе с тем, иногда белковая пища составляет до трети рациона[47].
Особенности охоты

Вероятно, на ранней стадии эволюции стрижи бросались за добычей с насеста, как мухоловки (Muscicapidae) и древесные стрижи, затем стали охотиться в воздухе за отдельными насекомыми, как ласточки, а впоследствии освоили «траление»[24]. Стрижи добывают «воздушный планктон» в воздухе, пролетая через рой с открытым ртом. Крупные стрижи легче добывают роящихся насекомых в местах концентрации отдельных видов на более обширных территориях, что привело к их более узкой специализации. Мелкие стрижи также могут охотится на роящихся насекомых, но имеют ограниченные кормовые территории. Обычно их кладки превосходят по размерам кладки крупных стрижей и требуют частого кормления быстро растущего потомства, что не даёт птицам отлетать далеко от гнёзд[45]. Помимо размеров насекомых, стратификация различных видов стрижей также происходит за счёт выбора различных высот, на которых ловят добычу птицы[45]. Стрижи охотятся за насекомыми в большом интервале высот — в Европе примерно до 100 метров от земной поверхности (на бо́льших высотах насекомых значительно меньше). Согласно исследованиям, проведённым в Эквадоре, крупные стрижи охотятся на больших высотах, чем мелкие. Аналогичные результаты были получены около пещер Ниах в Малайзии и у водохранилища Кариба в Зимбабве[45]. Межвидовая конкуренция стрижей зависит от того, насколько широко они могут открывать рот[45]. Стрижи часто образуют крупные смешанные стаи с другими быстрыми птицами, в том числе ласточками, пользуясь кратковременной концентрацией пищи[28].
Древесные стрижи обычно ловят насекомых в полёте, но ушастый клехо может иногда снимать их с листьев, сидя на ветке. Птицы часто привязаны к кроне домашнего дерева, с которого они отправляются на охоту, эффективно используя намного меньшие кормовые территории, чем настоящие стрижи[26]. Древесные стрижи могут делить кормовые территории в любое время года, часто можно встретить вместе две или три пары, охотящиеся совместно. Исключение составляет ушастый клехо: птицы кормятся на небольшой территории исключительно парой, изредка сопровождаемые одним птенцом[43], и никогда не охотятся на открытом пространстве[42].
На выбор растений влияет морфология клюва колибри[46]. В этом отношении растения можно разделить на две категории. Мелкие колибри с коротким клювом предпочитают цветки с венчиками длиной 17—20 мм и частотой секреции нектара 1—2 микролитра сахарозы в день. Более крупные колибри с длинными клювами предпочитают цветки с венчиками длиной 30—38 мм и частотой секреции нектара 4—27 мкл сахарозы в день[44]. Кроме того, колибри с высокими энергетическими потребностями, связанными с большой нагрузкой на крылья, предпочитают кормовые территории вокруг соцветий, а колибри с низкими потребностями используют редко расположенные цветы или цветы с короткими венчиками (цветы насекомых)[46]. Некоторые колибри прокалывают венчик сбоку и потребляют нектар, не опыляя растений[46][47].
Размножение
_in_Kawal_WS%252C_AP_W_IMG_2133.jpg.webp)
Поведение в период ухаживания у стрижеобразных довольно разнообразно. Воздушные ритуалы характерны для колибри и настоящих стрижей. Яркие типичные колибри демонстрируют в воздухе своё оперение, менее яркие колибри-отшельники могут токовать группами, у них пение является особенно важной частью брачного поведения[10]. Стрижи совершают воздушные погони и синхронные манёвры в воздухе. Возможность синхронного полёта в таких парах служит индикатором дальнейшего успеха при размножении. При глубоком нырке может формироваться V-образная поза, когда оба крыла птицы высоко подняты над телом. Она особенно выражена в уже сформированных парах и обычно встречается у догоняющих птиц. В брачном полёте стрижи громко кричат[28]. Некоторые звуковые сигналы древесных стрижей также могут носить характер ухаживания[48].
Некоторые виды стрижей и древесных стрижей, возможно, совокупляются в воздухе. Такое поведение было зафиксировано у пары блестящих клехо[48], у чёрных стрижей, у белобрюхих мохноногих стрижей и у некоторых других представителей стрижиных[49]. В то же время известны описания усатых древесных стрижей[48] и чёрных стрижей[49], спаривающихся на насесте. По мнению Марина и Г. Стайлса, воздушное спаривание влечёт за собой большой риск неудачи и стерильности яиц, что невыгодно видам с небольшими кладками[49]. Специфическое поведение самки колибри, в особенности неподвижное сидение около самца, побуждает самца к последующей копуляции. Самцы при этом демонстрируют поведение, схожее с охранным[50].
Колибри в основном полигамны[10][50], а стрижи моногамны[39].
Гнёзда и гнездовые колонии

Стрижи устраивают многочисленные гнездовые колонии. Обычно они гнездятся в расщелинах или ямах, внутри полых деревьев или в пещерах, могут строить гнёзда в искусственных сооружениях (зданиях, колодцах, туннелях). Сами гнёзда птицы размещают на выступах или прикрепляют к опоре сбоку с помощью большого количества выделяемой липкой слюны. Гнездо изнутри представляет собой мелкую чашу из перьев и растительного материала, некоторые саланганы используют слюну как основной, а иногда и единственный, материал при строительстве гнезда[10][21][49]. Сделанные полностью из слюны гнёзда саланганы-водорослееда являются деликатесом в китайской кухне. Один килограмм включает 80—120 гнёзд, только в Индонезии собирают до 200 тонн гнёзд в год. Во время сезона гнёзда собираются дважды, так что для размножения птицы вынуждены их строить в третий раз, при этом из-за сильного истощения слюнных желёз гнёзда содержат кровь. Всё вместе это приводит к сокращению популяции стрижей со съедобными гнёздами[51]. Стрижи подсемейства Cypseloidinae и, возможно, колючехвостые стрижи (Hirundapus) не используют слюну при строительстве гнезда[49]. Они выскребают гнездо, скрепляют его с помощью грязи или крайне медленно строят на постепенно «прорастающем» мхе. Так как эти стрижи могут использовать хитин насекомых из погадок, возможно, вместе с ними они получают слюну[52]. Пальмовые стрижи нового (Tachornis) и старого (Cypsiurus) света размещают под пальмовыми листьями свои сложные мешкообразные гнёзда со входом снизу через трубчатый туннель. Белоголовый американский стриж (Streptoprocne semicollaris) может вообще не строить гнёзда, откладывая яйца в небольшом углублении на голом песке или на выступах внутри пещер[10].
Очень маленькие чашеобразные гнёзда древесных стрижей размещаются на тоненьких ветках высоко на деревьях и вмещают только одно яйцо[10][7], расположенное вертикально. Пустое гнездо напоминает естественный нарост на ветке, однако яйцо в нём хорошо заметно[48].
Строительством гнезда колибри — чаще всего чашеобразного, из растительных волокон, паутины, кусочков коры и лишайников — занимаются самки[10]. Типичные колибри обычно строят открытые гнёзда, в которые можно попасть с лёта. Их гнёзда обычно двуслойные: за внешним камуфляжным слоем из грубых материалов следует внутренний мягкий слой, способствующий оптимальной изолированности от внешней среды[50]. Гнёзда колибри-отшельников существенно отличаются, представляя собой подвешенные плетённые корзины[10] или другие конструкции с открытыми стенками. Плохая изолированность компенсируется высокой температурой окружающей среды, кроме того, сквозь такую конструкцию проходит много света, а дождевая вода протекает сквозь гнездо и не оказывает влияние на яйца[50]. Крайне редко колибри образуют гнездовые скопления[53].
Яйца

Размер кладки настоящих стрижей варьирует от одного до шести белых яиц, которые высиживают оба родителя. Большие кладки обычно встречаются у северных видов родов Chaetura и Aeronautes[10], у подсемейства Cypseloidinae в кладке одно или два яйца[54]. Древесные стрижи всегда откладывают только одно яйцо. Чтобы его не сдуло ветром, родители приклеивают яйцо ко дну гнезда слюной, вращение яйца во время насиживания заменяется колебанием ветки на ветру[10]. Подобным образом некоторые стрижи, в частности Cypsiurus parvus, также приклеивают яйца к гнезду[10]. У стрижей размеры яиц могут составлять от 15,5 × 10 мм у вилохвостого карликового стрижа до 43 × 28,5 мм у ошейникового американского стрижа[49]. Инкубационный период составляет 17—25 дней[55]. Размеры яиц древесных стрижей варьируют от 20 × 15 мм у ушастого клехо до 30 × 20 мм у усатого древесного стрижа. Птицы всегда откладывают одно белое или бледно-серое яйцо. Продолжительность инкубационного периода неизвестна[48].
Кладка колибри состоит из двух белых яиц, насиживанием которых занимается самка[10]. Яйца колибри очень малы и могут варьировать от 11 × 8 мм у колибри-пчёлки до 20 × 12 мм у исполинского колибри. Масса яиц составляет от 0,4 г до 1,4 г. У маленьких колибри масса яиц составляет до 35 % от массы самки[30][50]. По всей видимости, именно этим объясняется тот факт, что маленькие колибри, в отличие от средних и крупных, обычно делают только одну кладку в год[50]. По другим сведениям, тропические колибри могут откладывать яйца 2—3 раза в год, а птицы, у которых сезон размножения привязан к тёплому времени года или к сезону дождей, — только раз[30]. Инкубационный период в среднем составляет 16—19 дней[50].
Птенцы
Птенцы появляются на свет полностью голыми[10][49][55], вскоре покрываются густым белым или серым пухом[55]. В течение первых нескольких недель жизни температура тела птенцов может падать до температуры окружающей среды, не причиняя им вреда. Птенцы представителей Cypseloidinae обычно покрыты пухом, что способствует терморегуляции[10]. Кормление происходит редко[55], птенцы могут оставаться без присмотра в течение длительных промежутков времени, пока взрослые тратят время на добычу относительно небольших насекомых. В плохую погоду родители могут отсутствовать часами[10]. Взрослые стрижи отрыгивают болюс с насекомыми непосредственно в горло птенцов или делят его между ними[49]. Аналогичным образом питаются древесные стрижи, птенцы которых на более поздних стадиях способны брать пищу из рта родителей[48]. Непереваренные остатки насекомых птенцы стрижей выбрасывают через рот в виде погадок[55]. Часто птенцы обладают большим количеством жира, что помогает им выжить при редком кормлении и плохой погоде, при этом вес птенцов за период голодания может уменьшаться в два раза[49].
Период гнездования у стрижей длиннее, чем у других птиц схожего размера, и не связан с размерами самих стрижей. Птенцы чёрного стрижа остаются в гнезде в среднем 42 дня, в плохую погоду — до 56 дней. Дольше всех в гнезде могут оставаться птенцы пятнистолобого американского стрижа — 65—70 дней в особенно плохую погоду. За неделю до вылета из гнезда птицы теряют вес, что помогает им достичь оптимальной нагрузки на крыло, необходимой для быстрого и манёвренного полёта[49]. У усатого древесного стрижа общий период гнездования составляет более 60 дней[48].
Слепые и беспомощные птенцы колибри имеют короткий клюв, на спине у них растёт два ряда эмбрионального пуха. В возрасте до 9 дней у них открываются глаза, появляются основные перья на крыльях, спине и хвосте. В возрасте 7—12 дней птенцы способны к самостоятельной терморегуляции и самка перестаёт греть их. К концу этого периода они достигают до 80 % массы взрослых птиц. Незадолго до вылета из гнезда птенцы становятся более активными[50]. Птенцы колибри остаются в гнезде 23—26 дней, в Андах — до 30—40 дней[50] (по другим данным, покидают гнездо на 25—35-й день[34]). Ещё около месяца после этого самка продолжает кормить птенцов. Во время кормления маленькие птенцы получают нектар и крошечных членистоногих, которые самка отрыгивает, вставляя тонкий клюв в рот каждого птенца[50].
 Птенец блестящего клехо
Птенец блестящего клехо Дымчатый иглохвост с птенцами
Дымчатый иглохвост с птенцами Самка колибри-каллиопы кормит птенцов
Самка колибри-каллиопы кормит птенцов
Продолжительность жизни
Выживаемость птенцов пальмовых стрижей крайне мала и составляет 26 %, в то время как выживаемость птенцов дымчатых иглохвостов — 96,1 %. После первого года уровень смертности стрижей очень низок. Например, в колонии бледного стрижа на Гибралтаре уровень смертности в первый год составил 67,3 %, а в последующие — 26 %. Показатели смертности чёрного стрижа, дымчатого иглохвоста и красношейного американского стрижа составляют 15—19, 15—17 и 19—29 %, соответственно[49].
Для колибри характерна высокая смертность в раннем возрасте. Успешные показатели размножения (отношение числа выживших птенцов к числу отложенных яиц) у некоторых тропических видов варьируют от 20 до 40 %, а в случае колибри — горных звёзд и шлемоносных колибри составляют 60 %. При этом чаще всего у горных звёзд потери связаны с неблагоприятными климатическими условиями и реже с разорением гнёзд хищниками. В то время как у североамериканских колибри успешные показатели в целом идентичны, достигая 60 % у трёхцветного селасфоруса, смертность в основном связана с хищниками. Данные о доле выживших в год малочисленны, у североамериканских видов этот показатель составляет от 40 до 50 %[50].
Средняя продолжительность жизни стрижей в дикой природе составляет 7—10 лет[31]. Рекордный возраст зафиксирован у белобрюхого стрижа — 26 лет[49], для чёрного стрижа известен возраст 21 год[31], для дымчатого иглохвоста — 14 лет[49]. В дикой природе максимальный возраст колибри в среднем составляет 5—8 лет, в неволе птицы живут на несколько лет дольше. Рекордные показатели принадлежат особям синегорлого сверкающего колибри, возраст которого составлял 12 лет, и рубиновогорлого колибри — 11 лет[50].
Эволюция
| Кладограмма стрижеобразных по Ksepka et al.[56] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Предполагается, что стрижеобразные — очень древняя группа, отделившаяся от праракшеобразных в позднемеловой эпохе около 70 млн лет назад. Центр происхождения стрижеобразных неясен[7]. Кости их крыла в основном короткие и крепкие. Из-за их размера, обычно немногим больше 10—15 мм, они чаще бывают нетронутыми в отложениях, но их и сложнее заметить. С конца XIX века было обнаружено довольно много остатков птиц, подобных стрижам[57].
По-видимому, древесные стрижи отделились от общего ствола стрижеобразных ещё до разделения настоящих стрижей и колибри, они сохранили ряд примитивных черт[7]. В 1984 году британский орнитолог и палеонтолог К. Харрисон описал остатки Eocypselus vincenti из нижнеэоценовых отложений Англии. Сам Харрисон отнёс вид к монотипическому семейству Eocypselidae. Ряд исследователей считает вид близким к древесным стрижам[13], другие рассматривают его в качестве базального общего предка, сохраняющего монофилию стрижей, древесных стрижей и колибри[56].
Родственным древесным стрижам считается ископаемое семейство Aegialornithidae (Lydekker, 1891)[13][12], древнейшие находки которого (роды Primapus, Cypselavus и Aegialornis) были обнаружены в Англии и Франции[12]. Окаменелые остатки самого раннего предположительного предка стрижеобразных были найдены в Англии в отложениях нижнего эоцена. Птица получила название Primapus lacki в честь британского орнитолога Дэвида Лэка, занимавшегося изучением стрижей. По всей видимости, представители рода Primapus вымерли в конце эоцена. Фрагменты ещё пяти видов птиц были обнаружены в отложениях верхнего эоцена и олигоцена во Франции, они были объединены в род Cypselavus[4]. Cypselavus gallicus считается самым ранним представителем древесных стрижей[13].
Scaniacypselus, окаменелые остатки которого из среднего эоцена были обнаружены в Дании и Германии, возможно, является самым ранним представителем настоящих стрижей[13]. К эоцену относят появление современных родов Cypseloides, Chaetura[12]. В отложениях в штате Вайоминг были обнаружены остатки стрижей, схожих с представителями Chaetura (нижний эоцен)[4], а в отложениях во Франции — Cypseloides mourerchauvirea (верхний эоцен) и Cypseloides ignotus (нижний миоцен)[4]. Другие источники относят самые ранние остатки настоящих стрижей к миоцену[10].
Учёные полагают, что колибри оформились не раньше миоцена, их предки походили на стрижей, имели бо́льшие размеры и менее длинный клюв по сравнению с современными представителями семейства. Немногие ископаемые колибри относятся к поздним эпохам, из отложений верхнего плейстоцена известно несколько современных видов[12]. Предположительно, первые колибри появились в Южной Америке, а затем распространились в Центральную и Северную Америку. Согласно предположениям Харрисона, колибри появились в верхнем мелу или нижнем кайнозое, однако их остатков, относящихся к этому периоду, не обнаружено[57]. На Северном Кавказе был описан Jungornis tesselatus из нижнего олигоцена, некоторые учёные считают его возможным предком колибри[13]. Ряд учёных объединяет роды Jungornis, Procypseloides, Scaniacypselus из эоцена и олигоцена в вымершее семейство Jungornithidae, и рассматривают его как общего предка настоящих стрижей и колибри[12].
По мнению российского учёного Александра Альфредовича Карху, современная систематика должна отражать две основные линии эволюции стрижеобразных. Таксоны Aegialornis, Primapus и Cypselavus, относящиеся к палеоцену и раннему эоцену, он поместил рядом с родом Hemiprocne в семейство древесных стрижей в подотряд Hemiprocni, а таксоны Procypseloides, Scaniacypselus и семейство Jungornithidae, относящиеся к палеоцену, поместил рядом с семейством настоящих стрижей в подотряд Apodi, отдельно разместив колибри. Последнюю группу объединяет схожая структура плечевой кости[4]. Укороченная плечевая кость у стрижей компенсируется длинными первостепенными маховыми перьями и сильно удлинённой кистью, что позволяет им делать быстрые взмахи крыла и скользить по воздуху. У колибри первостепенные маховые крылья также довольно длинные, но общая длина крыла относительно размеров тела короче, чем у стрижей, что позволяет им совершать трепещущий полёт. Таким образом, отсутствие оперения у многих остатков стрижеподобных птиц не позволяет их правильно классифицировать[13][56].
См. также
Примечания
- Коблик, 2001, с. 52.
- Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Русский язык, РУССО, 1994. — С. 150. — 2030 экз. — ISBN 5-200-00643-0.
- Hackett S. J. et al. A Phylogenomic Study of Birds Reveals Their Evolutionary History (англ.) // Science. — 2008. — 27 June (iss. 5883). — P. 1763—1768. — doi:10.1126/science.1157704.
- HBW Alive: Family Apodidae, Systematics.
- Apodiformes (swift) (англ.) информация на сайте Paleobiology Database. (Дата обращения: 2 декабря 2019)
- Жизнь животных, 1986, с. 314.
- Коблик, 2001, с. 53.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Owlet-nightjars, treeswifts, swifts (англ.). IOC World Bird List (v11.2) (15 июля 2021). doi:10.14344/IOC.ML.11.2. Дата обращения: 16 августа 2021.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Hummingbirds (англ.). IOC World Bird List (v11.1) (20 января 2021). doi:10.14344/IOC.ML.11.1. Дата обращения: 12 января 2021.
- Parkes K. C. Apodiform (англ.). Encyclopaedia Britannica. Дата обращения: 7 марта 2019. Архивировано 3 декабря 2018 года.
- Farner D. Avian Biology, Volume VIII. — Elsevier, 2012. — P. 134—135. — 280 p. — ISBN 978-1-4081-2501-4.
- Коблик, 2001, с. 54.
- Mayr G. Phylogeny of Early Tertiary Swifts and Hummingbirds (Aves: Apodiformes) (англ.) // The Auk. — Oxford University Press, 2003. — P. 145—151. — doi:10.2307/4090149. Архивировано 24 февраля 2019 года.
- Caprimulgiformes (англ.). Handbook of the Birds of the World Alive. Дата обращения: 18 февраля 2019.
- Sangster G. A name for the clade formed by owlet-nightjars, swifts and hummingbirds (Aves) (англ.) // Zootaxa. — 2005. — Iss. 799. — P. 1—6. — doi:10.11646/zootaxa.799.1.1.
- HBW Alive: Family Trochilidae.
- HBW Alive: Family Apodidae.
- HBW Alive: Family Hemiprocnidae.
- Коблик, 2001, с. 62.
- HBW Alive: Family Trochilidae, Morphological aspects.
- HBW Alive: Family Apodidae, Morphological aspects.
- HBW Alive: Family Hemiprocnidae, Morphological Aspects.
- Коблик, 2001, с. 66.
- Коблик, 2001, с. 55.
- Swifts (англ.). Encyclopaedia Britannica. Дата обращения: 7 марта 2019. Архивировано 27 июня 2018 года.
- HBW Alive: Family Hemiprocnidae, Food and feeding.
- Коблик, 2001, с. 63.
- HBW Alive: Family Apodidae, General Habits.
- HBW Alive: Family Apodidae, Voice.
- Коблик, 2001, с. 67.
- Коблик, 2001, с. 58.
- HBW Alive: Family Apodidae, Movements.
- HBW Alive: Family Hemiprocnidae, Systematics.
- Коблик, 2001, с. 68.
- HBW Alive: Family Trochilidae, Status and Conservation.
- HBW Alive: Family Trochilidae, Movements.
- Коблик, 2001, с. 69.
- HBW Alive: Family Hemiprocnidae, Movements.
- Коблик, 2001, с. 56.
- HBW Alive: Family Apodidae, Habitat.
- HBW Alive: Family Hemiprocnidae, Status and Conservation.
- HBW Alive: Family Hemiprocnidae, Habitat.
- HBW Alive: Family Hemiprocnidae, General Habits.
- HBW Alive: Family Trochilidae, Habitat.
- HBW Alive: Family Apodidae, Food and Feeding.
- HBW Alive: Family Trochilidae, Food and feeding.
- Коблик, 2001, с. 65.
- HBW Alive: Family Hemiprocnidae, Breeding.
- HBW Alive: Family Apodidae, Breeding.
- HBW Alive: Family Trochilidae, Breeding.
- HBW Alive: Family Apodidae, Relationship with Man.
- Marin, Stiles, 1992, p. 316.
- HBW Alive: Family Trochilidae, General habits.
- Lack D. A review of the genera and nesting havits of swifts // Auk. — 1956. — Vol. 73. — P. 13—32.
- Коблик, 2001, с. 57.
- Ksepka D. T., Clarke J. A., Nesbitt S. J., Kulp F. B., Grande L. Fossil evidence of wing shape in a stem relative of swifts and hummingbirds (Aves, Pan-Apodiformes) (англ.) // Proceedings: Biological Sciences. — Royal Society, 2013. — Vol. 280, no. 1761. — P. 1—8. — doi:10.1098/rspb.2013.0580. Архивировано 29 марта 2019 года.
- Harrison C. J. O. A revision of the fossil swifts Vertebrata, Aves, Suborder, Apodi),with descriptions of three new genera and two new species (англ.) // Mededelingen van de werkgroep voor tertiaire en kwartaire geologie. — 1984. — Vol. 21, iss. 4. — P. 157—177.
Литература
- Гладков Н. А., Иноземцев А. А., Михеев А. В., Дроздов Н. Н., Ильичев В. Д., Константинов В. М., Курочкин Е. Н., Потапов Р. Л. Жизнь животных. Том 6. Птицы / под ред. В. Д. Ильичева, А. В. Михеева, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1986. — Т. 6. — 527 с.
- Коблик Е. А. Отряд стрижеобразные — Apodiformes // Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). — М.: Издательство МГУ, 2001. — Т. 3. — С. 38—40, С. 52—70. — 360 с. — 400 экз. — ISBN 5-211-04072-4.
- Chantler P. Family Apodidae (Swifts) (англ.). Handbook of the Birds of the World Alive (10 сентября 2014). Дата обращения: 20 января 2019.
- Marin M., Stiles F. G. On the biology of five species of swifts (Apodidae, Cypseloidinae) in Costa Rica // Proceeding of the Western forundation of vertebrate zoology. — 1992. — Vol. 4. — P. 287—351.
- Parkes K. C. Apodiform (англ.). Encyclopaedia Britannica. Дата обращения: 7 марта 2019.
- Schuchmann K. L., Bonan A. Family Trochilidae (Hummingbirds) (англ.). Handbook of the Birds of the World Alive (10 сентября 2014). Дата обращения: 25 декабря 2018.
- Wells D. Family Hemiprocnidae (Treeswifts) (англ.). Handbook of the Birds of the World Alive (10 сентября 2014). Дата обращения: 20 февраля 2019.