Рубиновогорлый колибри
Рубинового́рлый коли́бри[1][2][3][4][5], или рубиного́рлый коли́бри[6], или краснозо́бый коли́бри[7][5], или обыкнове́нный колибри[5], или обыкнове́нный (рубинового́рлый, краснозо́бый) архило́хус[5] (лат. Archilochus colubris), — вид птиц семейства колибри, названный так из-за ярко-красного воротника самцов. Остальное оперение — изумрудно-зелёное с металлическим блеском сверху, светлое снизу. Рубиновогорлый колибри — единственный вид колибри, который гнездится в восточных районах Северной Америки. На зиму птицы улетают в Мексику и Центральную Америку, двигаясь вдоль побережья Мексиканского залива либо непосредственно через залив. Питаются нектаром цветковых растений и мелкими насекомыми, весной могут пить сок деревьев из отверстий в стволах, которые пробивает желтобрюхий дятел-сосун. Заботой о потомстве занимается исключительно самка: она строит гнездо из белого пуха растений, маскирует его снаружи лишайником и откладывает два белых гладких яйца, которые насиживает 12—14 дней. Вылупившихся птенцов самка обогревает ещё 9 дней, покидая их только для приёма пищи, а позже прилетает только для того, чтобы покормить. Через 18—22 дня птенцы покидают гнездо.
| Рубиновогорлый колибри | ||||||||
|---|---|---|---|---|---|---|---|---|
| ||||||||
| Научная классификация | ||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Подкласс: Инфракласс: Клада: Отряд: Подотряд: Колибри (Trochili) Семейство: Подсемейство: Вид: Рубиновогорлый колибри |
||||||||
| Международное научное название | ||||||||
|
Archilochus colubris (Linnaeus, 1758) |
||||||||
| Синонимы | ||||||||
|
||||||||
| Ареал | ||||||||
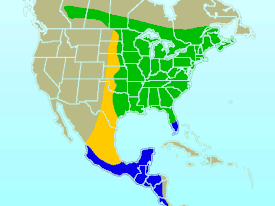 Гнездовой ареал Места зимовки Путь миграции |
||||||||
| Охранный статус | ||||||||
| ||||||||



Рубиновогорлый колибри был включён Карлом Линнеем в десятое издание «Системы природы» в 1758 году, но ещё раньше был описан в работах Марка Кейтсби и Джорджа Эдвардса. Международный союз орнитологов относит вид к роду колибри-архилохусов, включающему также черногорлого архилохуса, обитающего в западных районах Северной Америки. Международный союз охраны природы присвоил рубиновогорлому колибри статус вида, вызывающего наименьшие опасения. В силу энергозатратного территориального и брачного поведения взрослых самцов намного меньше, чем самок. За последние 50 лет численность рубиновогорлого колибри увеличилась в два раза.
Описание
Общие сведения

.jpg.webp)
Рубиновогорлый колибри — маленький представитель семейства, длина его тела 76—89 мм, средняя масса 3,0 г (2,3—5,15 г) у самцов и 3,5 г (2,56—6,09 г) у самок[8]. Размеры птиц не отличаются географически. Сравнение массы особей в Алабаме, через которую пролетают птицы из разных регионов во время миграции, не показало каких-либо существенных различий в зависимости от региона, в котором птицы появились на свет[9][10]. Вместе с тем самки рубиновогорлого колибри крупнее самцов: отношение длины крыла самца и самки в среднем составляет 0,87, а массы тела — 0,88 (для измерений, проведённых в мае, когда у птиц почти отсутствуют связанные с миграцией или размножением излишки массы). Различия в размерах между самцами и самками рубиновогорлого колибри одни из наибольших среди всех видов колибри[11].
За яркое оперение на шее этот вид колибри называют «рубиновогорлым» или «краснозобым» (англ. ruby-throat)[10][12]. Самца рубиновогорлого колибри отличает сверкающий рубиново-красный воротник, светлое сероватое оперение груди, зеленоватое по бокам, и блестящее изумрудно-зелёное оперение сзади. Голова зелёная с белым пятном за глазом. У самок отсутствует яркий воротник, а оперение спереди светло-коричневое. Оперение молодых птиц схоже с оперением самок, у молодых самцов со временем на горле появляется всё больше красных перьев[13]. Крыло рубиновогорлого колибри несёт 10 первостепенных маховых перьев и 6 второстепенных, хвост — 10 рулевых перьев. На основании данных из Алабамы, Пенсильвании и Мичигана длина крыла взрослого самца составляет 33,29—43,1 мм, самки — 38,05—47,25 мм; хвоста — 23,6—32,1 мм и 21,1—31 мм соответственно[8]. В сложенном состоянии крылья рубиновогорлого колибри не выступают за хвост[13].
Половой диморфизм выражен не только в окраске оперения, но и в форме клюва[8]. При изучении трёх основных признаков морфологии клюва индекс кривизны надклювья (англ. maxillary curvature index) у самцов и самок совпадал, а индекс кривизны подклювья (англ. mandibular curvature index) и круговая кривизна клюва (англ. circle-curvature index) — отличались. На основании более полного исследования морфологии клюва, включившего 35 различных признаков, учёные пришли к выводу, что у рубиновогорлого колибри длинный и изогнутый клюв самок сильно отличается от короткого и более прямого клюва самцов (данные отличаются от исследования индексов в силу того, что кривизна клюва непостоянна)[14]. Длина клюва взрослых самцов составляет 14,24—19,8 мм, самок — 15,62—20,8 мм[8], в среднем она слегка превышает размеры головы[13]. Различия в морфологии клюва между самцами и самками встречаются также у Eulampis jugularis (у этого тропического колибри половой диморфизм выражен ещё сильнее), разноклювой гуйи (Heteralocha acutirostris), зелёного лесного удода (Phoeniculus purpureus) и других птиц. В случае рубиновогорлого колибри, у которого в районах гнездового ареала отсутствуют конкуренты, различия в форме клюва самцов и самок позволили занять более широкую экологическую нишу[14]. У молодых особей на клюве присутствуют складки, которые со временем исчезают, что позволяет определять возраст[8][15].
Окраска

Сверху оперение самцов рубиновогорлого колибри металлического бронзово-зелёного цвета, в который также окрашены два центральных рулевых пера, остальные перья хвоста тёмно-бронзового цвета с фиолетовым отливом. Маховые перья тёмно-коричневые, с фиолетовым оттенком. Горло самцов покрывает ярко-красный металлический воротник (более золотистый или зеленоватый по бокам горла), над которым располагается бархатная чёрная маска, покрывающая подбородок, скулы, уши и область под глазами. Непосредственно за глазом расположено маленькое белое пятно[8]. Оперение самцов в верхней части груди очень светлое, коричневато-белое, более тёмное коричневато-серое в нижней части груди и на брюхе, ещё более тёмное по бокам, с металлическим бронзово-зелёным оттенком[8][13]. Подхвостье белое. У взрослых самцов глубина разреза на хвосте может достигать 6—9 мм[8].
Оперение самок металлического бронзово-зелёного, золотисто-зелёного или зеленовато-бронзового цвета сверху, в такой же цвет окрашены два центральных рулевых пера[8]. На трёх внешних рулевых перьях самок хорошо заметны широкие белые кончики, которые отсутствуют у самцов[8][13], остальная часть внешних рулевых перьев бронзово-зелёная с внешней стороны и чёрная — с внутренней. Маховые перья тёмно-коричневые, с фиолетовым оттенком. Маленькое пятно за глазом не такое белое, как у самцов. Маска у самок тёмно-серая, а горло обычно тусклое, серовато-белое, лишь у старых самок может развиться несколько красных перьев в центре горла. Оперение снизу серовато-белое или коричневато-белое, более светлое в районе горла[8].
Взрослые птицы линяют дважды в год, соответственно этому различают два сезона, хотя внешний вид оперения меняется мало. В период с июля по ноябрь у некоторых самцов воротник может быть неполным; кроме того, могут отмечаться ювенильные рулевые перья с белыми кончиками[8].
Пуховой наряд только что вылупившихся птенцов состоит из сероватого пуха и 12 пар нитевидных перьев на спине длиной 4 мм от охристо-жёлтого до желтовато-коричневого цвета. Его можно наблюдать преимущественно с апреля по июнь[8]. Ювенильное оперение самцов с мая по август похоже на оперение взрослых самок, но кончики свежих перьев спины окрашены в цвета от сероватого до светло-коричневого; на кончиках рулевых перьев небольшие белые пятна, а на горле довольно много тёмных, но не красных прожилок и пятен. У самок оперение в этом возрасте отличается более крупными белыми пятнами на кончиках рулевых перьев, более широкими первостепенными маховыми перьями и отсутствием либо меньшим количеством тёмных пятен и прожилок на горле. Среди учёных нет согласия в том, как определять пол птиц в этом возрасте. Некоторые исследователи используют раскраску горла, другие предпочитают измерять размер белых пятен на рулевых перьях, длину и остроту маховых перьев или глубину разреза на хвосте. С точностью 95 % пол птиц в этом возрасте определяется по длине крыла: у самцов она меньше 43 мм, у самок — больше или равна[8].

Оперение молодых птиц с июля по октябрь отличается более зелёными перьями спины, без сероватых кончиков, появлением красных перьев на горле у самцов. У некоторых самцов воротник в это время почти полный, но у большинства птиц сформирован лишь частично. Оперение колибри-первогодок с октября по июль очень слабо отличается от предыдущего, иногда у самцов на этой стадии сохраняется ювенильное оперение с более тусклым и не до конца сформированным воротником. Самки могут сохранить рулевые перья ювенильного оперения. Внешние первостепенные перья самок и молодых птиц шире, чем у взрослых самцов, и менее заострены на концах. Взрослые самцы испытывают наибольшую нагрузку на крылья из-за звуков, производимых ими во время ухаживания[8].
Географические вариации оперения и стратегий линьки незначительны. Отмечается, что многие самцы, прилетающие в октябре в Гватемалу, имеют оранжевый, а не красный воротник. В других регионах оранжевый оттенок воротника встречается у самцов с конца лета и до зимы. Изменение цвета может быть связано как с возрастом перьев, так и с химическими изменениями их меланиновой структуры. Возможно, как и в случае охристого колибри (Selasphorus rufus), некоторые самцы не меняют перья воротника во время весенней линьки[8].
Рубиновогорлый колибри перья тела чистит клювом, а перья головы и шеи — лапами. Во время потягивания расправляет крылья вниз вдоль тела, может при этом распушать хвост. Купается в собирающейся на листьях воде, солнечные ванны принимает на широких тёмных поверхностях, лёжа на животе и расправив крылья[16].
Линька
.jpg.webp)
Линька рубиновогорлого колибри, как и других североамериканских мигрирующих видов колибри, обычно очень сложна: исследователи рассматривают стратегии линьки по-разному и дают им разные названия[8].
Первая летняя линька молодых птиц приводит лишь к частичной смене перьев воротника у самцов (крайне редко к сентябрю — октябрю формируется полностью окрашенный воротник) и контурных перьев у самцов и самок. Эта линька начинается в местах гнездования, но может прерываться на миграцию и продолжаться на зимних квартирах, её полную продолжительность учёные оценивают с июля по ноябрь. Смена перьев воротника может продолжаться во время первой линьки маховых перьев либо прерываться на этот период[8].
Предбрачная линька проходит в период с октября по апрель в местах зимовки, при этом первая зимняя линька обычно начинается на один месяц позже, чем последующие. Продолжительность линьки отдельных особей меньше. В это время может проходить полная или частичная смена перьев[8]. Первым меняется внутреннее первостепенное маховое перо, за ним последовательно остальные, и только последнее — внешнее — маховое перо, как и у большинства колибри, меняется раньше предпоследнего (p1—p8, p10, p9). Учёные объясняют это тем, что во время смены самого длинного девятого махового пера только что сменившееся десятое перо максимально сильное, что сохраняет способность птиц к маневренному полёту[8][15]. После замены шестого первостепенного махового пера начинается смена второстепенных маховых и рулевых перьев. Смена второстепенных маховых перьев обычно осуществляется попеременно (s1, s6, s2, s5, s3, s4), а рулевых — от внутренних к внешним (r1—r5)[8]. Согласно другим исследованиям, смена рулевых перьев начинается лишь после полной замены маховых перьев[15]. В целом сначала осуществляется смена маховых и рулевых перьев, а за ними довольно быстро в конце февраля — начале апреля происходит смена контурных перьев. В последнюю очередь происходит смена перьев воротника. Внешние рулевые перья и некоторые перья воротника в первую линьку могут оставаться от ювенильного оперения[8].
Частичная послебрачная линька проходит в местах гнездования и во время осенней миграции с июля по сентябрь; возможно, она продолжается в октябре — ноябре после перелёта птиц к местам зимовки. Смена контурных перьев начинается с перьев головы. Определить стадию линьки по внешним проявлениям почти невозможно, результаты некорректны в половине случаев[8]. Некоторые учёные относят описанную выше смену полётных перьев к послебрачной, а не к предбрачной линьке[8].
Схожие виды
.jpg.webp)
Родственный черногорлый архилохус (Archilochus alexandri) немного меньше рубиновогорлого колибри, его отличает фиолетовый воротник, более короткие крылья и непокрытые ноздри[8]. Молодых птиц и самок этих двух видов различать заметно сложнее: у рубиновогорлого колибри голова сзади и спина заметно ярче и зеленее, оперение по бокам и подхвостье имеют светло-коричневый оттенок, маска на лице темнее, а белое пятно за глазом выделяется сильнее. В то же время у черногорлого архилохуса голова обычно серовато-зелёная, боковые перья — серовато-оранжевые[8][13], а рулевые — голубовато-зелёные[13]. Кончики внешних первостепенных маховых перьев у рубиновогорлого колибри узкие и заострённые, в то время как у черногорлого архилохуса — более широкие и тупые[8][13]. Кроме того, клюв рубиновогорлого колибри короче и толще, чем вытянутый и искривлённый клюв черногорлого архилохуса, а различия в морфологии клюва, связанные с половым диморфизмом, более заметны[14].
У представителей рода колибри-архилохусов (Archilochus), который включает два вида, в отличие от родственных колибри-селасфорусов (Selasphorus) и калипт (Calypte), внутренние первостепенные маховые перья (p1—p6) заметно у́же, чем внешние (p7—p10)[8]. Воротник трёхцветного селасфоруса (Selasphorus platycercus) розово-пурпурный[8][13], кроме того, у этого вида длиннее хвост[13]. Сама форма воротника отличает рубиновогорлого колибри, у которого в красный цвет окрашено всё горло, от представителей рода калипт (Calypte), у которых цветовое пятно не заходит на стороны[8]. У самки калипты Коста (Calypte costae) более короткий хвост и отсутствует черноватый отлив на тусклом серо-зелёном оперении. Самки калипты Анны (Calypte anna) в целом крупнее самок рубиновогорлого колибри, они имеют другую форму кончиков первостепенных перьев и более тусклое оперение[8][13].
Вокализация
Вокализация рубиновогорлого колибри изучена слабо; в отличие от черногорлого архилохуса, сонографическому анализу подверглись только звуки, издаваемые при агрессивных столкновениях. По акустической структуре, длине и синтаксису звукового сигнала вокализации рубиновогорлого колибри и черногорлого архилохуса схожи. Вокализация самки отличается более плавными и менее хриплыми позывками по сравнению с самцом. Область переднего мозга, отвечающая за пение, находится в зачаточном состоянии у самцов рубиновогорлого колибри и совсем не развита у самок[17].
В репертуаре рубиновогорлого колибри самыми сложными являются агрессивные сигналы. Основной позывкой при столкновении птиц является быстрый скрипучий крик, состоящий из пяти различных типов звуков, которые повторяются для создания интенсивности. Отдельно выделяется шестой, победный, сигнал. Не связанные со столкновением звуковые сигналы в полёте представляют собой простую цепочку звуков. В качестве территориальных позывок выступают серии монотонных звуковых сигналов. Сразу после совокупления самцы издают серию взволнованных сигналов и высоких быстрых резких криков. По окончании сезона размножения резкие позывки рубиновогорлого колибри сменяются на более спокойные. Самки у гнезда и перед совокуплением издают звуки «mew», а во время кормления — приглушённые сигналы «tic-tic». Во время осенней миграции и самцы, и самки издают повторяющийся сигнал «chee-dit». В это время птицы могут собираться в большие группы и реже демонстрируют агрессию. В Коста-Рике рубиновогорлые колибри довольно тихие, от птиц можно услышать мяукающие сигналы «tew» или «tyew»[17].
В течение дня вокализация рубиновогорлого колибри ограничивается агрессивными взаимодействиями. Территориальная позывка исполняется на рассвете, для неё самцы используют более высокие и открытые насесты. Учёные полагают, что стандартные песенные насесты отсутствуют, хотя самцы предпочитают находиться около центра своей территории[17].
Взрослые особи обоих полов производят звуки и крыльями. Во время первой линьки первостепенные маховые перья самцов становятся короткими, узкими и заострёнными, что позволяет им создавать более высокие жужжащие звуки переменной тональности. Самцы используют их во время агрессивных столкновений или ухаживаний. Самец может менять звук, издаваемый крыльями: внутренние первостепенные маховые перья с угловатыми кончиками создают громкое жужжание во время «челночных» движений брачного представления, а заострённые внешние — резкий трескучий звук в нижней части «нырков» (черногорлый архилохус в это время издаёт слабую трель). У самок первостепенные маховые крылья более длинные, широкие и округлые, они производят низкий, мягкий и тихий гул. Частота звука, издаваемого крыльями, составляет около 70 Гц у самцов и 52 Гц у самок[17].
Распространение
Ареал
Рубиновогорлый колибри обитает на востоке Северной Америки[18] и на большей части своего ареала является единственным гнездящимся видом колибри[14][19]. Лишь в западных районах Техаса и округах Команче и Стивенс в Оклахоме ареал пересекается с родственным черногорлым архилохусом, а в центральных и южных районах Техаса — с юкатанской амазилией (Amazilia yucatanensis)[19]. Непосредственный ареал рубиновогорлого колибри (англ. extent of occurence) составляет 3 350 000 км². Международный союз охраны природы относит к нему в первую очередь территории в Канаде и США, а также территории Багамских Островов, Бермудских Островов, Каймановых Островов, Кубы, Доминиканской Республики, Сальвадора, Гватемалы, Гаити, Гондураса, Мексики, Никарагуа, Панамы, Пуэрто-Рико, Теркс и Кайкос. Кроме того, птиц отмечали в Белизе, Коста-Рике, Никарагуа и Сен-Пьер и Микелон[20].
Гнездовой ареал вида простирается от центральных и южных районов Альберты и Саскачевана через южные районы Манитобы и крайний северо-восток Монтаны и Северной Дакоты до центральных районов Онтарио и южных районов Квебека, Нью-Брансуика, Острова Принца Эдуарда, Новой Шотландии и далее на юг до Великих равнин, восточных и южных районов Техаса и центральных районов Флориды (крайне редко птицы гнездятся ещё южнее), включая побережье Мексиканского залива. Рубиновогорлые колибри широко распространены в Приморских провинциях Канады. Гнёзда отмечали в долине Кодрой на острове Ньюфаундленд. В Квебеке ареал колибри простирается до 49° с. ш., вместе с тем гнездящихся птиц не отмечали на островах Антикости и Мадлен в заливе Святого Лаврентия. В Онтарио рубиновогорлые колибри в основном сконцентрированы около озёр в южной части провинции, а при продвижении на север до 50° с. ш. гнёзда встречаются редко. В районе Великих равнин граница гнездового ареала совпадает с границей леса и достигает восточных районов Южной Дакоты и Небраски, центральных и южных районов Канзаса, центральных районов Оклахомы и побережья Техаса. Западная граница ареала в центральных и южных регионах США проходит по 97—98° з. д. На севере ареала птицы распространяются дальше на запад, включая округ Киттитас в штате Вашингтон, север Альберты и северо-восток Британской Колумбии (возможно, они представляют собой неизвестную ранее часть популяции)[18]. Ввиду особенностей рациона рубиновогорлого колибри северная граница его ареала коррелирует с распространением желтобрюхого дятла-сосуна (Sphyrapicus varius) и проделанных им отверстий в деревьях. Именно с увеличением численности последних учёные связывают распространение гнездового ареала рубиновогорлого колибри на северо-восток, хотя оно затронуло и другие экорегионы, в которых дятлы-сосуны не встречаются[18][21].
На зиму птицы мигрируют в Мексику, Белиз, Гондурас, Гватемалу, Сальвадор, Никарагуа, Коста-Рику и Панаму. В Мексике они встречаются как на тихоокеанских склонах на юге штата Синалоа, так и во внутренних регионах в штате Оахака, а также на атлантических склонах на юге штата Веракрус. Птицы зимуют на полуострове Юкатан, включая острова Косумель и Хольбош. В Коста-Рике рубиновогорлые колибри встречаются на тихоокеанском побережье, в основном на северных склонах гор, а в Панаме — в самых западных регионах страны на тихоокеанском побережье, куда они изредка долетают. Рубиновогорлые колибри не встречаются на атлантическом побережье Гондураса и Никарагуа[18]. Ранее рубиновогорлые колибри крайне редко зимовали на юго-востоке атлантического побережья США и на севере Флориды, в последнее время это происходит чаще. В Луизиане рубиновогорлые колибри, окольцованные в летние месяцы, не были отловлены повторно в зимние, а птицы, окольцованные в зимние месяцы, не были отловлены повторно в летние. Чтобы определить, есть ли птицы, которые обитают в этом регионе круглый год, требуются дополнительные исследования. Распространение рубиновогорлого колибри в зимнее время на атлантическое побережье и северные районы Мексиканского залива может быть связано с мягкими зимами, большим количеством искусственных кормушек и популярностью экзотических растений среди населения[18].
В Мексике птиц отмечали на высоте до 3000 метров над уровнем моря, в Сальвадоре — до 1066 метров, в Гондурасе — до 1500 метров[18]. Международный союз охраны природы в качестве верхней отметки называет высоту 1900 метров над уровнем моря[20].
Среда обитания
Обширный ареал рубиновогорлого колибри включает разнообразные места обитания[21]. Рубиновогорлый колибри встречается в смешанных или лиственных лесах, на опушках леса или в садах. На юго-востоке США может устраивать гнёзда в деревьях рода сосна (Pinus), в национальном парке Фанди в канадской провинции Нью-Брансуик встречается в зрелых лесах с преобладанием клёна сахарного (Acer saccharum), берёзы аллеганской (Betula alleghaniensis), ели красной (Picea rubens). На западе Канады заселяет таёжные леса. В Луизиане птицы гнездятся в зрелых лесах с преобладанием ниссы лесной (Nyssa sylvatica) и таксодиума двурядного (Taxodium distichum)[22].
Во время миграции рубиновогорлый колибри предпочитает останавливаться в местах, схожих с гнездовыми. На побережье Мексиканского залива во время весенней миграции питается на жимолости японской (Lonicera japonica), охотится на насекомых на дубе виргинском (Quercus virginiana) и Celtis tenuifolia. Во время осенней миграции добывает пищу на кампсисе укореняющемся (Campsis radicans)[22].
В местах зимовки птицы селятся в тропических лиственных лесах. В штате Халиско в Мексике предпочитают балки, в штате Чьяпас — высокие тропические засушливые леса. В центральных районах Белиза встречаются в цитрусовых рощах, в Гуанакасте в Коста-Рике предпочитают вторичные кустарники или опушки леса, намного реже встречаются в лиственных или вечнозелёных лесах, а также по краям болот. В Центральной долине в Коста-Рике птиц отмечали на заросших пастбищах, вдоль изгородей и в высоком подлеске. На юго-востоке Луизианы зимующие птицы предпочитают сады в пригородах с дубом виргинским, а также болота с преобладанием растений родов нисса (Nyssa) и секвойя (Taxodium)[22].
Миграция
.jpg.webp)
Рубиновогорлый колибри во время миграции преодолевает одно из самых больших расстояний среди всех видов колибри[21]. На зиму птицы перелетают в Центральную Америку или на побережье Мексиканского залива, при этом особи, которые гнездятся на побережье, перелетают дальше на юг, а на зиму в эти места перелетают птицы из других регионов[23]. Для рубиновогорлого колибри характерна цепная миграция первого типа, когда миграция начинается с южных регионов и распространяется на север. В Алабаме, через которую проходят многие маршруты миграции рубиновогорлого колибри, на основе данных об изотопном составе водорода в перьях птиц первого года и карты осадков Северной Америки учёные определяют широту, на которой птицы появились на свет. Было показано, что на побережье Алабамы сначала прилетают птицы с южных широт, а позднее — с северных[23][10]. Схожий тип миграции демонстрируют некоторые древесницевые (Parulidae), а также полосатый ястреб (Accipiter striatus)[10].
В Пенсильвании миграцию начинают самцы, во время весенней миграции за ними следуют самки и молодые птицы вместе, а во время осенней — сначала самки, а затем молодые птицы обоих полов. В Алабаме во время осенней миграции не было отмечено какой-либо связи с возрастом или полом, в то же время весной взрослые птицы мигрируют в более компактный временной промежуток и обычно прибывают ранее молодых[23].
Пик весенней миграции не совпадает с пиком цветения каких-либо растений, но, возможно, связан с миграцией желтобрюхого дятла-сосуна. В Алабаму первые самцы прибывают в конце февраля, самки через 8—10 дней после самцов. На юго-западе Луизианы пик миграции приходится на вторую половину апреля, самцы летят примерно на три дня раньше самок. К концу апреля самцы прибывают на территорию заповедника Паудермилл (Powdermill Nature Reserve) в Пенсильвании, самки отстают от них на две недели. В Вермонте первые птицы (предположительно самцы) появляются в первую неделю мая, в южных районах Огайо — 25 апреля — 2 мая, в северных, близ озера Эри, — 5—12 мая. К середине мая птицы обоих полов достигают Нью-Брансуика в Канаде. Вместе с тем в середине марта рубиновогорлые колибри всё ещё часто встречаются в национальном парке Пало-Верде в Коста-Рике. Во время весенней миграции с марта по май птиц отмечали на севере Багамских Островов, на Кубе, на Каймановых и Бермудских Островах. Путешествующие вдоль береговой линии Мексиканского залива рубиновогорлые колибри остаются на берегу менее одного дня, прежде чем направиться вглубь континента: на юго-западе Луизианы лишь 2 % птиц были пойманы повторно с большим интервалом. По завершении миграции физическое состояние у самок намного лучше, чем у самцов[23]. На сроки весенней миграции, возможно, оказывает влияние изменение климата[23][12]. Согласно наблюдениям, в 1970—2002 годы в Массачусетсе птицы стали прилетать раньше; за 123 года наблюдений заметно сместились отметки первых прибывших рубиновогорлых колибри на востоке штата Нью-Йорк[23]. Более раннее прибытие было отмечено также в штатах Мэн и Южная Дакота[12]. Джейсон Куртер (Jason R. Courter) с соавторами отмечал, что в 2001—2010 годы волна миграции распространялась 38 дней между 33-й и 45-й широтами (по данным прилёта первой особи), преодолевая 32,1 км в день; в 1880—1969 годы, согласно оцифрованным исследователями данным, эти показатели составляли 33,8 дня и 36,2 км/день соответственно. Более тёплые февральские температуры в местах зимовки приводят к тому, что рубиновогорлые колибри прилетают в места гнездования раньше на всех исследуемых широтах. Вместе с тем на низких широтах (до 40° с. ш.) птицы после тёплой зимы в местах гнездования прилетали раньше, а на высоких широтах, наоборот, после тёплой зимы в местах гнездования птицы прилетали позднее. Возможно, это связано с жизненным циклом растений, которым требуются холодные зимние температуры, чтобы начать цветение. В таком случае птицы дольше остаются в местах зимовок и больше питаются по дороге, чтобы собрать достаточно энергетических ресурсов. Более ранние прилёты могут быть также связаны с увеличением численности вида[12].
.jpg.webp)
Осенняя миграция в северных широтах синхронизирована с пиком цветения Impatiens capensis и лобелии пурпурной (Lobelia cardinalis), которые, по-видимому, являются важным источником нектара в этот сезон[23][10]. По данным исследований в Пенсильвании и Онтарио, на миграцию оказывают влияние направление и скорость ветра: больше всего птиц отмечалось после прохождения холодного фронта с преобладающим северо-западным ветром. Птицы часто летят в середине дня, используя утренние часы для восстановления энергии. Вдоль озера Эри в Онтарио рубиновогорлые колибри летят со скоростью 26—63 км/ч; около 33 % птиц наблюдали до 10 часов утра, при этом была отмечена корреляция с северо-западным или западным ветром. На юго-востоке штата Нью-Йорк около 60 % птиц были отмечены в промежутке с 9 до 13 часов, с пиком наблюдений при лёгком или умеренном северном или западном ветре. В южных широтах связь между миграцией и цветением Impatiens capensis отсутствует, в начале миграции важную роль играет цветение кампсиса укореняющегося и Malvaviscus arboreus var. drumondii. Кроме того, в более равнинных южных широтах перемещения не связаны с погодными условиями. Мигрирующие птицы покидают места ночёвок в 8—9 утра в южной Луизиане, а в северной Алабаме птицы редко задерживаются более одного дня и покидают территорию в 16:00—17:30 (выводы сделаны для птиц с повышенной массой 4,7—6,0 г). При этом птицы медленно и тяжело поднимаются почти вертикально вверх до тех пор, пока не скрываются из вида. На побережье Алабамы рубиновогорлые колибри прибывают с массой, превышающей безжировую массу тела на 18—90 % (в среднем около 30 %). Состояние взрослых птиц лучше, чем молодых особей, а самцов — лучше, чем самок. Птицы могут оставаться на берегу от нескольких часов до 15 дней в зависимости от того, сколько энергетических ресурсов им необходимо пополнить[23]. Птицы около 75 % времени проводят на насесте, интенсивно питаясь лишь непосредственно после рассвета и перед закатом[16]. Обычно побережье Алабамы птицы покидают до полудня. Во время осенней миграции на крайнем юге Онтарио рубиновогорлого колибри крайне редко отмечают вплоть до середины августа, после чего начинается миграция с двумя пиками — 5 и 15 сентября, что, скорее всего, связано с перемещением самцов и самок, — и продолжается до 11 октября. К 9 сентября заповедник Паудермилл на западе Пенсильвании покидают все самцы, в то время как самки и молодые птицы остаются до 3 октября. В Огайо осенняя миграция самцов начинается в конце июля, а её пик приходится на середину августа, в то время как миграция самок и молодых птиц продолжается с 10 августа по 7 сентября. На севере Алабамы осенняя миграция в основном приходится на 15 августа — 1 сентября, а на побережье штата — 7 сентября — 12 октября. Во время осенней миграции в редких случаях птиц отмечали в Калифорнии, при этом в регионе Транс-Пекос на западе Техаса птицы встречаются чаще, чем предполагалось ранее. В северные районы полуострова Юкатан первые птицы прибывают в начале сентября, пик миграции приходится на 31 октября — 3 ноября. В Коста-Рику рубиновогорлые колибри прибывают в конце сентября — начале октября. Крайне редко в промежуток с октября по февраль птиц отмечали на севере Багамских Островов, на Кубе, на Каймановых Островах[23].
Основными маршрутами миграции являются перемещение вдоль побережья Мексиканского залива или его пересечение. Птицы из Алабамы в дневные часы предпочитают передвигаться вдоль побережья, при этом на путь до северных районов полуострова Юкатан они тратят более месяца. Повторная ловля птиц на орнитологических станциях подтверждает юго-западное — северо-восточное направление миграции, хотя такая статистика может быть обусловлена расположением самих станций. Большое количество птиц наблюдается над водами залива поблизости от берега во время весенней миграции. Также много рубиновогорлых колибри отмечено во время весенней и осенней миграций на побережье Мексиканского залива в Техасе. Несмотря на достаточное количество энергетических запасов, лишь некоторые особи совершают перелёт непосредственно через Мексиканский залив[23]. В некоторых народных рассказах американских индейцев утверждается, что рубиновогорлые колибри могут путешествовать на канадских казарках (Branta canadensis), зарываясь в их перья[24].
На основании 1224 окольцованных птиц в местах гнездования и 130 повторных поимок был сделан вывод, что молодые самцы возвращаются на предыдущее место в 4,7 % случаев (23 раза), взрослые самцы — в 10,5 % (16), для самок эти показатели составляют 15,5 % (57) и 16,2 % (34) соответственно. Для птиц, зимующих на севере Флориды и на юге Алабамы, показатели возвращения в те же районы зимовок в 1998—2008 годы составили 3,6 % для самцов и 7,4 % для самок, всего повторно было отловлено 17 из 327 окольцованных птиц. На побережье Южной Каролины в 2008—2012 годы эти показатели составили 14,6 % и 31,6 % соответственно. На юге Луизианы в 1999—2012 годы показатель варьировал от 1,6 до 21 %, в среднем составляя 9 %[16].
Численность и охранный статус
Международный союз охраны природы относит рубиновогорлого колибри к видам, вызывающим наименьшие опасения (LC), вид внесён во второе приложение CITES (Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения)[20].
Общая численность рубиновогорлого колибри в США и Канаде оценивается в 34 миллиона особей[25][26]. Такие данные были получены на основании проекта Breeding bird survey в 2005—2014 годы. При этом в Огайо в 2007—2011 годы обитало около 300 тысяч особей, в Пенсильвании в 2004—2009 годы — 250 тысяч, в Онтарио в 2001—2005 годы — 500 тысяч. В 1940—1990-е годы было отмечено сокращение популяции птиц на юге Аппалачей. В 1970—1980-е годы о сокращении численности сообщалось повсеместно, однако эта информация не была подтверждена документальными свидетельствами. Вместе с тем данные Breeding Bird Survey в 1966—2015 годы показывают, что численность птиц увеличивается в среднем на 1,44 % в год, в некоторых регионах прирост превышает 3 % в год. При этом не было отмечено ни одного региона с заметным снижением численности. За 45-летний период (1970—2014) численность птиц, согласно оценкам этого проекта, возросла на 110 %[26].
Помимо проекта Breeding Bird Survey, информацию о численности птиц предоставляет атлас гнездящихся птиц, выпускаемый отдельно в разных регионах. Первое издание атласа увидело свет в 1980-е, второе — в 2000-е годы. По данным этого атласа в Мэриленде в 2002—2007 годы стали отмечать на 10 % больше птиц по сравнению с 1983—1987 годами. Аналогично в Пенсильвании в 2004—2008 годы птиц на 11 % больше по сравнению с первым изданием атласа. В Вермонте увеличение численности между 1976—1981 и 2003—2007 годами составило 2 %, а в штате Нью-Йорк между 1980—1985 и 2002—2005 годами — 21,5 %[26].

Как и остальные мигрирующие птицы, рубиновогорлые колибри сталкиваются с окнами, вышками и транспортными средствами. Объём этих столкновений остаётся неясным из-за сложностей в их обнаружении, однако учёные полагают, что такие несчастные случаи являются важной причиной смертности[25][26]. Также неясными остаются долгосрочные последствия исчезновения лесов в местах размножения и зимовки. Известно, что в районах использования гербицидов в Новой Шотландии не было отмечено ни одного рубиновогорлого колибри, вместе с тем прямое воздействие пестицидов на рубиновогорлого колибри не изучено[25]. Некоторые риски связаны с колючими растениями, в частности завезённым в Америку лопухом (Arctium). Самки могут запутываться лапами в материале для гнезда: с лапок птиц снимали клубки растительных волокон размером 18,0 × 17,5 мм. Обычно такие проблемы обнаруживались у неокольцованных птиц[26]. Положительный эффект, по-видимому, может оказывать высадка подходящих цветковых растений в местах гнездования. Вместе с тем искусственные кормушки могут задерживать осеннюю миграцию и увеличивать риск пострадать от нападений домашних кошек или столкнуться с окнами[25].
На взрослых рубиновогорлых колибри могут нападать хищные птицы — полосатый ястреб (Accipiter striatus), дербник (Falco columbarius), воробьиная пустельга (Falco sparverius), миссисипский коршун (Ictinia mississippiensis), а также крупные воробьинообразные — американский жулан (Lanius ludovicianus), балтиморская иволга (Icterus galbula), королевский тиранн (Tyrannus tyrannus). Однако они не являются значительной причиной смертности[26][16]. При нападении рубиновогорлые колибри скрываются в густых зарослях, но могут и атаковать хищника в ответ: во время осенней миграции колибри нападал на воробьиную пустельгу, пролетая неоднократно перед её лицом и планомерно атакуя её клюв, пока она не отступила. В другом случае самка рубиновогорлого колибри не принимала участие в атаке на голубую сойку (Cyanocitta cristata) другими птицами, хотя находилась поблизости. На рубиновогорлого колибри также могут нападать богомолы Tenodera sinensis, стрекозы, лягушки; известно о попадании птиц в сети пауков родов Argiope, Araneus, Neoscona. Около кормушек основным врагом является, по-видимому, домашняя кошка (Felis catus). Опасность близ кормушек также представляют жалящие насекомые, в особенности шершень (Vespa crabo), длина которого составляет 25—50 мм. Птенцов рубиновогорлого колибри может съесть голубая сойка, случаи потребления яиц колибри неизвестны[16].
Рубиновогорлые колибри могут переживать снегопады и плохую погоду поздней весной; осенью покидают территории на северо-востоке США, когда температура стабильно не превышает 0 °C. Вместе с тем во время холодной погоды в Алабаме была найдена самка рубиновогорлого колибри, которая умерла от голода, насиживая яйца[26]. Фатальными для рубиновогорлого колибри могут стать поражения клюва. У птиц часто обнаруживают вшей, информация о других паразитах или болезнях отсутствует[26].
Питание
Рацион

.jpg.webp)
Основу рациона рубиновогорлого колибри составляют цветочный нектар и мелкие насекомые[27].
Птицы предпочитают нектар красных трубчатых цветков как местных видов растений, так и завезённых садовых. Среди видов, на цветках которых кормится рубиновогорлый колибри, отмечены: конский каштан красный (Aesculus pavia var. pavia), Impatiens capensis, Impatiens pallida, Aquilegia canadensis, кампсис укореняющийся (Campsis radicans), Ipomoea coccinea, монарда дудчатая (Monarda fistulosa), монарда двойчатая (Monarda didyma), Castilleja coccinea, жимолость вечнозелёная (Lonicera sempervirens), жимолость японская (Lonicera japonica), жимолость канадская (Lonicera canadensis), лобелия пурпурная (Lobelia cardinalis), Silene regia, Silene rotundifolia, Silene virginica, гельземий вечнозелёный (Gelsemium sempervirens), Bignonia capreolata, Erythrina herbacea, спигелия мариландская (Spigelia marilandica), иногда кормится серёжками Salix amygdaloides. Кроме того, в рацион входит нектар растений родов шалфей (Salvia), юстиция (Justicia), многоколосник (Agastache), куфея (Cuphea). В муниципалитете Паленке в Мексике питаются на цветках растений рода инга; в провинции Гуанакасте в Коста-Рике — Ruellia inundata, Gliricidia sepium, Samanea saman, Thalia geniculata, инги съедобной (Inga edulis), Combretum farinosum, растений рода стахитарфета (Stachytarpheta); в Центральной долине в Коста-Рике — на цветках лантаны сводчатой (Lantana camara), растений родов Samanea, Stachytarpheta, гибискус (Hibiscus), инга, иногда питаются на Erythrina poeppigiana, которые вместе с растениями рода инга высаживают для создания тени на кофейных плантациях[27].
Подходящие для рубиновогорлого колибри цветки обычно характеризуются разбавленным нектаром. В среднем колибри перерабатывают лишь 7 % полученного нектара, и, возможно, он не является основным источником белков в рационе этого вида. Нектар цветков в Колорадо содержит заметное количество солей, предположительно схожая концентрация солей наблюдается в цветках на востоке Северной Америки, позволяя рубиновогорлому колибри поддерживать солевой баланс. Воду птицы получают вместе с нектаром[27].
Из насекомых в рацион рубиновогорлого колибри входят кровососущие комары (Culicidae) и другой гнус, плодовые мушки дрозофилы (Drosophila), мелкие пчёлы; также он поедает пауков. Подбирает с листьев и коры гусениц, тлей, яйца насекомых. Защищает территории с плодоносящими деревьями, на которых гниющие фрукты привлекают большое количество насекомых. На основе анализа помёта учёные полагают, что насекомые составляют около 50—60 % рациона рубиновогорлого колибри[27].
Когда нектара недостаточно, птицы могут питаться соком таких деревьев, как берёза бумажная (Betula papyrifera), берёза тополелистная (Betula populifolia), берёза аллеганская (Betula alleghaniensis), лириодендрон тюльпановый (Liriodendron tulipifera) и клён красный (Acer rubrum)[27]. Концентрация сахара в соке деревьев ниже, чем в нектаре, но она мало меняется в течение дня и, скорее всего, служит более надёжным источником энергии для самок в сезон размножения, чем нектар[27][21].
В неволе колибри могут питаться заменяющей нектар смесью из кукурузного сиропа, фруктового детского питания, калорийных добавок, протеинового порошка, витаминов для птиц и воды, а также живыми дрозофилами, оставаясь при этом здоровыми[27].
Добыча пропитания
Рубиновогорлый колибри обычно кормится в полёте, зависая около цветка на трепещущих крыльях; может кормиться сидя, если это позволяет строение растения или конструкция кормушки. Насекомых ловит на лету, захватывая их широкой поверхностью подклювья, дистальная (ближе к концу) часть которого способна отгибаться вниз. Кроме того, птицы могут собирать насекомых с листьев, коры или из паутины. Сок деревьев добывает через отверстия, созданные желтобрюхим дятлом-сосуном, за которым они могут следовать, чтобы всё время питаться свежим соком. Обычно рубиновогорлые колибри кормятся в светлое время суток. На севере Луизианы в сентябре были отмечены самец и самка, которые кормились ночью при искусственном освещении[27].
Рубиновогорлые колибри могут запоминать новые источники пищи, в частности кормушки. Они узнают их по расположению в пространстве и цвету. Представители вида обладают цветовым зрением, причём, кроме видимого диапазона, их глаза чувствительны также к ближнему ультрафиолету. Птицы выбирают конкретные цветки на основании энергетической ценности, а не их цвета. Вместе с тем предпочитают определённые места кормления, а также определённую форму цветков. Известно, что рубиновогорлый колибри различает первый день цветения Malvaviscus arboreus var. drumondii от второго: цветки первого дня предпочтительнее, так как они содержат больше нектара. При искусственном обогащении нектаром втородневных цветков эта разница стирается, то есть основным фактором при выборе является количество в них нектара, а не форма или цвет. Учёные полагают, что не исключено влияние обоняния: родственные черногорлые архилохусы при поиске кормушек научились различать обонятельные сигналы[27].
Несмотря на то, что у цветков, форма которых эволюционировала вместе с колибри, обычно отсутствует губа, нижняя губа монарды дудчатой и монарды двойчатой может облегчать питание рубиновогорлых колибри[27].
Экологическая роль

Рубиновогорлый колибри опыляет многие виды цветковых растений в восточной части Северной Америки. К опылению преимущественно этим видом, по-видимому, приспособлен кампсис укореняющийся. В Иллинойсе и Миссури рубиновогорлые колибри переносили в 10 раз больше пыльцы из расчёта на одно рыльце за один визит, чем шмели (Bombus) и медоносные пчёлы (Apis mellifera). Учёные отмечают особые отношения рубиновогорлого колибри с лобелией пурпурной: в начале сезона колибри кормятся, протыкая венчик, а со временем переходят на кормление непосредственно из цветка, таким образом опыляя его. Различное расположение пыльцы у монард предотвращает гибридизацию близкородственных видов[27].
Большинство подходящих для колибри цветковых растений на востоке США опыляются не только рубиновогорлыми колибри, в этом регионе птицы сыграли небольшую роль в их эволюции. На юго-востоке США отмечали рубиновогорлого колибри, кормящегося на некоторых цветковых растениях, предположительно опыляемых в основном бабочками рода Manduca. Возможно, рубиновогорлый колибри является основным опылителем редкого гименокаллиса вида Hymenocallis coronaria. Убрав рубиновогорлого колибри в качестве опылителя Silene regia и оставив растению только возможность опыления насекомыми, учёные отметили резкое снижение плодоносности. Возможно, большое производство нектара, а также форма цветков растений рода недотрога призваны привлекать для опыления рубиновогорлого колибри, а гибкие цветоножки способствуют его эффективности. Преимущественно рубиновогорлым колибри опыляется ирис буро-жёлтый (Iris fulva), в то время как шмели опыляют близкородственный Iris brevicaulis.
Метаболизм
При комфортной температуре окружающей среды птица массой 3,2 г в состоянии покоя потребляет 0,022 мл кислорода в минуту. В состоянии трепещущего полёта терморегуляции способствует генерируемое этим полётом мышечное тепло. У самцов затраты на полёт больше, чем у самок, что связано с различиями в форме крыла. Кроме того, высокие энергетические затраты вызывает линька[27].
Накапливать энергетические запасы для осеннего перелёта птицы начинают с начала июля. На юге Онтарио масса птиц перед миграцией повышается на 0,8 г в 4 дня. На юго-западе Южной Каролины у перелётных птиц дополнительная масса составляла 1,7—2,6 г, на побережье Алабамы — 0,53—2,80 г (в среднем 0,92 г), при этом скорость пополнения запасов составляла 0,10 г в день[23]. Однако часть птиц придерживается другой стратегии и не наращивает свою массу перед перелётом. Время, проведённое в утренние часы за добычей корма, у тех и других не отличается, при этом птицы, накапливающие запасы, больше питаются в середине дня, а наиболее усиленно — непосредственно перед наступлением темноты[28].
В связи с тем, что рубиновогорлый колибри возвращается на север в то время, когда ночные температуры ещё довольно низки (около +1 °C), а иногда возможны сильные заморозки (до −15 °C в Манитобе), по-видимому, они могут впадать в анабиоз. Птиц в таком состоянии отмечали в Алабаме во время резкого понижения температуры[27]. Готовящиеся к миграции рубиновогорлые колибри также могут впадать в анабиоз; учёные полагают, что это является одной из причин лишь незначительного уменьшения массы птиц в ночные часы[28].
Поведение
Полёт

Маленькие короткие лапки не позволяют рубиновогорлому колибри ходить или прыгать, он может перемещаться исключительно по воздуху. Энергозатратный трепещущий полёт этого вида колибри требует 204 калорий на грамм массы тела в час, частота биений крыльев при этом составляет 53 удара в секунду. Для сравнения: в покое рубиновогорлый колибри потребляет 20,6 кал/(г·ч). На основе экспериментов в газовых смесях различного состава учёные сделали вывод о существенных энергетических резервах рубиновогорлого колибри, по-видимому, необходимых для сложных манёвров во время ухаживания и переноса дополнительной массы в начале миграции. Увеличение массы птицы влияет на возможность осуществлять трепещущий полёт, оставаясь на одном месте, но не на максимальную скорость, а вот потеря перьев во время линьки влияет на оба типа перемещений. На трепещущий полёт большое влияние оказывает площадь первостепенных маховых перьев, при этом даже после потери 30 % площади поверхности крыла самка рубиновогорлого колибри способна к трепещущему полёту. Снижение аэродинамических способностей во время линьки компенсируется потерей массы[16].
В полёте рубиновогорлый колибри может поджимать или распускать хвост, а также издавать угрожающие звуки. Будучи на насесте, он дополнительно распускает крылья[16]. Спят птицы с вытянутой шеей, задранным немного вверх клювом и поднятыми перьями на теле[16].
Территориальное поведение
.jpg.webp)
В сезон размножения самец рубиновогорлого колибри активно защищает свою территорию, которая в первую очередь включает источники пищи и укрытия, а не места для спаривания. Если на территории самца оказывается недостаточно пищи для привлечения самок, то он может обосноваться в другом месте, переместившись на расстояние до трёх километров. При нарушении границ территории самцами своего вида рубиновогорлый колибри начинает издавать громкие звуки, громкость и интенсивность которых увеличивается, если соперник не покидает территорию. Взрослый самец преследует нарушителя, прогоняя его со своей территории, иногда происходят физические столкновения, которые включают в себя уколы клювом и удары лапами[16]. Рубиновогорлый колибри преследует и воробьинообразных птиц, проникающих на его территорию. Ранней весной в Манитобе рубиновогорлый колибри питается серёжками Salix amygdaloides, несмотря на то, что на растении обычно доминирует Setophaga tigrina[16].
Территориальное поведение менее заметно в другое время года. В конце лета рубиновогорлый колибри проигрывает охристому и многим другим более крупным видам колибри в штате Алабама, в Коста-Рике или в штате Халиско в Мексике. В Коста-Рике рубиновогорлый колибри удерживает территорию от Cynanthus canivetii, который немного уступает ему размерами. В ранние утренние часы самец может удерживать территорию при скоплении Ruellia inundata, когда отсутствуют более доминирующие колибри, в частности колибри-амазилии (Amazilia). Охрана территории продолжается и во время осенней миграции, птицы защищают искусственные кормушки на расстоянии до восьми метров[16].
Если пищевые ресурсы позволяют, территории самцов могут быть на расстоянии около 15 метров друг от друга. Более близкое соседство с другими представителями своего вида возможно лишь во время ухаживаний, кормления птенцов или при охране территорий[16].
Брачное поведение

Предположительно, у самца рубиновогорлого колибри может быть несколько самок, но, возможно, несколько партнёров бывает не только у самцов, но и у самок. Самец и самка остаются вместе только на период ухаживания и совокупления, всё остальное время птицы проводят в одиночестве. При строительстве гнезда самец может навещать самку и демонстрировать «челночные» движения, издавая при этом интенсивные позывки и звуки крыльями, которые самка игнорирует до завершения строительства гнезда[16].
Некоторые элементы поведения самца при появлении самки на его территории во время сезона размножения совпадают с обычным территориальным поведением, особенно в самом начале. Самец распускает перья воротника и начинает преследование самки, не вытесняя её при этом со своей территории. Затем самец начинает «нырять» (англ. Dive Display) — повторять в воздухе нырки и петли длиной дуги менее 1 метра, начиная с высоты 12—15 метров над самкой. Сразу после приземления самки на насест самец демонстрирует быструю серию горизонтальных перемещений из стороны в сторону на расстоянии менее 0,5 метра от самки, во время этого «челночного» движения (англ. Shuttle Display) он держит перья воротника распущенными и, помимо интенсивного шума, издаваемого крыльями, не производит никаких звуков. В это время самка может скрываться в густых зарослях и отвечать движениями головы, синхронными с полётом самца, иногда с закрытыми глазами. Согласная на копуляцию самка издаёт мяукающий звук, быстро распускает хвост в одну сторону, опускает крылья и может взмахнуть ими. Сразу после этого самец садится на самку и остаётся на её спине 2—3 секунды. Спаривание происходит на высоте менее 1,5 м от земли. После копуляции самка несколько секунд остаётся на ветке, попеременно распушая и прижимая контурные перья, самец сразу перелетает на высокий насест, с которого поёт беспорядочные громкие песни[16].
Размножение
Гнездо

Строительством гнезда занимается исключительно самка и начинает его сразу по прибытии в места размножения. Для гнезда она выбирает место ближе к концу нисходящей ветки с довольно открытой площадкой снизу и навесом сверху, который защищает гнездо от прямых солнечных лучей. Гнёзда часто расположены на растениях родов дуб (Quercus), граб (Carpinus), тополь (Populus), каркас (Celtis), сосна (Pinus), а также на берёзе аллеганской (Betula alleghaniensis). Иногда рубиновогорлый колибри строит гнездо около воды или непосредственно над водой. На юге Онтарио рубиновогорлые колибри отдают предпочтения лиственным деревьям[29]. В среднем гнездо расположено на высоте 5—7 м от земли, но высота может существенно варьироваться в пределах 0,5—15 м. Гнездо на высоте 0,5 м было обнаружено в Коннектикуте на побеге сумаха голого (Rhus glabra) и, скорее всего, не использовалось. Важным фактором при определении подходящего места является наличие поблизости нектара и насекомых[29].
Строительство гнезда продолжается 6—10 (изредка 5) дней. В первую очередь самка создаёт плоское основание гнезда из пуха бодяка (Cirsium) или одуванчика (Taraxacum), которое прикрепляет паутиной к верхней стороне ветки. Она утрамбовывает основание ногами, делая его жёстким, в отличие от стенок гнезда, для которых используется белый пух растений, чешуи почек и паутина. С помощью паутины она связывает растительные материалы, двигая клювом по контуру восьмёрки, дополнительно может использовать сосновую смолу. Прижимая материал между клювом и грудью и вращаясь в гнезде, самка формирует край гнезда. Снаружи гнездо замаскировано лишайниками. Самка может использовать материалы старого гнезда при строительстве нового, иногда для второй кладки она лишь подправляет гнездо. Наружный диаметр гнезда у основания составляет 45—50 мм, в верхней части чуть меньше; высота гнезда — 40 мм. Внутренний диаметр — 28 мм, глубина — 25—29 мм, толщина стенок — 10 мм[29].
Известно о двух самках в Мичигане, которые устроили гнёзда в 50 метрах друг от друга, в Кентукки минимальное расстояние составляло 69 метров[16].
Кладка
_(20797676905).jpg.webp)
Кладка обычно состоит из двух яиц, иногда из одного яйца, ещё реже — трёх. Из 77 исследованных гнёзд кладка из двух яиц встречалась в 66 случаях, одного яйца — в 10 случаях — и трёх яиц — в 1 случае[26][29].
Яйца рубиновогорлого колибри белые, гладкие, эллиптически-яйцевидной или субэллиптической формы. Их размеры составляют в среднем 12,99 мм × 8,61 мм (12,12—13,96 мм × 8,29—9,10 мм), масса по некоторым данным составляет 0,56 г, масса пустой скорлупы — 0,027 г (0,022—0,030 г). Информация о толщине скорлупы отсутствует[29].
В южной Луизиане первое яйцо было отмечено на четвёртой неделе марта. В центральных районах Алабамы самка рубиновогорлого колибри откладывает первое яйцо на второй неделе апреля. Второе яйцо обычно откладывается спустя 1—3 дня после первого. Яйца рубиновогорлого колибри находили с 25 мая по 2 сентября на юге Онтарио, с 29 мая по 6 августа — в Вермонте, с 24 мая по 22 июля — в Массачусетсе, с 9 мая по 28 августа — в Пенсильвании, с 27 апреля по 14 августа — в Арканзасе[29].
В Алабаме насиживание яиц, которое начинается после откладки первого яйца, продолжается 12—14 дней. В других регионах инкубация может длиться дольше; вместе с тем ранние сведения о продолжительности насиживания в 28 дней, по-видимому, неточны. В Мичигане самки находились в гнезде 72—75 % дневного времени, покидая его для кормления более, чем на 2 минуты, 5—6 раз в час. В ночное время температура воздуха могла опускаться до 10,8—15,5 °C, а температура инкубации составляла 27—31,3 °C. Из яиц, температура которых опустилась до ночной температуры воздуха 14 °C и сохранялась таковой на протяжении 5,5 часов, через 10 дней успешно появились птенцы, которые вылетели из гнезда (общая продолжительность инкубации составила 17 дней)[29].
В Алабаме самка рубиновогорлого колибри делает две кладки за сезон[26][29]. Наличие второй кладки возможно и в Нью-Брансуике, где 18 июля были обнаружены самки на яйцах. Схожие поздние даты, которые могут свидетельствовать о второй кладке, отмечались и в других частях ареала[26]. Строительство второго гнезда самка может начать во время ухаживания за птенцами первой кладки[29]. В Алабаме были зарегистрированы и третьи кладки, но частота их появления неизвестна[26][29].
В июле 1890 года в гнезде рубиновогорлого колибри было отмечено яйцо буроголового коровьего трупиала (Molothrus ater) — гнездового паразита, самки которого откладывают яйца в гнёзда других маленьких птиц, — которое полностью заполнило гнездо[29].
Птенцы
_(14595932567).jpg.webp)
Все обязанности по уходу за птенцами выполняет самка. Птенцы появляются на свет с интервалом 1—3 дня, однако детали этого события почти неизвестны. После появления птенцов самка выкидывает из гнезда часть яичной скорлупы, которая потом находится под гнездом[29].
Вылупившиеся птенцы почти голые, их кожа может быть синего, тёмно-сланцевого, тёмно-серого или розового цвета, при этом в одной кладке могут быть птенцы разных цветов (как тёмные, так и розовые). По бокам спины проходит два тонких ряда жёлто-белых волосков. Клюв птенцов желтоватый, мясистая ткань на внутренней части подклювья и горло окрашены в цвета от оранжевого до ярко-красного. Яйцевой зуб отмечен не был. Масса птенцов составляет 0,62 г, координация слабая, вместе с тем птенец способен поднять голову в ответ на мяукающие звуки самки[29].
Первые 18 дней птенцы прибавляют массу со скоростью 0,47 г в день. В возрасте 2—3 дней они способны испражняться через край гнезда, до этого самка выбрасывает помёт за пределы гнезда[29]. В возрасте 7—8 дней у птенцов начинают прорезаться перья, примерно в это же время открываются глаза, а к 9-му дню самка перестаёт насиживать (сразу после появления птенцов самка проводит на гнезде 86 % времени, в дальнейшем всё меньше и меньше). Птенцы пробуют летать в возрасте 15 дней, а к 18—20 дням приобретают полное оперение[29].
Старший птенец покидает гнездо в возрасте 18—22 дней, младший следует за ним по достижении аналогичного возраста. В ранних работах время вылета из гнезда варьирует от 14 до 28 дней, но, вероятно, является неточным. После вылета птенцов из гнезда самка продолжает их кормить ещё 4—7 дней. Масса молодых птиц составляет 4,85 г, что значительно превышает массу взрослых. После того как самка перестаёт их кормить, масса резко падает до тех пор, пока птенцы не разовьют навыки добывания корма. Через 30 дней после вылета из гнезда длина клюва птенцов достигает длины клюва взрослых птиц[29].
Кормление птенцов обычно начинается вскоре после вылупления, но иногда может быть отложено на период до 11 часов. Птенцы рубиновогорлого колибри, как и остальных колибри, раскрывают клюв в ответ на движение воздуха от крыльев самки или на её мяукающие сигналы. Самка кормит птенца, вставляя свой клюв в его и срыгивая пищу. Птенцам в возрасте 15—16 дней самка приносит зажатых в клюве насекомых[29]. Трёхдневные птенцы рубиновогорлого колибри просят еду и издают вой, цель которого остаётся неизвестной[17].
Успех размножения и дальнейшая выживаемость

Сведения об успехе размножения рубиновогорлого колибри отсутствуют. Выживаемость взрослых птиц не зависит от их возраста. В заповеднике Паудермилл на западе Пенсильвании средняя годовая выживаемость самцов составляла 31,2 %, самок — 42,3 %; в северных и центральных районах Южной Каролины эти показатели составляли 30 % и 43 % соответственно[26]. В Иллинойсе и Индиане ежегодно повторно отлавливалось от 6 до 16,1 % птиц (в среднем 9,2 %), а в горах Адирондак в штате Нью-Йорк в 1991—2011 годы — 24—43 %[26].
Среди взрослых птиц преобладают самки. Высокий уровень смертности самцов связан с меньшими размерами тела и большими затратами энергии на охрану территории[26][16]. В заповеднике Паудермилл коэффициент отлова самок и самцов первого года составлял 1,1 к 1 (самок несколько больше, чем самцов) и сильно смещался для взрослых птиц, составляя 1,4 к 1 весной и 4,1 к 1 осенью[16][11]. Схожее соотношение было отмечено в Огайо, Миссури, Северной Каролине и Мэриленде[11]. На севере Южной Каролины среди птиц первого года отмечается больше самцов (0,67 к 1), а среди взрослых особей — самок (1,5 к 1)[16].
Половозрелость рубиновогорлого колибри наступает, по-видимому, на втором году жизни, при этом из-за того, что молодые птицы прилетают позже взрослых и их линька ещё не завершена, они могут приступать к размножению позже. Непосредственные наблюдения молодых самок в Луизиане в начале апреля показывали накопление небольшого количества полостной жидкости в области яичника, указывающей на овуляцию, однако ни одной яйцеклетки обнаружено не было[26].
Максимальный известный возраст самки в США составлял 9 лет и 2 месяца. На юге Луизианы отмечали двух самок возрастом более 7 лет[26].
Систематика

Вид был описан создателем современной систематики Карлом Линнеем в десятом издании «Системы природы» в 1758 году под названием Trochilus colubris[30][31]. Ещё до этого вид приводился в работах английских натуралистов Марка Кейтсби (1729[32], Mellivora avis carolinensis) и Джорджа Эдвардса (1743[33], Mellivora gula rubra)[30]. Эдвардс при этом писал: «Мне сообщили, что никакая птица данного рода, кроме этой, никогда не посещает английские колонии в Северной Америке» (англ. I have been informed that no Birds of this Genus, except this one, ever visits the English Colonies in North America)[33]. Род колибри-архилохусы был выделен немецким зоологом Людвигом Райхенбахом в 1854 году[31].
Международный союз орнитологов не выделяет подвидов рубиновогорлого колибри[31][9]. В 1862 году на основании заспиртованного экземпляра, потерявшего окраску, американский орнитолог-любитель Джордж Ньюболд Лоренс описал вид Trochilus aurigularis, который позднее был признан синонимом Archilochus colubris[9].
Ближайшим видом, родственным рубиновогорлому колибри, является черногорлый архилохус (Archilochus alexandri). Учитывая отмечаемую время от времени гибридизацию, некоторые учёные рассматривают эти два вида как надвид. Случаи гибридизации участились после того, как ареал рубиновогорлых колибри расширился и начал включать в себя районы Оклахомы, Техаса и Луизианы, в которых уже обитали черногорлые архилохусы. Около 10 % самцов, пойманных на юго-западе Оклахомы, имели промежуточную окраску оперения воротника. Изучение двух гибридных особей, пойманных на северо-востоке Техаса и юго-востоке Луизианы, показало, что они совмещают особенности окраски оперения обоих предков. Исследование митохондриальной ДНК гибридного самца из Луизианы показало, что его мать принадлежала к виду рубиновогорлый колибри[9].
Род Archilocus является сестринским по отношению к кладе аметистовых колибри (Calliphlox) и колибри-пчёлок (Mellisuga). Эти три рода вместе образуют сестринскую связь с группой калипт (Calypte), колибри-селасфорусов (Selasphorus) и колибри-эльфов (Atthis), а все вместе они образуют так называемую группу колибри-пчёлок[9].
Примечания
- Галушин В. М., Дроздов Н. Н., Ильичёв В. Д. и др. Фауна мира: Птицы: Справочник / под ред. В. Д. Ильичёва. — М.: Агропромиздат, 1991. — С. 184, 185. — 311 с. — ISBN 5-10-001229-3.
- Коблик Е. А. Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). Часть 3 / науч. ред. к. б. н. М. В. Калякин. — М.: Издательство МГУ, 2001. — С. 68. — 360 с. — (Разнообразие животных). — ISBN 5-211-04072-4.
- Карташёв Н. Н. Систематика птиц: Учеб. пособие для ун-тов. — М.: Высшая школа, 1974. — С. 232. — 368 с.
- Харитонов С. П., Харитонова И. А., Коблик Е. А. Заметки о птицах полуострова Юкатан // Русский орнитологический журнал. — 2020. — Т. 29, № 1935. — С. 2659—2677.
- Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Русский язык, РУССО, 1994. — С. 156. — 2030 экз. — ISBN 5-200-00643-0.
- Гладков Н. А., Дроздов Н. Н. Отряд Стрижеобразные (Apodiformes) // Жизнь животных. Том 6. Птицы / под ред. В. Д. Ильичёва, А. В. Михеева, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1986. — С. 321. — 527 с.
- Ильяшенко В. Ю. Птерилография птенцов птиц мира: гоацинообразные, туракообразные, кукушкообразные, стрижеобразные, птицы-мыши, трогонообразные, ракшеобразные, птицы-носороги, дятлообразные, воробьинообразные. — М.: Товарищество научных изданий КМК, 2015. — С. 41. — 292 с. — ISBN 5-9906895-6-X.
- Birds of the World: Ruby-throated Hummingbird, Appearance.
- Birds of the World: Ruby-throated Hummingbird, Systematics.
- Zenzal Jr. T. J., Contina A. J., Kelly J. F., Moore F. R. Temporal migration patterns between natal locations of ruby-throated hummingbirds (Archilochus colubris) and their Gulf Coast stopover site (англ.) // Movement Ecology. — 2018. — Vol. 6. — doi:10.1186/s40462-017-0120-2.
- Mulvihill R. S., Leberman R. C., Wood D. S. A possible relationship between reversed sexual size dimorphism and reduced male survivorship in the Ruby-throated Hummingbird (англ.) // Condor. — 1992. — Vol. 94, iss. 480—489.
- Courter J. R., Johnson R. J., Bridges W. C., Hubbard K. G. Assessing Migration of Ruby-Throated Hummingbirds (Archilochus colubris) at Broad Spatial and Temporal Scales (англ.) // The Auk. — 2013. — Vol. 130, iss. 1. — P. 107—117. — doi:10.1525/auk.2012.12058.
- Howell S. N. G. Ruby-Throated Hummingbird // A Guide to the Birds of Mexico and Northern Central America. — U.S.A.: Oxford University Press, 1995. — P. 424. — 851 p. — ISBN 0-19-854012-4.
- Berns C. M., Adams D. C. Bill Shape and Sexual Shape Dimorphism between Two Species of Temperate Hummingbirds: Black-Chinned Hummingbird (Archilochus alexandri) and Ruby-Throated Hummingbird (A. colubris) (англ.) // The Auk. — 2010. — Vol. 127. — P. 626—635. — doi:10.1525/auk.2010.09213.
- Baltosser W. H. Annual molt in Ruby-Throated and Black-Chinned Hummingbirds (англ.) // The Condor. — 1995. — Vol. 97. — P. 484—491.
- Birds of the World: Ruby-throated Hummingbird, Behavior.
- Birds of the World: Ruby-throated Hummingbird, Sounds and Vocal Behavior.
- Birds of the World: Ruby-throated Hummingbird, Distribution.
- Tweit R. C. Ruby-throated Hummingbird (англ.). The Texas Breeding Bird Atlas (2008). Дата обращения: 3 июля 2021. Архивировано 11 июля 2021 года.
- Рубиновогорлый колибри (англ.). The IUCN Red List of Threatened Species. Дата обращения: 13 июня 2021.
- Miller R. S., Nero R. W. Hummingbird-sapsucker associations in northern climate (англ.) // Canadian Journal of Zoology. — 1983. — Vol. 61. — P. 1540—1546.
- Birds of the World: Ruby-throated Hummingbird, Habitat.
- Birds of the World: Ruby-throated Hummingbird, Movements and Migration.
- Солдаткин Е. Бесплатные пассажиры // Юный натуралист. — 1989. — № 3. — С. 36—37.
- Birds of the World: Ruby-throated Hummingbird, Conservation and Management.
- Birds of the World: Ruby-throated Hummingbird, Demography and Populations.
- Birds of the World: Ruby-throated Hummingbird, Diet and Fouraging.
- Hou L., Welch Jr. K. C. Premigratory ruby-throated hummingbirds, Archilochus colubris, exhibit multiple strategies for fuelling migration (англ.) // Animal Behavior. — 2016. — Vol. 121. — P. 87—99. — doi:10.1016/j.anbehav.2016.08.019.
- Birds of the World: Ruby-throated Hummingbird, Breeding.
- Linnaeus C. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. — Editio decima, reformata. — Stockholm: Holmiae. (Laurentii Salvii), 1758. — Vol. 1. — P. 120. — 824 p.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Hummingbirds (англ.). IOC World Bird List (v11.1) (20 января 2021). doi:10.14344/IOC.ML.11.1. Дата обращения: 13 июня 2021.
- Catesby M. The Natural History of Carolina, Florida and the Bahama Islands. — London: W. Innys and R. Manby, 1729. — Vol. 1. — P. 65.
- Edwards G. A Natural History of Uncommon Birds. — London: Printed for the author at the College of Physicians, 1743. — Vol. 1. — P. 38.
Литература
- Weidensaul S., Robinson T. R., Sargent R. R., Sargent M. B., Zenzal T. J. Ruby-throated Hummingbird (Archilochus colubris) (англ.). Birds of the World (22 июля 2019). doi:10.2173/bow.rthhum.01. Дата обращения: 13 июня 2021.