Прохибитин
Прохибити́н (англ. Prohibitin, PHB) — многофункциональный[1] белок, кодируемый у человека геном PHB[2]. Ген Phb выявлен у животных, грибов, растений и одноклеточных эукариот. На основе сходства с дрожжевыми PHB1 и PHB2 соответственно прохибитины делятся на два класса — прохибитины типа I и типа II. Все организмы, за исключением приматов и грызунов, имеют единственную копию гена прохибитина[3][4][5]. Название происходит от изначального предположения об ингибиторной роли белка в клеточном цикле[6].
Строение
У прохибитинов выделяют следующие домены: (1) N-концевая трансмембранная α-спираль (остатки 2—24), (2) эволюционно консервативный домен PHB (остатки 55—172), (3) C-концевой суперспиральный домен (остатки 175—252), (4) сигнал ядерного экспорта (NES, остатки 257—270)[7]. Домен PHB характерен и для иных белков каркаса семейства SPFH (stomatin/prohibitin/flotillin/HflK/C)[1][7]. C-Концевой домен участвует во взаимодействии PHB1 и PHB2[1]. Более 50 % аминокислотных остатков PHB1 и PHB2 идентичны друг другу[8].
PHB1 и PHB2 образуют гетеродимеры, формирующие кольцеобразные заякоренные в мембране структуры диаметром 20—25 нм и массой порядка 1,2 МДа из чередующихся субъединиц[1][8][9]. Стехиометрия этих структур надёжно не установлена[8].
Структура прохибитинов не установлена[1]. Проводилось моделирование структуры различными методами[10][11].
Локализация
Гены прохибитина эволюционно консервативны, обнаружены у всех исследованных видов эукариот, экспрессируются повсеместно[3]. Ген Phb (около 11 тысяч пар оснований, 7 экзонов) расположен в локусе BRCA1 хромосомного региона 17q21[12] и кодирует белок массой около 30 кДа[3]; ген Phb2 (около 7 тысяч пар оснований, 10 экзонов) расположен в хромосомном участке 12p13 и кодирует белок массой около 37 кДа[13]. У человека идентифицированы четыре псевдогена[3]. Нарушение экспрессии одного гена не влияет на экспрессию другого, но приводит к разрушению белка[9].
Прохибитины локализованы в различных клеточных органеллах: митохондриях[14], ядре[15][16][6], плазматической мембране[3][6], эндоплазматической сети[1], фагосомах макрофагов[1]. Наибольшая концентрация PHB наблюдается во внутренней митохондриальной мембране[17]; отсутствие его в цитозольной фракции свидетельствует о нерастворимости белка[3]. Локализация прохибитинов определяет их функцию, особенно в опухолевых клетках[1]. В частности, повышенная экспрессия PHB1 на поверхности таких клеток является важным фактором лекарственной устойчивости[1]. В нормальных клетках молочной железы PHB1 локализован преимущественно в митохондриях, а при раке молочной железы — в ядре; при этом действие противоракового препарат камптотецина приводит к транспорту прохибитина из ядра в митохондрии[1].
Функции
Первоначально считался суппрессором пролиферации клеток и опухолей[3]. Эта антипролиферативная активность проявляется различным образом, в частности, 3'-нетранслируемая область (3'-НТО) гена PHB обладает самостоятельной антипролиферативной активностью[6]. Мутации PHB человека сопутствуют спорадическому раку молочной железы[18][19]. Прохибитин транскрибируется в виде двух мРНК с различной длиной 3'-НТО. Предполагается, что более длинная 3'-нетранслируемая область может функционировать как транс-регуляторная некодирующая РНК[2][20].
В дрожжах является важным фактором, влияющим на продолжительность жизни[21].
Протективная роль PHB при недостатке холестерина связана с активацией холестерин-зависимого промотора[22]. Экспрессия PHB в матке индуцируется эстрогеном[22].
Играет важную роль в корректном развитии печени, однако повышенная экспрессия PHB может быть фактором риска при развитии рака печени[22]. Нарушения активности PHB1 в печени приводят к ожирению[7].
На клеточном уровне прохибитин может иметь различные функции.
Митохондриальная функция и морфология
Прохибитины митохондрий подвергаются посттрансляционной модификации[21]. Выступают в качестве шаперонов белков дыхательной цепи или структурного каркаса оптимальной морфологии митохондрий[6][1]. Недавно у прохибитинов было продемонстрировано положительное, а не отрицательное регуляторное действие на пролиферацию клеток как растений, так и мышей. Прохибитины участвуют в регуляции дыхательной активности митохондрий[23] и отвечают за стабильность организации и количество копий митохондриальной ДНК[9].
Митохондриальные PHB защищают клетки от окислительного стресса и связанных с ним воспалительных процессов[7][1].
Отсутствие прохибитина приводит к нарушениям в формировании крист митохондрий из-за нарушений в процессинге OPA1[8]. Целостность внутренней мембраны митохондрий при пониженной концентрации фосфатидилэтаноламина и кардиолипина также поддерживается прохибитином благодаря формированию функциональных липидных микродоменов[9].
Модуляция транскрипции
Оба человеческих прохибитина локализованы в ядре[6] и модулируют транскрипционную активность, прямо или косвенно взаимодействуя с различными факторами транскрипции, в том числе ядерными рецепторами. Тем не менее, доказательств ядерной локализации прохибитинов и их связывания с факторами транскрипции у других организмов (дрожжей, растений, Caenorhabditis elegans и др.) было найдено мало, поэтому возможно, что модуляция транскрипции — уникальная функция прохибитинов млекопитающих[24][25][26][27].
Прохибитин подавляет транскрипцию, опосредованную факторами семейства E2F[16]. В большинстве случаев имеет антипролиферативное действие, однако в некоторых случаях проявляет противоапоптотический эффект[6].
B-Лимфоциты
Прохибитин заякорен в плазматической мембране B-лимфоцитов N-концевым фрагментом; остальная часть белка экспонирована в цитоплазму[3]. Ассоциирован с рецептором антигенов IgM, как и прохибитин-подобный белок[3].
Роль в клеточной миграции и адгезии
Благодаря взаимодействию с C-Raf выступает в качестве модулятора адгезии и миграции эпителиальных клеток[28]. Подавление экспрессии PHB приводит к повышению адгезии и формированию кластеров в клеточных линиях злокачественных опухолей[28].
Рецепторная функция и роль в патогенезе инфекционных заболеваний
В клетках эпителия желудочно-кишечного тракта связывается с капсульным полисахаридом Vi Salmonella typhi, возбудителя брюшного тифа[28].
Прохибитин может выступать в качестве рецептора при проникновении вирусов в клетку. В частности, PHB1 человека может выступать в качестве рецептора для вируса чикунгунья (CHIKV)[29]. Проникновение вируса денге серотипа 2 в клетки комаров Aedes aegypti и A. albopictus также опосредуется прохибитинами[30]. О роли прохибитинов в вирусном патогенезе известно мало.
Показано взаимодействие прохибитина с гликопротеином ВИЧ-1, неструктурным белком nsp2 SARS-CoV, капсидным белком VP1 вируса ящура, вирусным онкопротеином SV40Tag[1].
При персистенции вируса гепатита C экспрессия вирусного корового белка приводит к снижению концентрации прохибитина, нарушению его взаимодействия с оксидазой цитохрома c и, как следствие, к окислительному стрессу, являющемуся причиной онкогенеза при гепатите C[31][7].
Регуляция
Регуляция прохибитинов осуществляется путём посттрансляционных модификаций — фосфорилирования остатков серина, треонина и тирозина (в частности, протеинкиназой Akt), гликозилирования, трансамидирования, нитрозилирования остатков тирозина[1].
Пальмитирование PHB по остатку цистеина-69 усиливает взаимодействие его с мембраной, транслокацию, фософрилирование по остатку тирозина и связывание с белком липидных рафтов Eps 15 homology domain protein 2[32]
PHB2 регулируется вторичным мессенджером сфингозин-1-фосфатом[1].
Прохибитины как биологическая мишень действия низкомолекулярных соединений
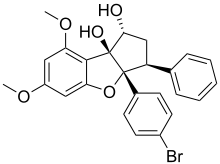
Наиболее изученные модуляторы прохибитинов относятся к классу флаваглинов[1], первоначально идентифицированных в растениях рода Аглайя[33]. Их действие на прохибитины проводит к ингибированию сигнального пути Ras—C-Raf—MEK—ERK[1], критически важного для выживаемости раковых клеток. Синтетический флаваглин FL3 обладает цитопротекторным действием в отношении кардиомиоцитов[1]. Положительный эффект флаваглинов был продемонстрирован в моделях рака, воспалительных и сердечно-сосудистых заболеваний, однако клинические испытания не проводились[1].
Взаимодействие PHB1 с меланогенином оказывает влияние на пигментацию млекопитающих[1].
Повышенная экспрессия PHB, локализованного на плазматической мембране, приводит к устойчивости раковых клеток к действию паклитаксела[22].
Взаимодействие PHB2 с капсаицином приводит к нарушению взаимодействия прохибитина с ANT2, выбросу прохибитина из митохондрий и транслокации его в ядро, разрушению митохондрий, выбросу цитохрома c в цитоплазму и индукции апоптоза[34].
Взаимодействия с другими белками
Выявлены взаимодействия прохибитинов более чем с 60[1] белками:
Список литературы
- Thuaud, Frédéric. Prohibitin Ligands in Cell Death and Survival: Mode of Action and Therapeutic Potential / Frédéric Thuaud, Nigel Ribeiro, Canan G. Nebigil … [и др.] // Chemistry & Biology. — 2013. — Vol. 20, no. 3. — P. 316—331. — doi:10.1016/j.chembiol.2013.02.006.
- Entrez Gene: PHB prohibitin.
- McClung, J. K. Prohibitin: potential role in senescence, development, and tumor suppression / J. K. McClung, E. R. Jupe, X. T. Liu … [и др.] // Experimental Gerontology. — 1995. — Vol. 30, no. 2. — P. 99–124. — doi:10.1016/0531-5565(94)00069-7. — PMID 8591812.
- Van Aken O; Pecenkova T; van de Cotte B; De Rycke, Riet; Eeckhout, Dominique; Fromm, Hillel; De Jaeger, Geert; Witters, Erwin; Beemster, Gerrit T. S. Mitochondrial type-I prohibitins of Arabidopsis thaliana are required for supporting proficient meristem development (англ.) // The Plant Journal : journal. — 2007. — Vol. 52, no. 5. — P. 850—864. — doi:10.1111/j.1365-313X.2007.03276.x. — PMID 17883375.
- Mishra S., Murphy L.C., Murphy L.J. The Prohibitins: emerging roles in diverse functions (англ.) // Journal of Cellular and Molecular Medicine : journal. — 2006. — Vol. 10, no. 2. — P. 353—363. — doi:10.1111/j.1582-4934.2006.tb00404.x. — PMID 16796804.
- Mishra, Suresh. Prohibitin: a potential target for new therapeutics / Suresh Mishra, Leigh C. Murphy, B. L. Gregoire Nyomba … [и др.] // Trends in Molecular Medicine. — 2005. — Vol. 11, no. 4. — P. 192–197. — doi:10.1016/j.molmed.2005.02.004. — PMID 15823758.
- Theiss, Arianne L. The role and therapeutic potential of prohibitin in disease / Arianne L. Theiss, Shanthi V. Sitaraman // Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. — 2011. — Vol. 1813, no. 6. — P. 1137-1143. — doi:10.1016/j.bbamcr.2011.01.033.
- Merkwirth, Carsten. Prohibitin function within mitochondria: Essential roles for cell proliferation and cristae morphogenesis / Carsten Merkwirth, Thomas Langer // Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. — 2009. — Vol. 1793, no. 1. — P. 27–32. — doi:10.1016/j.bbamcr.2008.05.013.
- Osman, Christof. Prohibitins and the functional compartmentalization of mitochondrial membranes / Christof Osman, Carsten Merkwirth, Thomas Langer // Journal of Cell Science. — 2009. — Vol. 122. — P. 3823-3830. — doi:10.1242/jcs.037655.
- 1LU7
- Winter, Anja. Molecular modeling of prohibitin domains / Anja Winter, Outi Kämäräinen, Andreas Hofmann // Proteins: Structure, Function, and Bioinformatics. — 2007. — Vol. 68, no. 1. — P. 353—362. — doi:10.1002/prot.21355.
- White, John J. Assignment of the human prohibitin gene (PHB) to chromosome 17 and identification of a DNA polymorphism / John J. White, David H. Ledbetter, Roger L. Eddy Jr. … [и др.] // Genomics. — 1992. — Vol. 11, no. 1. — P. 228–230. — doi:10.1016/0888-7543(91)90126-Y. — PMID 1684951.
- Chowdhury I, Garcia-Barrio M, Harp D, Thomas K, Matthews R, Thompson WE. The emerging roles of prohibitins in folliculogenesis (англ.) // Frontiers in Bioscience. — Frontiers in Bioscience, 2012. — Vol. 4. — P. 690—699. — doi:10.2741/410. — PMID 22201905.
- Ikonen E., Fiedler K., Parton R.G., Simons K. Prohibitin, an antiproliferative protein, is localized to mitochondria (англ.) // FEBS Letters : journal. — 1995. — Vol. 358, no. 3. — P. 273—277. — doi:10.1016/0014-5793(94)01444-6. — PMID 7843414.
- Fusaro G., Dasgupta P., Rastogi S., Joshi B., Chellappan S. Prohibitin induces the transcriptional activity of p53 and is exported from the nucleus upon apoptotic signaling (англ.) // Journal of Biological Chemistry : journal. — 2003. — November (vol. 278, no. 48). — P. 47853—47861. — doi:10.1074/jbc.M305171200. — PMID 14500729.
- Wang, Sheng. Prohibitin co-localizes with Rb in the nucleus and recruits N-CoR and HDAC1 for transcriptional repression / Sheng Wang, Gina Fusaro, Jaya Padmanabhan … [и др.] // Oncogene. — 2002. — Vol. 21, no. 55. — P. 8388-8396. — doi:10.1038/sj.onc.1205944.
- Tatsuta T., Model K., Langer T. Formation of Membrane-bound Ring Complexes by Prohibitins in Mitochondria (англ.) // Molecular Biology of the Cell : journal. — 2005. — January (vol. 16, no. 1). — P. 248—259. — doi:10.1091/mbc.E04-09-0807. — PMID 15525670.
- Sato T, Saito H, Swensen J, Olifant A, Wood C, Danner D, Sakamoto T, Takita K, Kasumi F. The human prohibitin gene located on chromosome 17q21 is mutated in sporadic breast cancer (англ.) // Cancer Research. — American Association for Cancer Research, 1992. — Vol. 52. — P. 1643—1646. — PMID 1540973.
- Sato T; Sakamoto T; Takita K; Saito, Hiroko; Okui, Keiko; Nakamura, Yusuke. The human prohibitin (PHB) gene family and its somatic mutations in human tumors (англ.) // Genomics : journal. — 1993. — Vol. 17, no. 3. — P. 762—764. — doi:10.1006/geno.1993.1402. — PMID 8244394.
- Jupe ER; Liu XT; Kiehlbauch JL; McClung, JK; Dell'Orco, R.T. The 3' untranslated region of prohibitin and cellular immortalization (англ.) // Experimental Cell Research : journal. — 1996. — Vol. 224, no. 1. — P. 128—135. — doi:10.1006/excr.1996.0120. — PMID 8612677.
- Dell'Orco, R. T. Prohibitin and the senescent phenotype / R. T. Dell'Orco, J. K. McClung, E. R. Jupe … [и др.] // Experimental Gerontology. — 1996. — Vol. 31, no. 1–2. — P. 245–252. — doi:10.1016/0531-5565(95)02009-8. — PMID 8706794.
- Zhou, Tian-Biao. Signaling pathways of prohibitin and its role in diseases / Tian-Biao Zhou, Yuan-Han Qin // Journal of Receptors and Signal Transduction. — 2013. — Vol. 33, no. 1. — P. 28-36. — doi:10.3109/10799893.2012.752006.
- Coates PJ; Nenutil R; McGregor A; Picksley, SM; Crouch, DH; Hall, PA; Wright, E.G. Mammalian prohibitin proteins respond to mitochondrial stress and decrease during cellular senescence (англ.) // Experimental Cell Research : journal. — 2001. — Vol. 265, no. 2. — P. 262—273. — doi:10.1006/excr.2001.5166. — PMID 11302691.
- Montano M.M., Ekena K., Delage-Mourroux R., Chang W., Martini P., Katzenellenbogen B.S. An estrogen receptor-selective coregulator that potentiates the effectiveness of antiestrogens and represses the activity of estrogens (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — June (vol. 96, no. 12). — P. 6947—6952. — doi:10.1073/pnas.96.12.6947. — PMID 10359819.
- Gamble S.C., Chotai D., Odontiadis M., Dart D.A., Brooke G.N., Powell S.M., Reebye V., Varela-Carver A., Kawano Y., Waxman J., Bevan C. Prohibitin, a protein downregulated by androgens, represses androgen receptor activity (англ.) // Oncogene : journal. — 2007. — March (vol. 26, no. 12). — P. 1757—1768. — doi:10.1038/sj.onc.1209967. — PMID 16964284.
- Kurtev V., Margueron R., Kroboth K., Ogris E., Cavailles V., Seiser C. Transcriptional regulation by the repressor of estrogen receptor activity via recruitment of histone deacetylases (англ.) // Journal of Biological Chemistry : journal. — 2004. — June (vol. 279, no. 23). — P. 24834—24843. — doi:10.1074/jbc.M312300200. — PMID 15140878.
- Park S.E., Xu J., Frolova A., Liao L., O'Malley B.W., Katzenellenbogen B.S. Genetic Deletion of the Repressor of Estrogen Receptor Activity (REA) Enhances the Response to Estrogen in Target Tissues In Vivo (англ.) // Molecular and Cellular Biology : journal. — 2005. — March (vol. 25, no. 5). — P. 1989—1999. — doi:10.1128/MCB.25.5.1989-1999.2005. — PMID 15713652.
- Rajalingam, Krishnaraj. Ras-Raf signaling needs prohibitin / Krishnaraj Rajalingam, Thomas Rudel // Cell Cycle. — 2005. — Vol. 4, no. 11. — P. 1503–1505. — doi:10.4161/cc.4.11.2142. — PMID 16294014.
- Wintachai P., Wikan N., Kuadkitkan A., Jaimipuk T., Ubol S., Pulmanausahakul R., Auewarakul P., Kasinrerk W., Weng W.Y., Panyasrivanit M., Paemanee A., Kittisenachai S., Roytrakul S., Smith D.R. Identification of prohibitin as a Chikungunya virus receptor protein (англ.) // Journal of Medical Virology : journal. — 2012. — Vol. 84, no. 11. — P. 1757—1770. — doi:10.1002/jmv.23403. — PMID 22997079.
- Kuadkitkan A., Wikan N., Fongsaran C., Smith D.R. Identification and characterization of prohibitin as a receptor protein mediating DENV-2 entry into insect cells (англ.) // Virology : journal. — 2010. — Vol. 406, no. 1. — P. 149—161. — doi:10.1016/j.virol.2010.07.015. — PMID 20674955.
- Koike, Kazuhiko. The oncogenic role of hepatitis C virus // Viruses and Human Cancer : From Basic Science to Clinical Prevention / Mei Hwei Chang ; Kuan-Teh Jeang, eds. — Berlin, Heidelberg : Springer, 2014. — P. 97-111. — (Recent Results in Cancer Research ; vol. 193). — ISBN 978-3-642-38965-8. — doi:10.1007/978-3-642-38965-8_6.
- Mishra, Suresh. The role of prohibitin in cell signaling / Suresh Mishra, Sudharsana R. Ande, B. L. Grégoire Nyomba // FEBS Journal. — 2010. — Vol. 277, no. 19. — P. 3937-3946. — doi:10.1111/j.1742-4658.2010.07809.x.
- Basmadjian, Christine. Flavaglines: potent anticancer drugs that target prohibitins and the helicase eIF4A / Christine Basmadjian, Frédéric Thuaud, Nigel Ribeiro … [и др.] // Future Medicinal Chemistry. — 2013. — Vol. 5, no. 18. — P. 2185-2197. — doi:10.4155/fmc.13.177.
- Kuramori, Chikanori. Capsaicin binds to prohibitin 2 and displaces it from the mitochondria to the nucleus / Chikanori Kuramori, [et al.] // Biochemical and Biophysical Research Communications. — 2009. — Vol. 379, no. 2. — P. 519-525. — doi:10.1016/j.bbrc.2008.12.103.
- Bacher S., Achatz G., Schmitz M.L., Lamers M.C. Prohibitin and prohibitone are contained in high-molecular weight complexes and interact with alpha-actinin and annexin A2 (англ.) // Biochimie : journal. — 2002. — December (vol. 84, no. 12). — P. 1207—1220. — doi:10.1016/s0300-9084(02)00027-5. — PMID 12628297.
- Wang S., Nath N., Fusaro G., Chellappan S. Rb and prohibitin target distinct regions of E2F1 for repression and respond to different upstream signals (англ.) // Molecular and Cellular Biology : journal. — 1999. — November (vol. 19, no. 11). — P. 7447—7460. — PMID 10523633.
- Joshi B., Ko D., Ordonez-Ercan D., Chellappan S.P. A putative coiled-coil domain of prohibitin is sufficient to repress E2F1-mediated transcription and induce apoptosis (англ.) // Biochemical and Biophysical Research Communications : journal. — 2003. — December (vol. 312, no. 2). — P. 459—466. — doi:10.1016/j.bbrc.2003.10.148. — PMID 14637159.
- Wang S., Zhang B., Faller D.V. Prohibitin requires Brg-1 and Brm for the repression of E2F and cell growth (англ.) // The EMBO Journal : journal. — 2002. — June (vol. 21, no. 12). — P. 3019—3028. — doi:10.1093/emboj/cdf302. — PMID 12065415.
- Wang S., Nath N., Adlam M., Chellappan S. Prohibitin, a potential tumor suppressor, interacts with RB and regulates E2F function (англ.) // Oncogene : journal. — 1999. — June (vol. 18, no. 23). — P. 3501—3510. — doi:10.1038/sj.onc.1202684. — PMID 10376528.
- Rasmussen RK; Ji H; Eddes JS; Moritz, Robert L.; Reid, Gavin E.; Simpson, Richard J.; Dorow, Donna S. Two-dimensional electrophoretic analysis of mixed lineage kinase 2 N-terminal domain binding proteins (англ.) // Electrophoresis : journal. — 1998. — Vol. 19, no. 5. — P. 809—817. — doi:10.1002/elps.1150190535. — PMID 9629920.