Глоссарий анатомии динозавров
В этом глоссарии объясняются технические термины, обычно используемые при описании окаменелых остатков динозавров. Помимо терминов, связанных с динозаврами, глоссарий охватывает термины с более широким употреблением, когда они имеют центральное значение при изучении динозавров или когда их обсуждение в контексте динозавров полезно. Глоссарий не охватывает ихнологические и гистологические термины, а также измерительные величины.
А
Акромион[1] (лат. acromion) — костный гребень на внешнем дистальном конце лопатки, который обеспечивает прикрепление ключицы[2][3]. У нодозаврид развивается выраженная шпора, известная как псевдоакромион, которая, вероятно, образовывала место прикрепления musculus scapulohumeralis anterior, и поэтому аналогична акромиону млекопитающих[4].
.JPG.webp)
Аксония (англ. axony) — термин, описывающий расположение доминирующего (самого большого и наиболее выступающего) пальца на кисти или стопе и чаще всего использующийся при описании следов. Если доминирующим является центральный палец (обычно, третий), как в случае с анатомией стопы большинства бипедальных динозавров, то такое расположение пальца называется мезаксоничным. У энтаксоничной кисти или стопы доминирующий палец располагается медиально (первый или второй палец); это редкое расположение можно найти на ногах завропод. И наоборот, у эктаксоничной кисти или стопы доминирующий палец находится снаружи (часто это четвёртый палец). Когда присутствуют два или четыре пальца и ни одна половина руки или ноги не доминирует над другой, это состояние называется параксоничным[5].
Арктометатарзальная стопа — стопа, у которой ближняя часть средней плюсневой кости зажата между окружающими её другими плюсневыми костями. Эта структура считается гомоплазией, развившейся независимо у различных групп меловых целурозавровых динозавров: тираннозавроидов, орнитомимозавров и троодонтид[6][7]. Известно, что наличие арктометатарзуса и более жёсткая щиколотка коррелируют с эффективным, быстрым бегом и свойственны для активных хищников[8].
Атлант (лат. atlas) — первый (передний) позвонок позвоночного столба. Он принимает затылочный мыщелок (лат. occipital condyle) основания черепа, образуя соединение между черепом и позвоночником. У динозавров были одиночные мыщелки, позволяющие вращать и атлант, и череп. У млекопитающих, напротив, видны двойные мыщелки, ограничивающие движение в одной плоскости; здесь вращательное движение в основном происходит между атлантом и осью второго позвонка. Атлант динозавров является примитивным, так как состоит из трёх отдельных элементов, отражающих базальное состояние рептилий: неврапофиза атланта (англ. atlantal neurapophysis), расположенного в верхней части позвонка[9], интерцентр (лат. intercentrum)[10]:191 спереди и тело позвонка (centrum) сзади. Перед дугой позвонка расположена дополнительная небольшая кость, proatlas. У динозавров эти три элемента обычно остаются разтдельными, но могут и сливаться вместе. У цератопсов атлант, а также второй и третий позвонки слиты в одну кость[11].
Б
Базальный бугор затылка[12]:340—342, или клиновидно-затылочный бугор[13] (англ. basal tuber, англ. sphenoccipital tubercle) — каждый из двух бугоркообразных расширений на нижней стороне черепной коробки, которые функционируют как места прикрепления нижних мышц шеи. Они образованы базиокципитальной и базисфеноидной костями и сильно выражены у ящеротазовых и умеренно — у птицетазовых[14][15].
Базикраний[16]:301 (лат. basicranium) — дно черепной коробки, сформированное такими элементами, как основная затылочная кость, основная клиновидная кость и парасфеноид[17].
Базисфеноид[16]:301, или основная клиновидная кость[10]:192 (лат. basisphenoid) — кость, образующая переднюю часть дна черепной коробки. Не видна снаружи в сочленённом черепе, сливается с основной затылочной сзади и с parasphenoid спереди. Пара отличительных отростков (лат. processi basipterygoidei, англ. basipterygoid processes), отходит от её нижней стороны и сочленяется с крыловидными отростками нёба[18].

Бедренная кость (лат. femur) — проксимальный элемент задней конечности. Головка бедренной кости направлена медиально и образует угол в 90 °, сочленяясь с вертлужной впадиной и образуя тазобедренный сустав. В верхней части бедренной кости есть несколько вертелов (отростков для прикрепления мышц)[19]:15:
- малый вертел (лат. trochanter minor, англ. lesser trochanter; также: передний вертел) расположен на передней поверхности сразу под головкой бедра. Такое расположение является уникальной особенностью для динозавров и ближайших к ним групп. К малому вертелу крепится musculus puboischiofemoralis, которая тянет заднюю конечность вперёд и внутрь и поэтому он имеет решающее значение для передвижения. Многие ранние динозавры дополнительно имели костный гребень или мышечный рубец, известный как вертельный выступ, который простирался от основания малого вертела к заднебоковому краю бедренной кости. Малый вертел у динозавров не гомологичен малому вертелу млекопитающих[19]:19[4]:179;
- большой вертел (лат. trochanter major, англ. greater trochanter; также: дорсолатеральный вертел) представляет собой гребень или выпуклость на боковой поверхности, направленную противоположно и продолжающую головку бедренной кости[20]:47. Он обеспечивает прикрепление двух мышц: m. puboischiofemoralis и m. ischiotrochantericus[4]:181;
- четвёртый вертел (англ. fourth trochanter; также: четвёртый трохантер[21]:32) — обширный фланец на задней поверхности бедренной кости, направленный назад. У динозавров четвёртый вертел резко асимметричен, проксимальная часть расширена больше, чем дистальная. Это синапоморфия динозавров. Четвертый вертел обеспечивал прикрепление длинной хвостово-бедренной мышцы (лат. m. caudofemoralis longus) — наиболее важной локомоторной мышцы, которая тянет бедро назад при сокращении, обеспечивая толчок[19]:16[4]:181-182.
Боковая затылочная кость[10]:192 (англ. exoccipitale, англ. exoccipital) — парная кость, которая вместе с непарной верхней затылочной костью (лат. supraoccipitale) и основной затылочной костью (лат. basioccipitale) образует затылочный бугор (англ. oxiput) в задней части черепа. У взрослых динозавров боковая затылочная кость сливается с опистотической костью, образуя структуру, также известную как экзокципитально-опистотическая. Своей нижней частью сливаются с базиокципиталом[18][19]:38.
Большеберцовая кость (лат. tibia) — бо́льшая из двух костей голени. Расположена медиальнее малоберцовой кости. Большеберцовая кость имеет выступающий гребень, идущий от её передней поверхности на её верхнем конце — кнемиальный гребень[12]:73 (также: большеберцовый гребень, англ. tibial crest). Кнемиальный гребень закреплял сухожилия трёхглавой мышцы бедра, которые отвечали за разгибание в колене (растяжение ноги), и увеличивал силу этих мышц[4]:178[22]:147.
В
Венечный отросток[16]:251 (англ. coronoid process) — костный выступ, идущий вверх от верхней поверхности нижней челюсти за зубным рядом. Служит местом прикрепления мышц, закрывающих челюсть; более высокий венечный отросток увеличивает плечо приложения силы этих мышц и, следовательно, силу укуса[23][24].
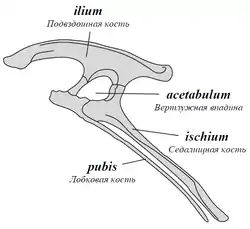
Вертлужная впадина[12]:340—342, или тазобедренная впадина (лат. acetabulum) — отверстие в тазу, образованное подвздошной, лобковой и седалищной костями, которое видно на латеральной и медиальной проекциях скелета. Она вмещает головку бедренной кости, образуя тазобедренный сустав. Большинство четвероногих имеет не перфорированную вертлужную впадину, лунка которой полностью заполнена костью, образуя углубление. Динозавры уникальны тем, что демонстрируют открытую, или перфорированную, вертлужную впадину, где вместо лунки расположено отверстие без заполняющей её кости[22]. Ориентация и положение вертлужной впадины — одна из основных морфологических черт, позволявшая динозаврам передвигаться в вертикальном положении, с ногами, расположенными под телом. У относительно небольшого числа динозавров, особенно анкилозавров (например, Texasetes pleurohalio), имеется закрытая вертлужная впадина, которая не является отверстием, а вместо этого напоминает неглубокое вогнутое углубление с каждой стороны тазового пояса[25].
Верхняя затылочная кость[10]:192 (лат. supraoccipitale) — парная кость в задней части черепной коробки. У взрослых динозавров она всегда сливается с парными боковыми затылочными костями (лат. exoccipitale), образуя структуру, известную как exoccipital-opisthotics. Вместе с боковыми затылочными верхняя затылочная кость образуют пару больших крыловидных отростков, идущих назад и в стороны от головного мозга, — парокципитальных отростков[12]:11. Эти отростки контактируют с теменной, чешуйчатой и квадратной костями[18][19]:38.
Верхняя челюсть (лат. maxilla) — парная кость черепа, несущая зубы. Спереди соединяется с предчелюстной костью и часто отмечает границу наружной ноздри. На большей части своей длины верхнечелюстная кость образует вентральную границу предглазничного окна, а также углубление, окружающее это окно — предглазничную ямку, которая расположена глубоко внутри верхней челюсти. У производных теропод верхнечелюстная кость может иметь 2 дополнительных отверстия меньшего размера: переднечелюстное отверстие и верхнечелюстное отверстие. На своем заднем конце верхняя челюсть сочленяется со скуловой и слёзной костями[19]:37[22]:139-140.
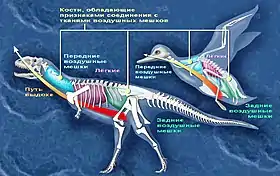
Воздушные мешки (англ. air sacs) — тонкостенные полупрозрачные наполненные воздухом мешки, соединённые с лёгким, присутствующие в организмах современных птиц. Вместе с лёгкими воздушные мешки образуют высокоэффективную дыхательную систему, которая у птиц способна извлекать до 160 % больше кислорода, чем это возможно у млекопитающих. Хотя внутри воздушных мешков происходит только ограниченный газообмен, они обеспечивают полноценную вентиляцию лёгких. Воздушные мешки, расположенные как сзади, так и спереди от лёгкого, обеспечивают постоянный поток воздуха через лёгкое. Меньшие по размеру и слепые наполненные воздухом мешки, выходящие из воздушных мешков и лёгких, известны как пневматические дивертикулы (лат. diverticula, ед. ч. diverticulum); они могут быть многочисленными и присутствовать в большей части тела. Некоторые пневматические дивертикулы проникают в кости по мере роста животного, создавая ямки (углубления) на костной поверхности, а также внутренние камеры внутри костей. Этот процесс известен как посткраниальная скелетная пневматизация. Хотя воздушные мешки не способны окаменеть, их присутствие, по крайней мере, у ящеротазовых, угадывается по характерным следам пневматизации в костях[26].
Вторичное нёбо (англ. secondary palate) — структура, напоминающая крышу, отделяющую носовые дыхательные пути от рта. Хоаны примитивных рептилий открываются в ротовую полость в передней части рта, что допускало блокировку дыхательных путей пищей во время питания. Вторичное нёбо, которое обычно встречается у млекопитающих, смещает хоаны назад, позволяя питаться и дышать одновременно[27]. У представителей нескольких групп, включая анкилозавров[28], спинозаврид[29], и целурозавров из группы манирапториформ[30], вторичное нёбо было костным. Напротив, мягкое вторичное нёбо предполагалось для некоторых других таксонов, таких как род платеозавр и инфраотряд стегозавры, на основании выступов и гребней нёбных костей[31][32]. Костное вторичное нёбо обычно образовано выступами предчелюстных и верхнечелюстных костей, которые встречаются с сошником на средней линии черепа. Анкилозавриды развили новую дополнительную часть вторичного нёба, которая простиралась вперёд от крыловидной и нёбной костей к задней части нёба, и которая называется каудовентральной нёбной полкой (англ. caudoventral palatal shelf). Соответственно, передняя часть, образованная предчелюстными и верхнечелюстными костями, называется ростродорсальной нёбной полкой (англ. rostrodorsal palatal shelf)[28].
Г

Гастралии, или брюшные рёбра[10]:192 (лат. gastralia, мн. ч.; ед. ч. — гастралий (лат. gastralium)) — часть кожного скелета, сухожильные образования, расположенные в дерме живота между грудиной и лобком. Гастралии примитивно присутствуют у динозавров и могут быть найдены у базальных завроподоморфов и теропод, но были независимо утеряны у завропод, птицетазовых и современных птиц. Обычно расположены в 8—21 ряд, каждый из которых состоит из четырёх элементов: правый и левый медиальные гастралии сочленяются по средней линии тела зигзагообразно, а латеральный гастралий сочленяется с боковым концом каждого медиального гастралия. Первый и последний ряды могут объединяться в единую дугообразную структуру в форме шеврона. Таким образом, все гастралии связаны друг с другом и работают как единое функциональное целое. Использование термина «брюшное ребро», которое иногда используется для обозначения гастралии и других структур брюшной полости, неоднозначно, поэтому не рекомендуется[33].
Гленоид[12]:68 (лат. glenoideum, англ. glenoid) — суставная поверхность, образованная коракоидом и лопаткой, которая принимает головку плечевой кости, образуя плечевой сустав. Это вогнутая круглая структура, окаймлённая костным ободком и расположенная на задней поверхности коракоида и дальней поверхности лопатки[3][22]:144.
Д
Дельтопекторальный гребень (англ. deltopectoral crest) — направленный вперёд костный выступ в верхней части плечевой кости. Особенно длинный и выступающий дельтопекторный гребень — синапоморфия динозавров, то есть признак, отличающий эту группу от других групп. У динозавров гребень составляет 30—40 % длины плечевой кости. Он обеспечивал поверхности для прикрепления мышц плеча и груди (дельтовидная и грудная мышцы соответственно), которые при сокращении притягивали переднюю конечность к туловищу[19]:14.
Добавочная надглазничная кость[12]:341 (англ. palpebral bone, также: supraorbital, adlacrimal[34]) — небольшая парная кость, расположенная на верхнем крае глазницы или образующая её часть. Происходит от остеодермы. С каждой стороны черепа может быть одна или несколько таких костей. Их наличие является синапоморфией птицетазовых динозавров, хотя этот костный элемент эволюционировал и в других кладах рептилий, включая группу Crocodyliformes[35].
Дуга позвонка[36]:18 (лат. arcus vertebrae, англ. neural arch) — дугообразный верхний компонент позвонка, расположенный над позвоночным каналом и телом позвонка. У взрослых динозавров тело и дуга обычно сливаются вместе через невроцентральный шов, который у молодых особей лежит между этими компонентами. Дуга может содержать целый ряд костных отростков, таких как невральные отростки, эпипофизы, поперечные отростки (которые соединяются с рёбрами) и зигапофизы (которые соединяются с соседними позвонками). У динозавров имелись сложные дуги позвонков, часто украшенные системой пластинок, ямок и / или плевроцелей, которые определяли места залегания воздушные мешков, расположенных вдоль позвонков.
З
Заглазничная кость[12]:65 (лат. os postorbitale, англ. postorbital bone) — парная кость на боковой стороне черепа, которая образует большую часть заднего края глазницы. У цератопсов видоизменена и образует выступающие рога над глазами[19]:38.
Задняя ушная кость[10]:192 (лат. opisthooticum, или paroccipitale) — парная кость задней части черепной коробки. У взрослых динозавров всегда сливается с парными боковыми затылочными костями (), образуя структуру, известную как экзокципитально-опистотические. Вместе с экзокципиталами опистотики образуют пару больших крыльевидных отростков, идущих назад и в стороны от головного мозга, парокципитальных отростков. Эти отростки контактируют с теменными, чешуйчатыми и квадратными отростками
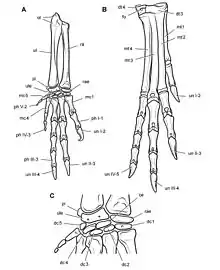
Запястье (англ. carpals) — отдел передней конечности между костями предплечья (лучевой и локтевой) и пястными костями (пястью)[24]. Базальные рептилии показывают 3 ряда костей запястья. У динозавров запястье часто не полностью окостеневает, а количество и идентичность запястных элементов во многих случаях остаются неясными. Особенно хорошо окостенение запястья происходит у базальных птицетазовых гетеродонтозавров, где оно состоит из 9 элементов. Проксимальный ряд состоит из os radiale (ниже лучевой кости), os ulnare (ниже локтевой кости) и гороховидной кости (лат. os pisiforme) — небольшого элемента ниже локтевой кости и латеральнее os ulnare). Дистальный ряд состоит из 5 элементов, расположенных выше 5 мететарсалий, которые обозначены как дистальные запястные кости 1-5. Средний ряд представлен единственным элементом, центральной костью лат. os centrale[37][38].
Затылок (англ. occiput) — область на задней стороне черепа[24]. Он состоит из четырёх костей, окружающих большое затылочное отверстие (лат. foramen magnum) — проход для спинного мозга, который открывается в полость мозга: надзатылочную кость (англ. supraoccipital bone) наверху, парные экзокципитальные кости (англ. exoccipitals) по бокам и базиокципитальную (англ. basioccipital) снизу. Непосредственно под большим затылочным отверстием находится шишковидный отросток — затылочный мыщелок (англ. occipital condyle), который соединяется с атлантом, первым позвонком шеи. Затылочный мыщелок в основном образован базиокципитальной костью, с меньшим вкладом экзокципитальных костей на верхнем крае[18][19]:38.
Затылочная связка (англ. nuchal ligament) — эластичная связка, которая тянется от задней части черепа до задней части шеи над позвоночником[27]:354.
Зигапофизы[12]:157 (лат. zygapophyses) — выступы дуги позвонка, соединяющие соседние позвонки. Каждый позвонок имеет 2 пары зигапофизов, которые сочленяются с зигапофизами следующего и предшествующего позвонка: спереди находятся презигапофизы (лат. praezygapophyses)[10]:195, которые проходят вперёд и имеют суставную поверхность, обращенную вверх. В задней части позвонка находятся постзигапофизы (лат. postzygapophyses)[10]:195, которые отходят назад, и их суставная поверхность обращена вниз[22]:143.
Зуб (лат. dens) — костное образование у животных, во множестве расположенные в ротовой полости. У динозавров зубы присутствуют на верхнечелюстной и предчелюстной костях верхней челюсти и зубной кости (англ. dentary) нижней челюсти[19]:40. Только самые базальные динозавры Eoraptor и Eodromaeus имели рудиментарные зубы на крыловидной кости нёба (птеригоиде) — нёбные зубы. Зуб можно разделить на 2 части: коронку, которая является верхней обнажённой частью, и корень, который находится внутри десны[39]. Зубы в основном состоят из двух типов тканей: эмали и дентина. Эмаль образует твёрдый и блестящий внешний слой, а более мягкий и похожий на кость дентин заполняет внутреннюю часть. Иногда поверх эмалевого слоя можно обнаружить цемент третьего типа[40].

Зубная батарея (англ. dental battery) — тип зубного ряда у некоторых растительноядных динозавров, где отдельные зубы плотно прилегают друг к другу, образуя непрерывную шлифовальную поверхность. У гадрозавридов эдмонтозавров зубная батарея каждой половины верхней и нижней челюстей содержит более 60 рядов зубов, причем каждый ряд включает до трёх функциональных зубов, установленных друг на друга, и до пяти замещающих зубов под этим набором, которые могли прорезаться из челюстей после того, как функциональные зубы стирались и выпадали. Это наиболее сложный тип зубных рядов у динозавров, который развился независимо у гадрозаврид, цератопсов и некоторых завропод и отличается по форме и функциям в этих отдельных группах[19]:185—186[41].
Зубная кость[10]:194 (англ. dentary) — самая крупная кость нижней челюсти. Это единственная нижнечелюстная кость, на которой есть зубы; она расположена впереди всех остальных челюстных костей, кроме птицетазовых, у которых переднюю часть нижней челюсти формирует предзубная кость (премаксиллой, англ. premaxilla). Передние концы зубных костей левой и правой ветвей челюсти соединяются вместе, образуя симфиз нижней челюсти[19]:40.
Зубной ряд (англ. dentition) — собирательный термин для всех зубов, присутствующих в челюстях отдельного динозавра. Зубной ряд может быть гомодонтным, когда присутствует только один тип зубов, или гетеродонтным, в случае разных типов[39]:232-233. Зубы динозавров постоянно менялись в течение жизни. Прорезавшиеся зубы, которые используются в текущий момент, называются функциональными зубами. Для каждого положения зуба обычно есть один или два непрорезавшихся замещающих зуба, которые последовательно мигрируют в зубную альвеолу и заменяют функциональный зуб после того, как последний выпал. Некоторые растительноядные виды могли иметь до шести замещающих зубов на каждое положение зуба. Обновление зубов начиналось с рассасывания корня функционального зуба[42].
Зубные альвеолы, или зубные лунки (англ. dental alveoli) — ямки в челюстях, вмещаюшие корни зубов. В альвеолах зубы динозавров периодически заменялись новыми, растущими под ними; выпадение старой коронки зуба происходило после рассасывания корня. Состояние глубоких альвеол, известное как текодонтия, присутствует у всех архозавров, включая динозавров. У большинства чешуйчатых (лат. Squamata), с другой стороны, наблюдаются такие состояния, как плевродонтия, когда зубы срастаются с внутренними сторонами костей челюсти, или акродонтия зубного ряда, при которой зубы срастаются с верхней частью костей челюсти без лунок. Состояние текодонтии исторически использовалось для определения сборной парафилетической группы текодонтов (лат. Thecodontia), которая в настоящее время считается устаревшей[43].
К
Квадратная кость (лат. os quadratum, англ. quadrate bone) — большая парная эндохондральная кость в задней части черепа. Столбчатая структура, нижний конец которой сочленяется с нижней челюстью, образуя челюстной сустав, а её верхний конец достигает нижней части чешуйчатой кости. Квадратная кость является частью нёба[19]:39—40. Изменение и устойчивость морфологии квадратной кости у некоторых рептилий помогли палеонтологам в таксономии на уровне видов, а также идентификации мозазавров[44] и динозавров из подсемейства Spinosaurinae[45].
Квадратно-скуловая кость[12]:133 (лат. os quadrato-jugale, англ. quadratojugal bone) — кость в щёчной области черепа, которая видна при взгляде сбоку. Образует нижний задний угол черепа[19]:37—38.
Кисть (лат. manus, англ. manus, hand) — дистальная часть передней конечности. Скелет кисти составляют кости запястья, кости пясти и кости пальцев (фаланги)[22]. Первый (большой) палец динозавров в англоязычной терминологии называется pollex[46].
Клиновидно-решётчатая кость, или спленэтмоид[10]:193 (лат. sphenethmoideum) — трубчатая кость в передней части черепной коробки. Расположенная под лобными костями, позади mesethmoideum и перед глазнично-клиновидной костью (орбитосфеноидом), она вмещала обонятельные луковицы — орган обоняния. Как и в случае с mesethmoideum и орбитосфеноидом, сфенэтмоид не окостеневал у многих видов и поэтому редко сохраняется[47][18][19]:39.
Ключица (лат. clavicula, англ. clavicle) — парная изогнутая кость, расположенная выше коракоида в плече. Эти кости прикреплены к акромиону лопатки и лучше всего на скелете видны при виде спереди. Ключицы нечасто встречаются в скелетах динозавров, что, вероятно, связано с их низким потенциалом сохранения[48][19]:49.
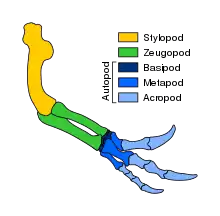
Конечности (англ. limbs) — парные придатки тела у животных, обособленные от туловища или головы, вместе с поясами составляющие часть добавочного скелета. Передние и задние конечности очень похожи по строению и делятся на 3 основные части[22]. Скелет верхней конечности, stylopodium, состоит из плечевой кости (передняя конечность) или бедренной кости (задняя конечность), а нижняя конечность, zeugopodium, состоит из локтевой и лучевой костей (передняя конечность) или большеберцовой и малоберцовой костей (задняя конечность). Дистальная часть — это аутоподия, состоящая из рук или ног. Сама аутоподия подразделяется на 3 сегмента: запястье, или карпалия, (передняя конечность) и предплюсна, или тарзалия (задняя конечность) образуют basipodium; пястье, или метакарпалия, (передняя конечность) и плюсна, или метатарзалия (задняя конечность) образуют metapodium; фаланги пальцев образуют acropodium[49]:18. Альтернативную терминологию предложили Гюнтер Вагнер и Чи-Хуа Чиу в 2001 году. Здесь аутоподиум подразделяется только на две части: мезоподиум, состоящий из узловых элементов (запястья или предплюсны), и акроподиум, который в соответствии с этим определением включает все аутоподиальные длинные кости, включая пястные кости (или плюсну) и фаланги[50].
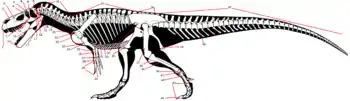
Кости, помимо зубов, являются наиболее распространёнными окаменелыми остатками динозавров. Cостоящие в основном из гидроксиапатита, кости сильно минерализованы и, следовательно, прочны. Мягкие части, напротив, сохраняются толко в виде отпечатков и только в исключительных случаях. Костные части включают череп, осевой скелет (позвоночник и рёбра) и аппендикулярный скелет (плечевой и тазовый пояса и конечности)[51]. Большинство этих костей парные, с зеркально перевёрнутым аналогом на другой стороне тела, или непарные, и в этом случае они обычно лежат на средней линии тела и разделяются ею на две симметричные половины[22]. Кроме того, к костям обычно относят ряд элементов, не связанных со скелетом, и лежащим в/на мягких покровах, такие, как брюшные рёбра (гастралии), костные щитки и шипы[51]. Палеонтологи чаще всего изучают морфологию костей, однако и их гистология (внутренняя микроструктура вплоть до клеточного уровня) и химический состав предоставляют важную информацию о биологии динозавров[52]. Существует 2 основных типа костей: кожная кость образуется непосредственно в коже, обычно вырастая из изначально тонких пластинок. Среди прочего, большинство костей, образующих внешнюю поверхность черепа и нижней челюсти, являются кожными костями. Напротив, эндохондральная кость формируется из хрящевого предшественника, который окостеневает[22].
Крестцовые позвонки (лат. vertebrae sacrales, англ. sacral vertebrates) — часть позвоночного столба, расположенная между поясничными и хвостовыми позвонками. Часто они слиты в структуру под названием крестец[22]:143. Иногда один или несколько спинных позвонков сливаются с крестцом в структуру, известную как сложный крестец (лат. synsacrum)[39]:368.
Крыловидная кость, или птеригоид[10]:193 (англ. pterigoideum) — парная кожная кость нёба. Большая и сложная, она соединяется как с черепной коробкой, так и с другими костями нёба. В своей задней части левый и правый птеригоиды окружают отверстие — межптеригоидную пустоту[19]:39—40.
Крыша черепа — верхняя поверхность черепа. У динозавров она состоит из четырёх парных костей: носовая, лобная и теменная кости являются крупными элементами, а предлобные кости маленькие и расположены между носовой, лобной и слёзной костями. В задней части черепа находятся надвисочные отверстия (англ. supratemporal fenestrae)[19]:38.
Л
Латеросфеноид[16]:301, или боковая клиновидная кость[10]:193 (англ. laterosphenoid), или плевросфеноид (англ. pleurosphenoid) — кость в виде крыла, расположенная в передней части боковой стенки черепной коробки. Снаружи не видна, расположена спереди от проотической кости (англ. prootic bone) и контактирует с теменной, лобной и заглазничной костями, которые расположены сверху. Латеросфеноид как отчётливая оссификация уникальна для архозавриформ; у других рептилий эта часть нейрокраниального хряща не окостеневает[14]:83[19]:39[24][18].
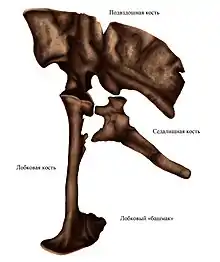
Лобковая кость (лат. os pubis) — одна из трёх парных костей, образующих таз. Расположена вентрально от подвздошной кости и кпереди от седалищной кости. У ящеротазовых ветвь лобковой кости примитивно направлена вперёд (пропубический таз). У птицетазовых исходная лобковая ветвь направлена назад (опистопубический таз); вместо этого имеется хорошо развитая передняя ветвь, или prepubis, которая у производных форм может быть больше, чем ветвь лобковой кости, направленная назад[53]. Левая и правая лобковые кости обычно слиты по всей своей длине, при этом дистальная часть расширяется, образуя т. н. лобковый «башмак», или «фартук»[54]:152. У некоторых представителей группы Neotheropoda дистальная часть лобкового шва образует отверстие — лобковое окно, видимое при взгляде снизу[54]:141. В проксимальной части лобковой кости имеется запирательное[36]:103, или обтураторное[12]:73 (обтюраторное[10]:192) отверстие (лат. foramen obturatum, англ. obturator foramen), которое расположено близко к вертлужной впадине и видно при взгляде сбоку. Это отверстие позволяет пройти запирательному нерву. У теропод это отверстие не полностью окружено костью, так как вентральная граница отсутствует; эта часть лобковой кости называется запирательной впадиной (англ. obturator notch)[54]:143.
Лобная кость (лат. os frontale, англ. frontal bone) — парная кожная кость крыши черепа, которая находится между носовой костью спереди и теменной сзади. Лобные кости, как правило, широкие и образуют верхний край глазницы, а также большую часть крышки черепной коробки[19]:38.
Локтевая кость (лат. ulna) — более крупная и задняя из двух костей предплечья[22]:145. На верхнем конце локтевой кости часто виден большой отросток, простирающийся за локтевой сустав — локтевой отросток (лат. olecranon)[36]:86, к которому прикреплялись трёхглавые мышцы (мышцы-разгибатели предплечья). Иногда значительное удлинение локтевого отростка давало этим мышцам повышенную нагрузку[19]:50[4]:173-174.
Лопатка (лат. scapula, англ. blade) — эндохондральная кость и самый большой элемент плечевого пояса. У взрослых особей лопатка обычно сливается с коракоидом, образуя скапулокоракоид[10]:197 (лат. scapulocoracoideum)[3], на котором имеется сочленовная ямка для плечевой кости — гленоид (лат. glenoideum)[10]:197.
Лучевая кость (лат. radius, англ. radius) — парная кость в составе предплечья, расположенная рядом с более крупной локтевой костью[22]:145.
М
Малоберцовая кость (лат. fibula) — длинная кость, расположенная латеральнее большеберцовой кости; вместе две кости образуют голень. Обычно более тонкая, чем большеберцовая кость, и особенно тонкая у производных теропод и птиц[22]:147[19]:53.
Медуллярная кость[21]:99—100 (англ. medullary bone) — особый тип костной ткани у птичьих и нептичьих динозавров, включая птиц, который образуется одновременно с производством яиц. Медуллярная кость образуется в особой полости в костях ног и необходима животным для восполнения кальция в организме во время откладывания яиц. Широкое распространение этого типа костной ткани у разных групп (она найдена у тероподов аллозавра и тираннозавра, у птицетазового тенонтозавра и завроподоморфа мусзавра) указывает на её присутствие у всех линий динозавров. Медуллярная кость имеет важное значение для определения пола животного, остатки которого послужили основой для окаменевшего экземпляра: её присутствие однозначно указывает на женский пол, однако её отсутствие может указывать как на самца, так и на самку, не находившуюся на момент смерти в состоянии откладывания яиц[21]:99—100.
Межключица (лат. interclavicle), или надгрудинник (лат. episternum)[10]:197 — непарная стержнеобразная кожная кость, расположенная между ключицами в плечевом поясе[48]. Примитивная особенность четвероногих; как правило, считается, что этот элемент скелета отсутствовал у динозавров, поскольку был утрачен ещё у динозавриформов[55]. Тем не менее наличие межключицы у некоторых таксонов динозавров высказывалось неоднократно[48].
Межпластинчатая эластическая связка (лат. ligamentum elasticum interlaminare) — связка, состоящая из непарных пучков, которые соединяют дуги соседних позвонков у динозавров и птиц[56]. Эта связка охватывает шею и хвост, и у современных птиц поддерживает вогнутость шеи[56][57]. Было высказано предположение, что распад этой связки во время разложения тела приводит к характерной позе смерти, когда позвоночник сильно прогибается[56].
Меккелева борозда[58] (англ. Meckelian groove) — продольный канал в нижней челюсти. В нём находились неокостеневшие остатки меккелева хряща, из которых окостеневали эндохондральные элементы нижней челюсти. Передняя часть мекелевой борозды обнажается на медиальной стороне нижнечелюстной кости рядом с её вентральным краем, а задняя часть окружена костью с обеих сторон, открываясь лишь дорсально через приводящую ямку[59][13]:44.
Мозжечок (лат. cerebellum) — дорсальная часть задней части мозга между стволом головного мозга и головным мозгом, которая служит для контроля баланса, позы и движения. Эту часть мозга обычно нельзя увидеть в окаменелостях динозавров, поскольку она редко встречается в эндокастах; исключением является возможное сохранение листков мозжечка у Conchoraptor[24][60][61].
Н
Надугловая кость[62] (лат. os surangulare / os suprangulare, англ. surangular / supraangular) — кожная кость в нижней челюсти, которая видна при взгляде сбоку, и находилась за зубной костью и над угловой костью[19]:40.
Наружное нижнечелюстное окно (англ. external mandibular fenestra) — отверстие в нижней челюсти между зубной, надугловой и угловой костями. Оно характерно для архозавров и присутствует у большинства групп динозавров[19]:40.
Невроцентральный шов (англ. neurocentral suture) — разделение между телом и дугой каждого позвонка. У молодых динозавров обычно присутствуют «открытые» (несращённые) нейроцентральные швы, которые становятся «закрытыми» (слитыми) по мере достижения ими взрослого возраста.
Нёбная кость (лат. os palatinum, англ. palatine) — парная кожная кость нёба. Контактирует с сошником и крыловидным отростком медиально, а также с верхней челюстью и скуловой костью латерально и образует задний край хоан, а также передний край подглазничных отверстий[19]:39—40.
Нёбо (англ. palate) — часть черепа между левой и правой верхнечелюстными костями, которая образует свод рта. Оно состоит из одной непарной кости — сошника — и пяти парных костей (нёбной, крыловидной, эктоптеригоидной, эпиптеригоидной и квадратной). Нёбо имеет 3 пары отверстий, а именно: хоаны, или внутренние ноздри, подглазничные отверстия между нёбными костями и эктоптеригоидами и подвисочные отверстия за эктоптеригоидами, которые переходят в подвисочные окна[19]:39—40.
Нижняя челюсть (лат. mandibula, англ. mandible) — непарная структура, образующая нижний отдел черепа. Каждая половина нижней челюсти, называемая нижнечелюстной ветвью (лат. ramus mandibulae)[36]:63, соединена с квадратной костью в задней части черепа, образуя челюстной сустав. Спереди обе нижнечелюстные ветви прикреплены друг к другу; область прикрепления называется нижнечелюстным симфизом. Нижняя челюсть состоит как из эндохондральных костей, окостеневших из меккелевого хряща, так и из кожных костей[59]. У динозавров только зубная кость (лат. os dentale, англ. dentary) несёт зубы[19]:40.
Носовая кость (лат. os nasale, англ. nasal) — самая передняя парная кость крыши черепа и самая большая кость верхней поверхности рыла. Расположена между предчелюстной и лобной костями[19]:38[22]:141.
О
Окостеневшие сухожилия — костные стержни, укрепляющие позвоночник[22]:148. Их присутствие в верхней части позвонков считается синапоморфией птицетазовых динозавров[63].
Орбитосфеноид[16]:301, или глазнично-клиновидная кость[10]:193 (англ. orbitosphenoid) — парная кость в передней части черепной коробки, которая подстилает переднюю часть головного мозга. Левая и правая половины закрывают канал для зрительного нерва. У динозавров окостеневает лишь изредка[18][19]:39[47].
Основная затылочная кость[10]:192 (англ. basioccipitale, англ. basioccipital) — непарная кость черепа и одна из четырёх костей, образующих затылочную часть черепной коробки. Она видна сзади на сочленённом черепе. Образует бо́льшую часть затылочного мыщелка — округлого отростка, соединяющего череп с первым шейным позвонком. Вверху срастается с парными exoccipitals. Также образует заднюю часть дна черепной коробки, сливаясь с базисфеноидом в его передней части[18].
Остеодермы — окостенения, располагающиеся в мезодермальном слое кожи. У некоторых видов они могут образовывать пластинчатые или шиповидные структуры, особенно у бронированных анкилозавров. Маленькие и неправильной формы остеодермы диаметром менее 1 см (или 5-15 мм[64]) в англоязычных источниках известны как ossicles[65][66].
Отверстие (лат. foramen, ед. ч.; мн. ч. лат. foramina) — отверстие в кости, через которое проходят кровеносные сосуды, нервы, мышцы и тому подобное[24]. Питательные отверстия — это отверстия, позволяющие кровеносным сосудам проникать внутрь костей, чтобы обеспечить доставку питательных веществ к кости[19]:224.
Отростки позвонка[36]:18 (лат. processus vertebrale, англ. neural spines) — отростки, поднимающиеся вверх от верхней поверхности дуги позвонка. Образуют единый ряд по средней линии позвоночника, причём их кончики обычно являются высшими точками отдельных позвонков. Обеспечивают поверхности прикрепления для мышц и связок, идущих поверх позвоночника[67]:8. У некоторых видов они могут быть значительно удлинены и образовать парус, или глубоко раздвоены, так что их верхние части образуют двойной ряд[68][69].
П
Пальцы (англ. digits) — дистальная часть аутоподии, следующая за пястью передней конечности и плюсной задней конечности. Обычно обозначаются римскими цифрами от I до V, где I обозначает самый внутренний палец, а V — самый наружный. Каждый палец состоит из одной или нескольких фаланг[22]:145.
Парапофиз[10]:195 (англ. parapophysis) — каждый из нескольких отростков в телах шейных и спинных позвонков, которые сочленяются с головками рёбер. Обычно они располагаются близко к переднему краю боковой поверхности тела позвонка и хорошо отделяются от диапофизов на поперечных отростках дуг, которые принимают бугорок ребра (лат. tuberculum costae). Однако положение парапофиза постепенно смещается на поперечный отросток вдоль позвоночника от шейных к задним спинным позвонкам, где он может образовывать единую структуру с диапофизом, который соединяется с одноглавым ребром[11]:225[67]:8.
Парасфеноид[1]:174 (лат. parasphenoideum, англ. parasphenoid) — единственная покровная кость черепной коробки. Является элементом нижней части черепной коробки, находится перед базисфеноидом и вытянутым отростком простирается вперёд под глазами. Этот отросток поддерживал мембрану, которая простиралась дорсально до крыши черепа, разделяя правое и левое глазные яблоки. Эта мембрана может окостенеть, и тогда её называют межглазничной перегородкой[10]:191 (англ. interorbital septum)[19]:39.
«Парус» — структура, возникающая в результате удлинения невральных отростков позвонков, обычно грудных, крестцовых и хвостовых. Паруса различаются по размеру и форме, от мелких гребней до высоких заметных структур[68][70]. В некоторых случаях, например, у Concavenator, невральные отростки образуют «горб»[71], а у Ichthyovenator — синусоидальный (волнообразный) парус[72]. Это состояние наиболее заметно у спинозаврид[70], но также встречается у кархародонтозаврид, Metriacanthosauridae[68][73], и нескольких видов орнитопод и завропод[74][75][76].
Передняя ушная кость[10]:192 (лат. prooticum, англ. prootic) — парная кость, образующая части боковой поверхности черепной коробки. Вместе с задней ушной костью (лат. opisthoticum), расположенной позади неё, и верхней ушной костью (лат. epioticum), расположенной выше, образует слуховую капсулу. Из них вторая срастается с боковой затылочной костью (лат. exoccipitale), а третья — с верхней затылочной (лат. supraoccipitale)[10]:192. Передняя ушная кость содержит отверстия для тройничного и лицевого нервов и входит внутрь пазух. Её внешняя поверхность обеспечивала грубые поверхности для прикрепления мышц, открывавших челюсть[19]:39.

Пигостиль (лат. pygostylus, urostylus, coccyx[77], англ. pygostyle) — костная структура, состоящая из сросшихся задних хвостовых позвонков. Обнаруживается у пигостилиевых птиц, где выполняет функцию закрепления длинных рулевых перьев[24]. Помимо птиц, пигостиль был у некоторых меловых динозавров, например, у Nomingia[78].
Пинеальное отверстие[12]:341 (англ. pineal foramen) (также: лобно-теменное отверстие; теменное отверстие; постфронтальное отверстие; межфронтальное отверстие) — непарное отверстие на средней линии крыши черепа, присутствующее у некоторых динозавров, но часто закрытое у взрослых особей. Его положение изменчиво; оно может располагаться между лобными, теменными костями или на стыке лобных и теменных костей. В этом отверстии располагается теменной глаз некоторых бесчелюстных, рыб, земноводных и рептилий. В окаменелых черепах многих вымерших позвоночных пинеальное отверстие в некоторых случаях больше, чем у любого живого позвоночного животного[79][80][81].
Пластинка (лат. lamina), или костная перегородка (лат. osseus septum) — гребень в позвонке, разделяющий пневматические камеры. Пластинки берут начало и заканчиваются на разных отростках и областях позвонка; от начальных и конечных точек прикрепления зависит их предназначение и названия. Пластинки делятся на 4 произвольные категории в зависимости от их конечных точек прикрепления: диапофизные пластинки, парапофизные пластинки, зигапофизные пластинки и позвоночные пластинки. Они также могут быть разделены на субпластинки, если они расщепляются по длине, или на сложные пластинки, если более одной пластинки образуют гребень[82].
Пластинчатая кость[10]:194 (лат. os spleniale, англ. splenial bone) — кожная кость нижней челюсти, которая расположена позади и медиальнее (по направлению к внутренней части) зубной кости (лат. os dentale). Пластинчатая кость покрывает бо́льшую часть внутренней стороны зубной кости и часто является самой обширной костью нижней челюсти на медиальной проекции[19]:40[59].
Плевроцели[12]:71 (англ. pleurocoels) — отверстия на боковых поверхностях позвонка, которые ведут во внутренние камеры в теле и / или дуге позвонка. Плевроцель может быть отдельной полостью или комплексом более мелких взаимосвязанных полостей[22]:143-144.
Плечевая кость (лат. humerus) — единственная кость плеча и самая большая кость передней конечности[22]:145.
Плечевой пояс (англ. pectoral girdle) — набор костей аппендикулярного скелета, который соединяется с передними конечностями. Строго говоря, грудной пояс включает лопатку, коракоид и ключицу, тогда как грудину по-разному относят либо к грудномуплечевому поясу, либо вместе с рёбрами к осевому скелету. В отличие от тазового пояса, который жёстко закреплён на позвоночнике, грудной пояс не имеет такого костного соединения, а вместо этого укреплён мышцами[3].
Плюсневые кости (лат. ossa metatarsi), или метатарзалии (англ. metatarsals) — длинные кости стопы. Вместе называемые плюсной, они соединяют предплюсну с пальцами. Обозначаются римскими цифрами от I до V, где I плюсневая кость является самой внутренней (соединяется с большим пальцем стопы), а плюсневая кость V — самой внешней[22]:147-148.
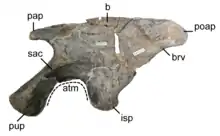
Подвздошная кость (лат. os ilium) — парная кость, наиболее крупная из трёх костей, формирующих тазовую кость. Посредством подвздошной кости тазовая кость сочленяется с позвоночником. Она состоит из основного тела — подвздошной лопасти, медиальная сторона которой прикрепляется к крестцовым позвонкам. Вентрально она образует утолщённый изогнутый гребень, нависающий над вертлужной впадиной, называемый надацетабулярным гребнем (англ. supraacetabular ridge), который распределяет нагрузку, передаваемую через бедренную кость. Переднее расширение подвздошной лопасти называется преацетабулярным отростком (англ. preacetabular process), а заднее расширение — постацетабулярным отростком (англ. postacetabular process)[83]; оба расширения позволяют оптимально разместить основные мышцы конечностей. Перед вертлужной впадиной находится толстый отросток — лобковая ножка (англ. pubic peduncle), которая, расширяясь вентрально, прикрепляется к лобковой кости, в то время как седалищная ножка (англ. ischiadic peduncle) простирается за вертлужной впадиной, прикрепляясь к седалищной кости[84]:33. Сразу за вертлужной впадиной на седалищной кости располагается противовертел (англ. antitrochanter), суставная поверхность которого простирается вентрально на седалищную кость и сочленяется с большим вертелом бедренной кости[19]:19[85]:726. Вентральная поверхность постацетабулярного отростка несёт углубление, называемое короткая ямка лат. brevis fossa, где берёт начало короткая хвосто-бедренная мышца лат. m. caudofemoralis brevis[84]:21. Латерально с короткой ямкой граничит короткая полка (англ. brevis shelf) — гребень, идущий вдоль вентролатерального края подвздошной кости между седалищной ножкой и задним концом кости[85]:733-734.
Подвисочное отверстие (англ. infratemporal fenestra, также: латеральное височное отверстие (англ. lateral temporal fenestra)) — главное отверстие на боковой стороне черепа, расположенное за глазницей[22]:140. Оно ограничено заглазничной костью в верхней передней части, чешуйчатой — в верхней задней части, квадратно-скуловой — в нижней задней части, и скуловой костью — в нижнем переднем углу[19]:37.
Подноздревая щель (англ. subnarial gap) — щель между предчелюстной и верхнечелюстной костями в мордах целофизид, дилофозаврид и спинозаврид. Подноздревая щель является причиной диастемы — разрыва в зубном ряду. Пространство между подноздревая щелью рыла и изогнутым вверх концом нижней челюсти называется подноздревой выемкой[86][87][88]. Подноздревая щель дилофозавра содержала глубокую выемку позади зубного ряда предчелюстной кости, называемую подноздревой ямкой[86].
Подноздревое окно (англ. subnarial foramen) — небольшое отверстие между предчелюстной и верхнечелюстной костями у ящеротазовых и некоторых базальных птицетазовых динозавров[19]:38.
Подушечка (англ. pad) — мясистая припухлость на внутренней стороне передней или задней конечности, которая у динозавров обычно наблюдают на следах. В зависимости от той части конечности, которую они поддерживают, их описывают как подушечки кисти, подушечки стопы или подушечки пальцев[5]:47. Палец может иметь отдельные подушечки фаланг (в англоязычной терминологии называемые также узлами пальцев, англ. digital nodes[89]), которые могут соответствовать отдельным фалангам[90]. У динозавров подушечки фаланг обычно имеют суставное расположение, при этом подушечка охватывает сустав между двумя фалангами. Люди, с другой стороны, демонстрируют мезартральное расположение, когда переход от одной подушечки к другой происходит под суставом. Подушечка, охватывающая сустав между пястной или плюсневой костями и первой фалангой, называется мететарсо-фаланговой подушечкой англ. metatarsophalangeal pad[89].
Подъязычная кость (лат. os hyoideum, англ. hyoid) — парная кость горловой области, расположенная ниже и позади черепа, которая поддерживает язык. Эти элементы редко встречаются у динозавров, поскольку они не сочленяются с остальными элементами скелета и поэтому легко теряются[4]:155.
.jpg.webp)
Поза смерти (англ. death pose), или опистотоническая поза (англ. opisthotonic posture) — обычная поза, наблюдаемая в сочленённых окаменелостях динозавров и птиц, где позвоночник сильно изогнут вверх (чрезмерно вытянут)[91]. Обсуждаются различные возможные причины, в том числе опистотонус (предсмертная агония)[91] и сокращение связки ligamentum elasticum interlaminare во время разложения[56].
Позвонок (лат. vertebra) — костный компонент позвоночного столба. Позвонки динозавров разделяют на шейные, спинные, крестцовые и хвостовые. Каждый позвонок имеет центральный компонент в форме катушки — тело, и верхний компонент дугообразной формы — невральный отросток, или неврапофиз (лат. neurapophys)[10]:195. Между этими компонентами проходит туннель в форме трубки — позвоночный канал. Через позвоночный канал каждого позвонка в позвоночнике проходит Спинной мозг нервной системы. Шейные, спинные и крестцовые позвонки соединяются с рёбрами посредством поперечных отростков по бокам их дуг. Хвостовые позвонки соединяются с шевронами на нижней стороне их тел[36]:18. Позвонки у пресмыкающихся сочленяются при помощи передних сочленовных отростков, или презигапофизов (лат. praezygapophyses), и задних сочленовных отростков, или постзигапофизов (лат. postzygapophyses)[10]:195.
Поперечный затылочный гребень (лат. crista occipitalis transversa, англ. transverse nuchal crest) — костный гребень в задней части черепа, встречающийся у многих теропод. Направленный поперечно (перпендикулярно средней линии черепа), он образует границу между крышей черепа и затылочной областью. В основном образован задним краем теменных костей с участием чешуйчатых костей. Гребень фиксирует мускулатуру шеи, а его задняя поверхность может содержать ямки для прикрепления затылочной связки. Поперечный затылочный гребень не следует путать с сагиттальным затылочным гребнем — срединным гребнем надзатылочной кости (англ. supraoccipital bone)[30]:82[92]:117[93]:167.
Поперечный отросток (англ. transverse process) — крыловидный выступ, отходящий в сторону от невральных дуг шейных, спинных, крестцовых и передних хвостовых позвонков. Их типичная горизонтальная ориентация приводит к характерной Т-образной форме позвонков при взгляде спереди или сзади. Своим концом поперечный отросток сочленяется с бугорком соответствующего ребра; расширенная область сочленения ребра называется диапофизом[10]:195 (лат. diapophysis, ед. ч.; мн. ч. — лат. diapophyses)[19]:38. Некоторые авторы, однако, называют весь поперечный отросток диапофизом, используя оба термина как синонимы[11]:224-225[67]:8.
Посткраниальный скелет[10]:195 (англ. postcranium, от лат. post cranium — «позади черепа») — часть скелета, которая следует за черепом. Он подразделяется на осевой скелет, который включает в себя все позвонки и рёбра, и аппендикулярный скелет, который включает пояса и конечности[22].
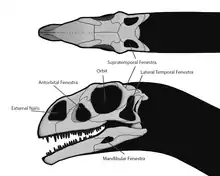
Предглазничное окно, или предглазничное отверстие[12]:163 (англ. antorbital fenestra) — отверстие в черепе, которое располагается перед глазницами. Данная особенность черепа во многом связана с архозаврами и впервые появилась в триасовом периоде. Среди сохранившихся архозавров предглазничное окно сохранилось у птиц, тогда как крокодилы утеряли его[94][95]. У некоторых видов архозавров окно закрылось, но его местоположение всё ещё отмечено впадиной, или ямкой, на поверхности черепа, называемой предглазничной ямкой[12]:163. В предглазничном окне располагаются придаточные пазухи, которые сливаются с соседней носовой капсулой[96].
Предзубная кость[10]:194; иногда транслитерируется как предентале[12]:159 (лат. os predentale, англ. predentary) — непарная кость, находимая перед нижнечелюстной костью, которая образовывала нижнюю часть клюва у птицетазовых динозавров[22]:142. Её наличие считается синапоморфией птицетазовых[97].
Предлобная кость[10]:194 (лат. prefrontalia, англ. prefrontal bone) — маленькая кость на боковом крае крыши черепа между лобной, слёзной и носовой костями[19]:38.
Предплюсна, или тарсалии[12]:71 (лат. tarsus) — набор костей задней конечности между плюсневыми костями снизу и большеберцовой и малоберцовой костями сверху[22]:148. Предплюсна состоит из верхнего ряда, образованного таранной костью, или астрагалом (лат. astragalus), и пяточной костью, и нижнего ряда мелких костей, расположенных поверх плюсневых. Верхний и нижний ряды могут двигаться друг против друга, образуя голеностопный сустав[19]:53.
Предсочленовная кость[10]:194 (лат. os prearticulare, англ. prearticular bone) — кость задней части нижней челюсти. Расположена на внутренней стороне последней, находится перед сочленовной костью и над угловой костью. Образует внутренний край приводящей ямки (англ. adductor fossa)[59].
Предчелюстная кость (лат. premaxilla) — самая передняя кость верхней челюсти и образует кончик рыла. Содержит переднюю часть верхнего зубного ряда, который продолжается на верхней челюсти, идущей следом[19]:36.
Приводящая ямка, или меккелево отверстие (лат. adductor fossa, англ. Meckelian orifice) — большое отверстие в нижней челюсти рептилий и динозавров, расположенное между областью, несущей зубы, и суставом челюсти. Она открывается дорсально и окружена латерально стенками надугловой кости, а медиально — стенками предсуставной кости; поскольку последняя обычно намного ниже, чем первая, ямка видна медиально. Дно отверстия образовано угловой костью и вмещает заднюю часть меккелева хряща. Приводящая ямка и окружающие её края служат точкой прикрепления основных приводящих мышц, закрывающих челюсть; она также позволяет главному нерву, артерии и вене челюсти проникать внутрь нижней челюсти[59].
Проатлас[10]:196 (лат. proatlas) — небольшая парная кость, расположенная между дугой атланта и основанием черепа. Элементы каждой пары имеют треугольную форму, причём широкий конец соединяется с дугой атланта, а острый конец — с боковой затылочной костью (лат. exoccipitale) черепа. Учитывая его небольшой размер, proatlas легко теряется во время фоссилизации, сбора или сохранения окаменелостей и поэтому встречается редко[11].
Пястные кости (лат. ossa metacarpi), или метакарпалии (англ. metacarpals) — длинные кости передней конечности, образующие скелет кисти. Расположены между запястьем и фалангами пальцев. Обозначаются римскими цифрами от I до V, где I пястная кость — самая внутренняя (соединяется с большим пальцем), а V — самая внешняя[22]:145.
Пятка — выпуклая часть тела на задней стороне ступни. Состоит из пяточной кости, расположенных на ней жировых тканей и кожи. Этот термин также относится к задней части следа, и в этом случае он не обязательно соотносится с анатомической пяткой, которая у многих динозавров была приподнята над землёй[5]:47.
Пяточная кость (лат. os calcaneum) — основная кость голеностопа, раположенная в задней части стопы и вместе с таранной костью (лат. astragalus) образующая верхний ряд костей предплюсны. Расположена латеральнее таранной и дистальнее малоберцовой кости[98].
Р
Рамфотека — роговой (ороговевший) покров на кончиках челюстей, обычно встречающийся у птицетазовых, орнитомимозавров и птиц[24][51]. Поскольку кератин фоссилизируется редко, рамфотека сохраняется только в виде отпечатков. На её наличие указывает шероховатая поверхность кончиков челюстей, которые сильно васкуляризированы (содержат множество кровеносных сосудов)[2].
Ребро — каждая из парных дугообразных плоских костей, идущих от позвоночника к грудине и составляющих грудную клетку. Рёбра прикрепляются к обеим сторонам спинных позвонков и обеспечивают защиту органов, расположенных в грудной полости[36]:26. Каждое ребро имеет 2 головки[99]: дорсальная головка (лат. tuberculum) сочленяется с поперечными отростками невральной дуги позвонка, а вентральная головка, (лат. capitulum) сочленяется с телом позвонка. Рёбра также присутствуют на шее (шейные рёбра) и крестце (крестцовые рёбра)[22].
Рог — вырост костей черепа, покрытый ороговевшей оболочкой. Поскольку ороговевшая оболочка, как правило, не сохраняется в окаменелостях, её присутствие у ряда динозавров было выведено из сравнений с современными полорогими. Другие черепные выросты или складки могут быть покрыты кожными структурами, как было предложено, например, для носового бугра пахиринозавра и куполов пахицефалозавров[100].
Родничок (лат. fonticulus, англ. fontanelle) — промежутки между костями, обычно костями черепа, которые в основном присутствуют у эмбрионов, но часто закрываются по мере роста животного[101]:25. Черепа цератопсов содержат отверстия между лобными и теменными костями, называемые лобными родничками (англ. frontal fontanelles), которые представляют собой отверстия надчерепной полости — особенность, уникальная для этой группы[102].
Ростральная кость[12]:159 (англ. rostral bone) — особая кость, находимая у цератопсов. Непарный элемент, покрывающий переднюю часть предчелюстной кости на кончике морды[19]:36. Ростральные и другие дополнительные окостенения, прикреплённые к черепам цератопсов собирательно называются эпиоссификациями (англ. epiossifications)[103].
Рострум (лат. rostrum) — направленный вперёд длинный отросток на кончике морды, образованный сращенными парасфеноидом (лат. parasphenoideum) и базисфеноидом лат. basisphenoideum[10]:194.
С
Седалищная кость (англ. ischium) — одна из трёх парных костей таза, расположенная ниже подвздошной кости и позади лобковой кости. Это стержневой элемент с расширенной «полкой» на проксимальном конце, запирательная пластина которой контактирует с подвздошной и лобковой костями (соответствующая запирательная пластина находится также в лобке). Как и лобковая кость, левая и правая седалищные кости сливаются по всей своей длине. Расширенный симфиз каудовентрально направленного вертела называется седалищным фартуком (англ. ischiadic apron)[54]:139. Дистальный конец седалищной кости часто расширяется в т. н. седалищный башмак (англ. ischial boot). На вентральной поверхности вертела имелся продольный гребень — запирательный отросток (англ. obturator process). У представителей базальных таксонов запирательный отросток расположен на проксимальном конце седалищной кости, а у целурозавров — на дистальном[54]:152.

Склеротикальное кольцо[10]:590 (лат. anulus ossicularis sclerae, англ. sclerotic ring) — кольцо из небольших пластинчатых костей, расположенных на глазу внутри склеры и вокруг зрачка. Отдельные кости склеротикального кольца называются склеральными косточками (англ. scleral ossicles)[104][105][19]:84.
Скуловая кость (лат. os jugale, англ. jugal bone) — кость черепа, которая определяет нижнюю границу глазницы и соединяется с верхней челюстью спереди и с квадратно-скуловой костью сзади. Её задний край раздваивается в месте сочленения с квадратно-скуловой костью, что считается синапоморфией динозавров[22]:140[19]:12.
Слёзная кость (англ. os lacrimale, англ. lacrimal bone) — кость черепа в форме стойки, отделяющая предглазничное окно от глазницы. В ней проходит канал слёзного протока, который соединяет глазницу с предглазничным окном. У некоторых видов верхний край слёзной кости имеет треугольный гребень или отросток, который возвышается над крышей черепа[19]:37[14]:62.
Слуховая капсула (англ. otic capsule) — утолщённая боковая часть черепной коробки, в которой находится внутреннее ухо. Она состоит из двух окостенений: проотического спереди и опистотического сзади. Между обеими костями лежит овальное окно (лат. fenestra ovalis) — большое отверстие, вмещающее стремечко[14].
Сочленение (лат. articulatio, англ. articulation) — подвижное соединение костей скелета. Термин также используется для описания сохранения окаменелых образцов: в сочленённом образце отдельные кости слиты в своём первоначальном анатомическом положении. И наоборот, кости несочленённого скелета выходят за пределы их первоначального анатомического строения. Образец, обнаруженный с несочленёнными костями, но в непосредственной близости друг от друга, называется связанным скелетом. Переход от полностью сочленённого скелета к кластеру изолированных и несочленённых костей происходит плавно[106].
Сочленовная кость[12]:340—342 (лат. os articulare, англ. articular bone) — самая задняя кость нижней челюсти. На своей дорсальной поверхности он содержит гленоид (англ. glenoid) — углубление, в которое входит нижний конец квадратной кости черепа, образуя челюстной сустав. Это единственная эндохондральная кость нижней челюсти, образующаяся непосредственно из задней части меккелева хряща. У млекопитающих этот суставной элемент переместился в череп, образуя молоточек среднего уха[59].
Сошник (лат. vomer) — непарная узкая кость передней части нёба, разделяющая левую и правую верхнечелюстные кости и хоаны[19]:39—40.
Спинные позвонки (англ. dorsals, англ. dorsal vertebrae, back vertebrae) — позвонки в области спины, между шейным и крестцовым отделами позвоночника. В терминологии Nomina Anatomica Avium (NAA) вместо этого термина используется термин «грудные позвонки» (англ. thoracic vertebrae). У млекопитающих термин «грудные позвонки» относится только к грудным позвонкам, несущим рёбра, тогда как позвонки нижней части спины, на которых рёбра отсутствуют, называются поясничными позвонками. Такое разделение отделов позвоночника не применимо для динозавров[22]:143, поскольку у многих из них поясничный отдел выражен нечётко, и в этом случае грудные и поясничные позвонки называются спинными[10]:196. У большинства динозавров было около 15 спинных позвонков. По сравнению с шейными, спинные позвонки короче, их дуги выше, а поперечные отростки более крепкие и направлены наружу[19]:45.
Стопа (лат. pes) — дистальная часть нижней конечности. Состоит из костей предплюсны, плюсневых костей и костей пальцев (фаланг)[36]:115. Первый палец стопы короткий и у большинства бипедальных динозавров не касался земли, а у птиц увеличен и развёрнут (направлен назад, а не вперёд)[2][107].
Т
Таз (лат. pelvis) — расположенная в основании позвоночника часть скелета, обеспечивающая прикрепление к туловищу задних конечностей, а также являющаяся опорой и костным вместилищем для ряда жизненно важных органов[36]:103—104. Таз состоит из подвздошной, лобковой и седалищной костей. Его внутренняя поверхность имеет прочное соединение с крестцовыми рёбрами крестцовых позвонков, которые могут срастаться в крестец[3].
Таранная кость (лат. astragalus; иногда встречается термин «астрагал»[12]:340—342) — одна из костей предплюсны, формирующая нижнюю часть голеностопного сустава посредством соединения с больше- и малоберцовой костями. Таранная кость не вращается относительно пяточной кости или большеберцовой кости; вместо этого у двуногих она часто сливается с обоими элементами. Тогда такой элемент скелета называется тибиотарзус (лат. tibiotarsus)[98].
Тело позвонка (лат. centrum) — элемент в виде короткого цилиндра, который вместе с дугой (лат. arcus vertebrae, англ. neural arch) образует позвонок[22]. У молодых особей и тело, и дуга являются отдельными элементами; слияние этих частей является важным остеологическим показателем зрелости. Передняя и задняя поверхности тела позвонка образуют сочленение с телами предыдущего и следующего позвонков. Форму тел позвонков можно классифицировать на основе морфологии их суставных поверхностей:
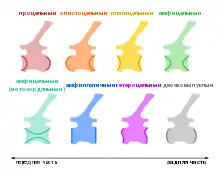
- амфицельные (англ. amphicoelous) тела сильно вогнуты с концов;
- платицельные (англ. platycoelous) тела слегка вогнуты с концов;
- амфиплатичные (англ. amphiplatian), или ацельные (англ. acoelous) тела плоские с обеих концов;
- процельные (англ. procoelous) тела спереди вогнутые, а сзади выпуклые;
- опистоцельные (англ. opisthocoelous) тела вогнутые сзади и выпуклые спереди;
- гетероцельные (англ. heterocoelous) тела имеют седловидную форму с обоих концов.
Амфицельная форма свойственна для тел позвонков примитивных тетрапод. У рыб концы тел глубоко вогнутые и соединяются через небольшое отверстие — проход для хорды. У рептилий этот тип тел позвонков присутствует у эмбрионов и у взрослых форм некоторых видов. У большинства видов, включая динозавров, тела позвонков сильнее окостеневают при закрытом хордовом отверстии, что улучшает сопротивление силам сжатия. Гетероцельные тела позвонков обеспечивают позвоночнику гибкость, предотвращая скручивание. Процельные и опистоцельные тела образуют вогнуто-выпуклые суставы, где выпуклые концы образуют мыщелок (англ. condyle), входящий в вогнутую часть — впадину (англ. cotyle). Такая конфигурация обеспечивает большую стабильность без ограничения мобильности. Подобная стабилизация лучше всего работает при длинных шее и хвосте, когда выпуклая часть направлена в сторону от тела. Позвонки завропод перед крестцом обычно опистоцельные, а хвостовые позвонки — процельные. Поскольку позвоночный столб может содержать различные морфологические типы тел позвонков, могут возникать переходные тела позвонков с двумя разными концами[11][108][109].
Теменная кость (лат. os parietale, англ. parietal) — парная кость, образующая боковую и верхнюю части крыши черепа. Расположена позади лобных костей[36]:39.
Теменные отверстия[10]:193 (англ. foramen parietale, англ. parietal fenestrae) — парные окна, находимые в «воротниках» цератопсов, образованных теменными костями. У Chasmosaurus и Pentaceratops эти отверстия были особенно большими[110].
У
Угловая кость[12]:340—342 (англ. angular bone) — крупная кость нижней челюсти. При виде сбоку она покрывает большую площадь задневентральной области челюсти, находясь позади зубной кости (англ. dentary) и ниже надугловой кости (англ. surangular). При медиальном виде нижняя челюсть видна ниже предсуставной кости (англ. prearticular). Она образует дно приводящей ямки и поддерживает заднюю часть меккелева хряща[59].
Х
Хвостовая корзина (англ. caudal basket) — сетчатая структура, находимая в хвосте пахицефалозавров. Хвостовая корзина окружает позвоночный столб (но не соединяется с ним) и состоит из костных элементов, каждый из которых в англоязычной терминологии называется myorhabdoi. Эти кости образуются внутри миосепты (соединительной ткани, разделяющие блоки мышечной ткани, миомеры) и представляют собой окостеневшие мышечные сухожилия. Следовательно, myorhabdoi образуются на периферии мышечной ткани ниже дермы (а не внутри неё, как в случае остеодермы). Они также отличаются от окостеневших сухожилий, которые укрепляли хвост у других птицетазовых динозавров, хотя, возможно, они имели схожие функции. Myorhabdoi в основном известны по костистым рыбам; пахицефалозавры — единственные известные четвероногие, у которых эти структуры окостенели. Сочленённая хвостовая корзина известна у образца Homalocephale[111].
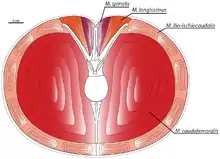
Хвостово-бедренная мышца[21]:104 (лат. musculus caudofemoralis) — парная мышца, которая является основной двигательной мышцей у всех длиннохвостых динозавров. Расположенные в основном в хвосте, они тянут бедро задней конечности назад при сокращении, обеспечивая таким образом толчок. У длиннохвостых динозавров m. caudofemoralis исключительно большая; её массу можно оценить в 58 % от общей массы хвоста тираннозавра. У динозавров выражены две части: m. caudofemoralis brevis берет начало на нижнем крае задней части подвздошной кости, а лат. musculus caudofemoralis longus берет начало в передней части хвоста, вентральнее поперечных отростков и ниже гипаксиальной мускулатуры хвоста. Каждая часть крепится к четвёртому трохантеру на тыльной стороне бедренной кости. Хвостово-бедренные мышцы теропод редуцировались в ходе эволюции, и у современных птиц они в основном утрачены. Птицы, в отличие от длиннохвостых динозавров, не полагаются на втягивание бедренной кости для движения вперёд, а вместо этого оттягивают голень вокруг коленного сустава[4][112].
Хвостовые позвонки (англ. caudals, от лат. cauda — хвост) — позвонки, составляющие хвост. У динозавров было около 50 хвостовых позвонков, хотя их количество, размер и форма значительно различались в разных группах. Число хвостовых костей уменьшалось по эволюционной линии, ведущей к современным птицам; у последних оставшиеся хвостовые кости сливаются в пигостиль. Несмотря на гибкость хвоста у ранних динозавров, некоторые группы укрепляли свой хвост с помощью окостеневших сухожилий (например, многие птицетазовые) или удлинённых презигапофизов (как у некоторых теропод, таких как дромеозавриды). У диплодоцидных завропод был удлинённый хлыстоподобный хвост, в то время как хвосты некоторых птицетазовых динозавров были снабжены утолщениями, шипами и / или пластинами[19]:45.
Хоаны[12]:166 (англ. choanae) — внутренние парные носовые отверстия в нёбе, которые продолжаются от наружных ноздрей, образующих носовой проход. Левое и правое отверстия разделены по средней линии черепа сошником. У динозавров хоаны обычно очень большие и удлинённые[19]:39[113].
Ф
Фаланга (лат. phalanx) — каждая из костей пальцев передней и задней конечности. Конечная фаланга пальца, называемая ногтевым суставом (англ. ungual), образовывает костную основу копыта или когтя[22]:145.
Ч
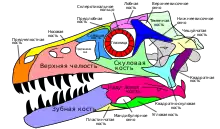
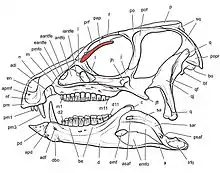
Череп (лат. cranium) — костная часть головы, защищающая её от повреждения и служащая местом прикрепления мягких тканей. Состоит из верхней части — собственно черепа, и нижней части — нижней челюсти. Верхняя часть состоит из черепной коробки, крыши черепа, верхней челюсти со скуловыми костями, и нёба. Черепная коробка имеет почти полностью эндохондральное происхождения, тогда как остальная часть черепа преимущественно состоит из костей дермы. Нижние челюсти состоят как из кожных, так и из эндохондральных костей. В черепе есть несколько отверстий, которые являются важными ориентирами в анатомических описаниях. К ним относятся: глазница (орбита), в которой находился глаз, а также наружные ноздри. Кроме того, в черепе были дополнительные отверстия, или окна, которые были типичны для диапсидных рептилий: предглазничное отверстие между ноздрёй и глазницей, подвисочное отверстие позади глазницы и надвисочное отверстие позади крыши черепа[22]:138—140[19]:32—36[14].
Черепная коробка (англ. braincase) — часть черепа, внутри которой находится мозг. В сочленённом черепе снаружи не видна, кроме своей задней части — затылочного бугра (лат. occiput). Является сложной структурой, пронизанной многочисленными отверстиями, содержащими кровеносные сосуды и нервы черепа. У взрослых особей отдельные кости черепной коробки, как правило, полностью сращены, причём границы между исходными элементами часто не видны. Анатомия черепной коробки динозавров консервативна, и по этой причине может использоваться, чтобы сделать вывод о взаимоотношениях группы, когда другие особенности скелета претерпели настолько глубокие изменения, что их происхождение более невозможно проследить. Черепная коробка может позволить реконструировать мозг и внутреннее ухо для оценки чувств и интеллекта[18].
Черепной кинетизм[21]:94 (англ. cranial kinesis) — способность частей черепа двигаться друг относительно друга в суставах. Хотя гипотеза черепного кинетизма была предложена для ряда таксонов нептичьих динозавров, обзор 2008 года обнаружил, что большинство из этих выводов проблематичны[114].
Чешуйчатая кость (лат. os squamosale, англ. squamosal bone) — парная кость в заднем углу нижней челюсти[19]:38.
Ш
Шевроны, или гемальные дуги — кости, прикреплённые к нижней стороне хвостовых позвонков, образующие вентральную поверхность хвоста. Шеврон состоит из левой и правой частей, которые обычно соединены друг с другом V-образным образом, охватывая большое отверстие, которое можно увидеть спереди или сзади. Отверстия нескольких последовательных шевронов образуют гемальный канал хвоста, который защищает нервы и кровеносные сосуды[24][19]:46.
Шейное полукольцо (англ. cervical half-ring) — поперечно ориентированный воротниковый ряд остеодерм, защищающих верхнюю часть шеи у анкилозавров (нижняя часть шеи остается открытой). У каждой особи обычно было 2 шейных полукольца, из которых переднее было меньше заднего. Шейные полукольца — это синапоморфия группы Ankylosauria[115].
Шейные позвонки (лат. vertebrae cervicales, англ. cervicals) — позвонки, составляющие шейный отдел позвоночника. У большинства динозавров было 9—10 шейных позвонков, хотя в некоторых группах, включая завропод, их было больше, как за счёт увеличения числа позвонков, так и за счёт интеграции грудных позвонков в шею[19]:44.
Шейные рёбра (англ. cervical ribs — кости, которые латеровентрально прикрепляются к шейным позвонкам. Они плезиоморфны для амниот (хотя и утеряны у млекопитающих) и включают передний и задний отростки. У некоторых завроподоморфов, особенно у некоторых завропод с длинной шеей, таких как жираффатитан, были чрезмерно удлинённые шейные ребра с задними отростками, перекрывающими 2 или 3 предшествующих позвонка. Гистологический анализ костей показал, что эти удлинённые отростки представляют собой окостеневшие сухожилия, что означает, что их концы были соединены с мышцами. Большая протяженность отростков увеличила бы расстояние между телом мышцы и позвонком, которым управляет мышца, что позволило бы расположить мышцу близко к телу, облегчая шею[116][117].
Э
Эктоптеригоид[12]:164, или наружная крыловидная кость[10]:193 (англ. ectopterygoid) — меньшая по размеру кость нёба. Она парная и кожная по происхождению и соединяется со скуловой костью латерально и с отростком крыловидной кости (англ. pterigoid) медиально[19]:40.

Эндокран (англ. endocast) — заполнение (слепок) нервных полостей, включая головной мозг и нервный канал позвонков. Таким образом, они могут регистрировать внешние особенности нервных структур, которые присутствовали в этих полостях, в первую очередь мозга. Однако, эндокраны не являются точными копиями нервных структур, поскольку нервные полости обычно содержат дополнительную ткань, которая может скрывать морфологию нервной структуры. Естественный эндокран образуется, когда нервная полость заполняется осадком, в то время как искусственные эндокраны могут быть изготовлены с использованием отливочного материала[60]:192.
Эпиптеригоид, или верхнекрыловидная кость, или столбчатая кость[10]:194 (англ. epipterygoid) — парная эндохондральная кость в задней части нёба. Представляет собой маленькую кость в форме пластинки, которая соединяется с крыловидным отростком (птеригоидом) и черепной коробкой[19]:39—40.
Эпипофизы (англ. epipophysis) — костные выступы шейных позвонков, встречающиеся у динозавров и некоторых ископаемых базальных птиц. Эти парные отростки располагаются над постзигапофизами в задней части дуг позвонков. Их морфология разнообразна и колеблется от небольших простых холмистых возвышений до больших сложных крыльевидных выступов. Эпипофизы обеспечивали большую площадь прикрепления нескольких шейных мышц, поэтому большие эпипофизы указывают на сильную мускулатуру шеи. Наличие эпипофизов считается синапоморфией динозавров[19]:14.
Эпистрофей (лат. axis) — второй шейный позвонок, следующий за атлантом. Он крупнее последнего, его тело сливается с дугой позвонка, а невральный отросток хорошо развит. Его небольшой интецентр intercentrum, расположенный между телом позвонка (centrum), отделён у теропод, но сливается с телом атланта у птицетазовых[11].
Я
Ямка (лат. fossa) — общий термин для обозначения углубления или бороздки на поверхности кости[24].
C
Caputegulae (лат.) — плоские кости, покрывающие черепа анкилозавров. Вместе с рогами пирамидальной формы они образуют орнамент черепа. Термин ввёл Уильям Т. Блоус в 2001 году[118] и может использоваться для обозначения элементов, представляющих как окостеневшие остеодермы, так и украшения черепа. Положение caputegulum на черепе можно уточнить с помощью специальных модификаторов: например, носовые caputegulae располагаются над носовыми костями[119].
Carinae (лат., мн. ч.; ед. ч. — carina) — эмалевые гребни, образующие режущие кромки на переднем и заднем краях зубов. Они обычно встречаются у хищных динозавров и часто имеют зазубрины[19]:41.
Carpometacarpus (лат.) — костный элемент кисти, состоящий из сросшихся костей запястья и пястных костей. Эта структура встречается у современных птиц, но также присутствовала у некоторых теропод, тесно связанных с птицами, и развивалась независимо у альваресзаврид[24].
Centrocoel (лат.) — пещеристая полость в теле позвонка, вмещающая костный мозг[20]:31, 47.
Cingulum (лат.; мн. ч. — cingula) — выпуклость в виде полочки, окружающая основание коронки зуба[40].
E
Epijugal (англ.) — оссификация кожи, уникальная для цератопсов. Она покрывает нижний конец обращённого вниз треугольного фланца скуловой кости, типичного для представителей этой группы. У Arrhinoceratops и Pentaceratops epijugal образует отчётливый скуловой рог. Эта кость может быть найдена у Yamaceratops и всех цератопсов более производных, чем последний[102][120].
Epoccipital (англ.) — кожные окостенения, выстилающие края воротников цератопсов. Являются отдельными костями у молодых особей, а у взрослых сливаются либо с чешуйчатыми, либо с теменными костями, в зависимости от своего расположения. Epoccipital чешуйчатых костей также известны как эпичешуйчатые кости (англ. episquamosals), а теменные — как эпипариетальные (англ. epiparietals). Некоторые авторы выступали против использования термина epoccipital, поскольку эти кости не имеют связи с затылочной областью черепа, вместо этого предпочитая термины «эпичешуйчатые» и «эпипариетальные». Epoccipital были скорее декоративными, чем функциональными элементами, и широко варьировались по форме, образуя сильно увеличенные шипы у центрозаврин[102][120]:540[103].
H
Hypex (англ.) — вершина угла, образованного между двумя пальцами. Hypex можно идентифицировать только при наличии мягких тканей, и поэтому он в основном используется при изучении окаменелых следов[5]:46—47.
M
Mesethmoid (англ.) — кость в передней части черепной коробки. Расположена под лобными костями и спереди от клиновидной кости и сочленяется спереди с межглазничной перегородкой. Состоит из горизонтальной дорсальной пластинки, расположенной непосредственно под лобными костями, от которой срединная перегородка (тонкая стенка кости на средней линии черепа) вентрально расширяется до места соединения с базисфеноидом. Как и в случае со клиновидно-решётчатой (спленэтмоидом) и глазнично-клиновидной костями (орбитосфеноидом), mesethmoid не окостеневает у многих видов и поэтому редко сохраняется; тем не менее, эта кость обычно встречается у птиц[19]:39[18][47].
P
Postparietal foramen (англ.) — непарное отверстие, расположенное на средней линии черепа между теменной и надзатылочной (англ. supraoccipital) костями, которое встречается у некоторых завроподоморфов. Как и в случае пинеального отверстия, оно, возможно, обнажало теменной глаз и, таким образом, служило для фоторецепции[79][121][122].
Posttemporal fenestra (англ.) — парное отверстие, расположенное ниже заднего края теменной части крыши черепа и над парокципитальным отростком черепной коробки. Возможно, представлял собой выход затылочной ветви глазной артерии[123].
S
Supratemporal fenestra (англ.) — парное отверстие, расположенное в задней части крыши черепа[22]:140. Обычно оно граничит с заглазничной, чешуйчатой и теменной костями[19]:38.
T
Thagomizer (англ.) — каждый из нескольких (от 4 до 10) хвостовых шипов стегозавров. Термин ввёл художник-карикатурист Гэри Ларсон в 1982 году[24].
Z
Ziphodont (англ.) — термин, относимый к зубам, имеющим форму лезвия, с загнутыми и приплюснутыми коронками, которые обычно несут пилообразную насечку. Зубы такой формы, адаптированные к хищному рациону их обладателей, изначально присутствуют у динозавров и встречаются у большинства нептичьих теропод[40].
Примечания
- Курочкин Е. Н., Лопатин А. В. Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Лопатин А. В.. — М.: ГЕОС, 2012. — Т. 2. — С. 314. — 419 (154 рис., 26 фототабл.) с. — ISBN 978-5-89118-594-4.
- Glossary // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 813–827. — ISBN 978-0-12-226810-6.
- Padian K. Pectoral girdle // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 530–536. — ISBN 978-0-12-226810-6.
- Dilkes D. W., Hutchinson J. R., Holliday C. M., Witmer L. M. Reconstructing the Musculature of Dinosaurs // The Complete Dinosaur (2 ed.) (англ.) / Brett-Surman M. K., Holtz T. R., Farlow J. O.. — 2012. — P. 150–190. — ISBN 978-0-253-00849-7.
- Leonardi G. Glossary and manual of tetrapod footprint palaeoichnology (англ.). — Brasilia: Publicação do Departemento Nacional da Produção Mineral Brasil, 1987. — P. 43—51.
- Snively E., Russell A. P., Powell G. L. Evolutionary morphology of the coelurosaurian arctometatarsus: descriptive, morphometric and phylogenetic approaches (англ.) // Zoological Journal of the Linnean Society. — 2004. — Vol. 142. — P. 525—553.
- Snively E. Functional morphology of the tyrannosaurid arctometatarsus. — Calgary, Alberta: The University of Calgary, 2000. — 273 p.
- Хоун Д. Хроники тираннозавра: Биология и эволюция самого известного хищника в мире / науч. ред. А. Аверьянов. — М.: Альпина нон-фикшн, 2017. — С. 182. — 358 с. — ISBN 978-5-91671-744-0.
- Tsuihiji T., Makovicky P. J. Homology of the neoceratopsian cervical bar elements (англ.) // Journal of Paleontology. — 2007. — Vol. 81, no. 5. — P. 1132–1138. — doi:10.1666/pleo05-164.1.
- Основы палеонтологии : Справочник для палеонтологов и геологов СССР : в 15 т. / гл. ред. Ю.А. Орлов. — М. : Наука, 1964. — Т. 12 : Земноводные, пресмыкающиеся и птицы / под ред. А.К. Рождественского, Л.П. Татаринова. — 724 с. — 3000 экз.
- Romer A. S. The Axial Skeleton // Osteology of the Reptiles (англ.). — The University of Chicago Press, 1976. — P. 218–297. — ISBN 978-0-226-72491-1.
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009. — Т. 291. — 377 с. — (Труды ПИН РАН). — ISBN 978-5-89118-461-9.
- Prieto-Márquez A., Norell M. A. Redescription of a nearly complete skull of Plateosaurus (Dinosauria: Sauropodomorpha) from the Late Triassic of Trossingen (Germany) (англ.) // American Museum Novitates. — 2011. — Vol. 3727. — P. 1–58. — doi:10.1206/3727.2.
- Romer A. S. The Skull — General Structure // Osteology of the Reptiles (англ.). — The University of Chicago Press, 1976. — P. 55–83. — ISBN 978-0-226-72491-1.
- Romer A. S. The Skull in the Various Reptile Groups // Osteology of the Reptiles (англ.). — The University of Chicago Press, 1976. — P. 85–197. — ISBN 978-0-226-72491-1.
- Алифанов В. Р. Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Курочкин Е. Н., Лопатин А. В.. — М.: Геос, 2012. — Т. 2. — 419 (154 рис., 26 фототабл.) с. — ISBN 978-5-89118-594-4.
- Rogers K. C., Forster C. A. The skull of Rapetosaurus krausei (sauropoda: Titanosauria) from the Late Cretaceous of Madagascar (англ.) // Journal of Vertebrate Paleontology. — 2004. — Vol. 24, no. 1. — P. 121–144. — doi:10.1671/A1109-10.
- Currie P. J. Braincase Anatomy // Encyclopedia of Dinosaurs / Currie P. J., Padian K.. — 1997. — С. 81—85. — ISBN 978-0-12-226810-6.
- Brusatte S. L. Dinosaur Paleobiology (англ.). — John Wiley & Sons, 2012. — ISBN 978-0-470-65657.
- Madsen J. H., Welles S. P. Ceratosaurus (Dinosauria, Theropoda): A Revised Osteology (англ.). — Utah Geological Survey, 2000. — P. 1—80. — ISBN 978-1-55791-380-7.
- Нэйш Д., Барретт П. Динозавры: 150 000 000 лет господства на Земле / Аверьянов А., д-р биол. наук.. — М.: Альпина нон-фикшн, 2019. — 223 с. — ISBN 978-5-91761-940-6.
- Holtz T. R., Brett-Surman M. K. The Osteology of the Dinosaurs // The Complete Dinosaur (2 ed.) (англ.) / Brett-Surman M. K., Holtz T. R., Farlow J. O.. — 2012. — P. 135–149. — ISBN 978-0-253-00849-7.
- Wu X.-C., Russell A. P. Functional morphology // Encyclopedia of dinosaurs (англ.) / Padian K., Currie P. J.. — San Diego: Academic Press, 1997. — P. 258–268. — ISBN 978-0-12-226810-6.
- Glossary // The Complete Dinosaur (2 ed.) / Brett-Surman M. K. Holtz T. R., Farlow J. O.. — 2012. — С. 1075–1081. — ISBN 978-0-253-00849-7.
- Vickaryous V., Maryanska T., Weishampel D. B. Chapter Seventeen: Ankylosauria // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — University of California Press, 2004. — ISBN 978-0520254084.
- Wedel M. J. Evidence for bird-like air sacs in saurischian dinosaurs (англ.) // Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. — 2009. — Vol. 311, no. 8. — P. 611–628. — doi:10.1002/jez.513.
- Fastovsky D. E., Weishampel D. B. Dinosaurs: A Concise Natural History (англ.). — Cambridge: GBR: Cambridge University Press, 2009. — 358 p. — ISBN 978-0-511-47789-8.
- Vickaryous M. K., Maryańska T. M., Weishampel D. B. Ankylosauria // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2009. — P. 363–392. — ISBN 978-0-520-24209-8.
- Sales M. A. F., Schultz C. L. Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil (англ.) // PLOS One. — 2017. — Vol. 12, no. 11. — P. e0187070. — doi:10.1371/journal.pone.0187070.
- Holtz T. R., Molnar R. E., Currie P. J. Basal Tetanurae // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2004. — P. 71—110. — ISBN 978-0-520-24209-8.
- Galton P. M. Stegosaurs // The Complete Dinosaur (2 ed.) (англ.) / Brett-Surman M. K., Holtz T. R., Farlow J. O.. — 2012. — P. 482–504. — ISBN 978-0-253-00849-7.
- Galton P. M., Upchurch P. Prosauropoda // The Complete Dinosaur (2 ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2004. — P. 232–258. — ISBN 978-0-520-24209-8.
- Claessens L. P. A. M. Dinosaur gastralia; origin, morphology, and function (англ.) // Journal of Vertebrate Paleontology. — 2004. — Vol. 24, no. 1. — P. 89—106. — doi:10.1671/A1116-8.
- Hyman L. H. Comparative Vertebrate Anatomy (2nd ed.) (англ.). — Chicago: The University of Chicago Press, 1942. — P. 177.
- Coombs W. P. The bony eyelid of Euoplocephalus (Reptilia, Ornithischia) (англ.) // Journal of Paleontology. — 1972. — Vol. 46, no. 5. — P. 637–650. — ISSN 0022-3360.
- Синельников Р. Д., Синельников Я. Р., Синельников А. Я. Атлас анатомии человека: Учебное пособие в 4 томах. — М.: РИА «Новая волна»: Издатель Умеренков, 2009. — Т. 1. — 344 с. — ISBN 978-5-7864-0199-9.
- Galton P. M. Notes on the postcranial anatomy of the heterodontosaurid dinosaur Heterodontosaurus tucki, a basal ornithischian from the Lower Jurassic of South Africa (англ.) // Revue de Paléobiologie, Genève. — 2014. — Vol. 1, no. 33. — P. 97–141. — ISSN 1661—5468.
- Läng É., Goussard F. Redescription of the wrist and manus of ?Bothriospondylus madagascariensis: new data on carpus morphology in Sauropoda (англ.) // Geodiversitas. — 2007. — Vol. 29, no. 4. — P. 549–560.
- Martin A. J. Introduction to the Study of Dinosaurs (2nd ed.) (англ.). — Blackwell Publishing, 2006. — 577 p. — ISBN 978-1-4051-3413-.
- Sander P. M. Teeth and jaws // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 717—725. — ISBN 978-0-12-226810-6.
- LeBlanc A. R. H., Reisz R. R., Evans D. C., Bailleul A. M. Ontogeny reveals function and evolution of the hadrosaurid dinosaur dental battery (англ.) // BMC Evolutionary Biology. — 2016. — Vol. 16, no. 1. — P. 152. — doi:10.1186/s12862-016-0721-1.
- Erickson G. M. Tooth Replacement Patterns // Encyclopedia of dinosaurs / Padian K., Currie P. J.. — San Diego: Academic Press, 1997. — С. 739–740. — ISBN 978-0-12-226810-6.
- Currie P. J., Padian K. Thecodontia // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 728. — ISBN 978-0-12-226810-6.
- DeBraga M., Carroll R. L. The Origin of Mosasaurs As a Model of Macroevolutionary Patterns and Processes // Evolutionary Biology (англ.) / Hecht M. K., MacIntyre R. J., Clegg M. T.. — Boston, MA: Springer, 1993. — Vol. 27. — P. 245—322. — ISBN 978-1-4613-6248-7.
- Hendrickx C., Mateus O., Buffetaut E. Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa (англ.) // PLoS ONE. — 2016. — Vol. 11, no. 1. — P. e0144695. — doi:10.1371/journal.pone.0144695.
- Padian K. Saurischia // Encyclopedia of dinosaurs (англ.) / Padian K., Currie P. J.. — San Diego: Academic Press, 1997. — P. 647–653. — ISBN 978-0-12-226810-6.
- Ali F., Zelenitsky D. K., Therrien F., Weishampel D. B. Homology of the "ethmoid complex" of tyrannosaurids and its implications for the reconstruction of the olfactory apparatus of non-avian theropods (англ.) // Journal of Vertebrate Paleontology. — 2008. — Vol. 28, no. 1. — P. 123–133. — doi:10.1671/0272-4634(2008)28[123:HOTECO]2.0.CO;2.
- Tschopp E., Mateus O. Clavicles, interclavicles, gastralia, and sternal ribs in sauropod dinosaurs: new reports from Diplodocidae and their morphological, functional and evolutionary implications (англ.) // Journal of Anatomy. — 2013. — Vol. 222, no. 3. — P. 321–340. — doi:10.1111/joa.12012.
- Schünke M., Schulte E., Schumacher U. Thieme Atlas of Anatomy: General Anatomy and Musculoskeletal System (англ.) / Ross L. M., Lamperti E. D.. — Thieme. — ISBN 978-3-13-142071-8.
- Wagner G. P., Chiu, Chi-Hua. The tetrapod limb: a hypothesis on its origin (англ.) // Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. — 2001. — Vol. 291, no. 3. — P. 226–240. — doi:10.1002/jez.1100.
- Padian K. Skeletal structures // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 668. — ISBN 978-0-12-226810-6.
- Bocherens H. Chemical composition of dinosaur fossils // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — San Diego: Academic Press, 1997. — P. 111—117. — ISBN 978-0-12-226810-6.
- Rasskin-Gutman D. Pelvis, comparative anatomy // Encyclopedia of dinosaurs (англ.) / Padian K., Currie P. J.. — San Diego: Academic Press, 1997. — P. 536–540. — ISBN 978-0-12-226810-6.
- Hutchinson J. R. The evolution of pelvic osteology and soft tissues on the line to extant birds (Neornithes) (англ.) // Zoological Journal of the Linnean Society. — 2001. — Vol. 131, no. 2. — P. 123–168. — doi:10.1111/j.1096-3642.2001.tb01313.x.
- Nesbitt S. J. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades (англ.) // Bulletin of the American Museum of Natural History. — 2011. — Vol. 352. — P. 1–292. — ISSN 0003-0090. — doi:10.1206/352.1.
- Reisdorf A. G., Wuttke M. Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in fossil vertebrates. Part I: Reptiles - The taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany) (англ.) // Palaeobiodiversity and Palaeoenvironments. — 2012. — Vol. 92, no. 1. — P. 119–168. — doi:10.1007/s12549-011-0068-y.
- Frankfurt N. G., Chiappe L. M. A possible oviraptorosaur from the Late Cretaceous of northwestern Argentina (англ.) // Journal of Vertebrate Paleontology. — 1999. — Vol. 19, no. 1. — P. 101—105. — doi:10.1080/02724634.1999.10011126.
- Лопатин А., Аверьянов А. Древнейшие плацентарные: начало истории успеха // Природа. — 2018. — № 4. — С. 34—40.
- Romer A. S. The Mandible // Osteology of the Reptiles (англ.). — The University of Chicago Press, 1976. — P. 198–217. — ISBN 978-0-226-72491-1.
- Holtz T. R., Brett-Surman M. K. Dinosaur Paleoneurology // The Complete Dinosaur (2 ed.) (англ.) / Brett-Surman M. K. Holtz T. R., Farlow J. O.. — 2012. — P. 191–208. — ISBN 978-0-253-00849-7.
- Kundrát M. Avian-like attributes of a virtual brain model of the oviraptorid theropod Conchoraptor gracilis (англ.) // Naturwissenschaften. — 2007. — Vol. 94, no. 6. — P. 499–504. — doi:10.1007/s00114-007-0219-1.
- Черепанов Г. О., Иванов А. О. Ископаемые высшие позвоночные. — 2-е изд. — СПб.: Изд-во С.-Петербургского ун-та, 2007. — С. 200. — 202 с. — ISBN 978-5-288-04308-6.
- Padian K. Ornithischia // Encyclopedia of dinosaurs (англ.) / Padian K., Curry P. J.. — San Diego: Academic Press, 1997. — P. 494–498. — ISBN 978-0-12-226810-6.
- Ford T. L. A review of ankylosaur osteoderms from New Mexico and a preliminary review of ankylosaur armor (англ.) // New Mexico Museum of Natural History and Science Bulletin. — 2000. — Vol. 17. — P. 157–176.
- Arbour V. M., Burns M. E., Bell P. R., Currie P. J. Epidermal and dermal integumentary structures of ankylosaurian dinosaurs (англ.) // Journal of Morphology. — 2014. — Vol. 275, no. 1. — P. 39–50. — doi:10.1002/jmor.20194.
- D'Emic M. D., Wilson J. A., Chatterjee S. The titanosaur (Dinosauria: Sauropoda) osteoderm record: review and first definitive specimen from India (англ.) // Journal of Vertebrate Paleontolog. — 2009. — Vol. 29, no. 1. — P. 165–177. — doi:10.1671/039.029.0131.
- Novas F. E. The age of dinosaurs in South America (англ.). — Bloomington: Indiana University Press, 2009. — ISBN 978-0-253-35289-7.
- Bailey J. B. Neural spine elongation in dinosaurs: sailbacks or buffalo-backs? (англ.) // Journal of Paleontology. — 1997. — Vol. 71, no. 6. — P. 1124—1146. — doi:10.1017/S0022336000036076.
- Upchurch P., Barrett P. M., Dodson P. Sauropoda // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmolska H.. — Berkeley: University of California Press, 2004. — P. 259–322. — ISBN 978-0-520-24209-8.
- Hone D. W. E., Holtz T. R. A Century of Spinosaurs — A Review and Revision of the Spinosauridae with Comments on Their Ecology (англ.) // Acta Geologica Sinica — English Edition. — 2017. — Vol. 91, no. 3. — P. 1120—1132. — doi:10.1111/1755-6724.13328.
- Ortega F., Escaso F., Sanz J. L. A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain (англ.) // Nature. — 2010. — Vol. 467, no. 7312. — P. 203–206. — doi:10.1038/nature09181.
- Allain R., Xaisanavong T., Richir P., Khentavong B. The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos (англ.) // Naturwissenschaften. — 2012. — Vol. 99. — P. 369—377. — ISSN 0028-1042. — doi:10.1007/s00114-012-0911-7.
- Eddy D. R., Clarke J. A. New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda) (англ.) // PLOS One. — 2011. — Vol. 6, no. 3. — P. e17932. — doi:10.1371/journal.pone.0017932.
- Benton M. J. Prehistoric Life (англ.). — Edinburgh, Scotland: Dorling Kindersley, 2012. — P. 338. — ISBN 978-0-7566-9910-9.
- Prieto-Márquez A., Chiappe L. M., Joshi S. H. The Lambeosaurine Dinosaur Magnapaulia laticaudus from the Late Cretaceous of Baja California, Northwestern Mexico (англ.) // PLOS One. — 2012. — Vol. 7, no. 6. — P. e38207. — doi:10.1371/journal.pone.0038207.
- Paul G. S. The Princeton Field Guide to Dinosaurs (англ.). — Princeton University Press, 2010. — P. 208–209.
- Baumel J. J. Handbook of Avian Anatomy: Nomina Anatomica Avium (англ.). — Cambridge, 1993. — P. 57, 92. — 779 p. — (Nuttall Ornithological Club, no. 23).
- Barsbold R., Osmólska H., Watabe M., Currie P. J., Tsogtbaatar K. New Oviraptorosaur (Dinosauria, Theropoda) From Mongolia: The First Dinosaur With A Pygostyle (англ.) // Acta Palaeontologica Polonica. — 2000. — Vol. 45, no. 2. — P. 97—106.
- Paulina Carabajal A., Carballido J. L., Currie P. J. Braincase, neuroanatomy, and neck posture of Amargasaurus cazaui (Sauropoda, Dicraeosauridae) and its implications for understanding head posture in sauropods (англ.) // Journal of Vertebrate Paleontology. — 2014. — Vol. 34, no. 4. — P. 870–882. — doi:10.1080/02724634.2014.838174.
- Edinger T. The size of parietal foramen and organ in reptiles: a rectification (англ.) // Bulletin of the Museum of Comparative Zoology. — 1955. — Vol. 114. — P. 1–34.
- Lehman T. M. A horned dinosaur from the El Picacho Formation of West Texas, and review of ceratopsian dinosaurs from the American Southwest (англ.) // Journal of Paleontology. — 1996. — Vol. 70, no. 3. — P. 494–508. — doi:10.1017/S0022336000038427.
- Wilson J. A. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs (англ.) // Journal of Vertebrate Paleontology. — 1999. — Vol. 19, no. 4. — P. 639–653. — doi:10.1080/02724634.1999.10011178.
- Langer M. C., Benton M. J. Early dinosaurs: a phylogenetic study (англ.) // Journal of Systematic Palaeontology. — 2006. — Vol. 4, no. 4. — P. 309–358. — doi:10.1017/S1477201906001970.
- Gauthier J. Saurischian monophyly and the origin of birds (англ.) // Memoirs of the California Academy of Sciences. — 1986. — Vol. 8. — P. 1—55.
- Novas F. E. Dinosaur monophyly (англ.) // Journal of Vertebrate Paleontology. — 1996. — Vol. 16, no. 4. — P. 723—741. — doi:10.1080/02724634.1996.10011361.
- Wells S. P. Dilophosaurus wetherilli (Dinosauria, Theropoda), osteology and comparisons (англ.) // Palaeontographica Abteilung A. — 1984. — Vol. 185. — P. 85–180.
- Charig A. J., Milner A. C. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey" (англ.) // Bulletin of the Natural History Museum of London. — 1997. — Vol. 53. — P. 11–70.
- Paul G. S. Predatory Dinosaurs of the World (англ.). — New York: Simon & Schuster, 1988. — P. 271–274.
- Thulborn R. A. Dinosaur tracks (англ.). — London, New York: Chapman and Hall, 1990. — P. 112–116. — ISBN 978-0-412-32890-9.
- Currie P. J., Sarjeant W. A. S. Lower cretaceous dinosaur footprints from the peace River Canyon, British Columbia, Canada (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology. — 1979. — Vol. 28. — P. 103–115. — doi:10.1016/0031-0182(79)90114-7.
- Faux C. M., Padian K. The opisthotonic posture of vertebrate skeletons: postmortem contraction or death throes? (англ.) // Paleobiology. — 2007. — Vol. 33, no. 2. — P. 201—226. — doi:10.1666/06015.1.
- Holtz T. R. Tyrannosauroidea // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2004. — P. 111–136. — ISBN 978-0-520-24209-8.
- Osmólska H., Currie P. J., Barsbold R. Oviraptorosauria // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2004. — P. 165–183. — ISBN 978-0-520-24209-8.
- Preushscoft H., Witzel U. Biomechanical Investigations on the Skulls of Reptiles and Mammals (англ.) // Senckenbergiana Lethaea. — 2002. — Vol. 82. — P. 207—222.
- Rayfield E. J., Milner A. C., Xuan V. B., Young P. G. Functional Morphology of Spinosaur "Crocodile Mimic" Dinosaurs (англ.) // JVP. — Vol. 27, no. 4. — P. 892—901.
- Witmer, L.M. The Evolution of the Antorbital Cavity of Archosaurs: A Study in Soft-Tissue Reconstruction in the Fossil Record with an Analysis of the Function of Pneumaticity (англ.) // JVP. — 1997. — Vol. 17, no. 1. — P. 1—76.
- Padian K. Phylogeny of dinosaurs // Encyclopedia of dinosaurs (англ.) / Padian K., Currie P. J.. — San Diego: Academic Press, 1997. — P. 646–651. — ISBN 978-0-12-226810-6.
- Padian K. Bipedality // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 68—70. — ISBN 978-0-12-226810-6.
- Schachner E. R., Lyson T. R., Dodson P. Evolution of the respiratory system in nonavian theropods: evidence from rib and vertebral morphology (англ.) // The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology: Advances in Integrative Anatomy and Evolutionary Biology. — 2009. — Vol. 292, no. 9. — P. 1501–1513. — doi:10.1002/ar.20989.
- Hieronymus T., Witmer L. Cranial rugosity and dinosaur "horns:" Rhino and giraffe as model systems for skin reconstruction in fossil taxa (англ.) // Journal of Vertebrate Paleontology. — 2004. — Vol. 24. — P. 70–70A.
- Witmer L. M. The evolution of the antorbital cavity of archosaurs: a study in soft-tissue reconstruction in the fossil record with an analysis of the function of pneumaticity (англ.) // Journal of Vertebrate Paleontology. — 1997. — Vol. 17, no. S1. — P. 1–76. — doi:10.1080/02724634.1997.10011027.
- Dodson P., Forster C. A., Sampson S. D. Ceratopsidae // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — University of California Press, 2004. — P. 494—513. — ISBN 978-0520254084.
- Horner J. R., Goodwin M. B. Ontogeny of cranial epi-ossifications in Triceratops (англ.) // Journal of Vertebrate Paleontology. — 2008. — Vol. 28, no. 1. — P. 134–144. — doi:10.1671/0272-4634(2008)28[134:OOCEIT]2.0.CO;2.
- Schmitz L. Quantitative estimates of visual performance features in fossil birds (англ.) // Journal of Morphology. — 2009. — Vol. 270, no. 6. — P. 759–773. — doi:10.1002/jmor.10720.
- Yamashita M., Konishi T., Sato T. Sclerotic Rings in Mosasaurs (Squamata: Mosasauridae): Structures and Taxonomic Diversity (англ.) // PLOS One. — 2015. — Vol. 10, no. 2. — P. e0117079. — doi:10.1371/journal.pone.0117079.
- Fiorillo A. R. Taphonomy // Encyclopedia of Dinosaurs (англ.) / Currie P. J., Padian K.. — 1997. — P. 713—716. — ISBN 978-0-12-226810-6.
- Naish D. Birds // The complete dinosaur (англ.) / Brett-Surman M. K., Holtz T. R., Farlow J. O.. — Indiana University Press, 2012. — P. 379–423.
- Fronimos J. A., Wilson J. A., Baumiller T. K. Polarity of concavo-convex intervertebral joints in the necks and tails of sauropod dinosaurs (англ.) // Paleobiology. — 2016. — Vol. 42, no. 4. — P. 624–642. — doi:10.1017/pab.2016.16.
- Kardong K. V. Skeletal System: The Axial Skeleton // Vertebrates — Comparative Anatomy, Function, Evolution (6. ed.) (англ.). — Mc Graw Hill, 2011. — P. 294–324. — 816 p. — ISBN 978-9-33-922211-6.
- Dodson P. Comparative craniology of the Ceratopsia (англ.) // American Journal of Science. — 1993. — Vol. 293A. — P. 200–234. — doi:10.2475/ajs.293.A.200.
- Brown C. M., Russell A. P. Homology and architecture of the caudal basket of Pachycephalosauria (Dinosauria: Ornithischia): the first occurrence of myorhabdoi in Tetrapoda (англ.) // PLOS ONE. — 2012. — Vol. 7, no. 1. — P. e30212. — doi:10.1371/journal.pone.0030212.
- Persons IV W. S., Currie P. J. The tail of Tyrannosaurus: reassessing the size and locomotive importance of the M. caudofemoralis in non-avian theropods (англ.) // The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. — 2011. — Vol. 294, no. 1. — P. 119–131. — doi:10.1002/ar.21290.
- Weishampel D. B. The nasal cavity of lambeosaurine hadrosaurids (Reptilia: Ornithischia): comparative anatomy and homologies (англ.) // Journal of Paleontology. — 1981. — P. 1046–1057.
- Holliday C. M., Witmer L. M. Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids (англ.) // Journal of Vertebrate Paleontology. — 2008. — Vol. 28, no. 4. — P. 1073–1088. — doi:10.1671/0272-4634-28.4.1073.
- Blows W. T. Notice of nodosaur (Dinosauria, Ankylosauria) remains from the mid-Cretaceous of Cambridge, England, with comments on cervical half-ring armour (англ.) // Proceedings of the Geologists' Association. — 2014. — Vol. 125, no. 1. — P. 56–62. — doi:10.1016/j.pgeola.2013.07.001.
- Christian A., Dzemski G. Reconstruction of the cervical skeleton posture of Brachiosaurus brancai Janensch, 1914 by an analysis of the intervertebral stress along the neck and a comparison with the results of different approaches (англ.) // Fossil Record. — 2007. — Vol. 10, no. 1. — P. 38–49. — doi:10.1002/mmng.200600017.
- Klein N., Christian A., Sander P. M. Histology shows that elongated neck ribs in sauropod dinosaurs are ossified tendons (англ.) // Biology Letters. — 2012. — Vol. 8, no. 6. — P. 1032–1035. — doi:10.1098/rsbl.2012.0778.
- Blows W. T. Dermal Armor of the Polacanthine Dinosaurs // The Armored Dinosaurs / Carpenter K.. — Indiana University Press, 2001. — С. 363–385. — ISBN 978-0-253-33964-5.
- Arbour V. M., Currie P. J. Euoplocephalus tutus and the Diversity of Ankylosaurid Dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA (англ.) // PLOS One. — 2013. — Vol. 8, no. 5. — P. e62421. — doi:10.1371/journal.pone.0062421.
- Makovicky P. Marginocephalia // The Complete Dinosaur (2 ed.) (англ.) / Brett-Surman M. K., Holtz T. R., Farlow J. O.. — 2012. — P. 526–549. — ISBN 978-0-253-00849-7.
- Harris J. D. The significance of Suuwassea emilieae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution (англ.) // Journal of Systematic Palaeontology. — 2006. — Vol. 4, no. 2. — P. 185–198. — doi:10.1017/S1477201906001805.
- Apaldetti C., Martinez R. N., Pol D., Souter T. Redescription of the skull of Coloradisaurus brevis (Dinosauria, Sauropodomorpha) from the Late Triassic Los Colorados Formation of the Ischigualasto-Villa Union Basin, northwestern Argentina (англ.) // Journal of Vertebrate Paleontology. — 2014. — Vol. 34, no. 5. — P. 1113–1132. — doi:10.1080/02724634.2014.859147.
- Langer M. C. Basal Saurischia // The Dinosauria (2nd ed.) (англ.) / Weishampel D. B., Dodson P., Osmólska H.. — Berkeley: University of California Press, 2004. — P. 25–46. — ISBN 978-0-520-24209-8.