Бипедализм
Бипедали́зм (также бипедальность или двуногость) — способ локомоции (передвижения) по твёрдой поверхности при помощи двух парных конечностей (ног). Характерен, в первую очередь, для многих птиц и нептичьих динозавров, некоторых сумчатых млекопитающих, тушканчиков, тушканчикоподобных грызунов (например, мешотчатых прыгунов) и человека. Также был характерен для некоторых примитивных архозавриформ. Животные, передвигающиеся на двух конечностях, называются бипедами.
В природе
В природе бипедализм неоднократно возникал в ходе эволюции независимо у различных групп наземных хордовых животных. Различают факультативный (используемый животным лишь время от времени, как, например, у утконосых динозавров) и облигатный (постоянный, как у страуса или человека) бипедализм.
Примитивные группы древних амниот изначально передвигались на четырёх широко расставленных в стороны конечностях, которые при локомоции раздвигались медиалатерально и не были способны постоянно поддерживать туловище над землёй (отсюда — «пресмыкание»), что соответствовало характерному для них менее интенсивному обмену веществ и сравнительно малоподвижному образу жизни. Бипедальными стали представители завропсид группы архозавриформ и синапсид группы цинодонтов. Соответствующие изменения в анатомии сопровождались значительным повышением интенсивности обмена веществ, поскольку постоянное поддержание тела над землёй и длительная локомоторная активность требовали намного больших энергозатрат по сравнению с «пресмыканием». Однако проследить подобные изменения на палеонтологическом материале сложно. Достоверно известно, однако, что современные представители данных групп амниот — соответственно млекопитающие и часть архозавров (птицы) — имеют интенсивный обмен веществ и эндотермную систему терморегуляции (теплокровность).
Завропсиды
У представителей клады завропсид, или рептилий, группы, ныне представленной пресмыкающимися[lower-alpha 1] и птицами, бипедализм впервые возник в среднем триасе у древних архозавров группы орнитодир и получил дальнейшее развитие у одних из её представителей, динозавроморф.

Современные пресмыкающиеся в основном сохраняют подобный (примитивный в эволюционном плане) характер локомоции, однако у некоторых продвинутых групп древних рептилий её развитие пошло в ином направлении — их конечности стали перемещаться непосредственно под туловище, занимая более габитуально вертикальное (выпрямленное) положение, что соответствовало более подвижному образу жизни, связанному с бегом и активным преследованием добычи (из современных нептичьих рептилий только у крокодилов конечности могут занимать положение под туловищем, причём исключительно при беге).
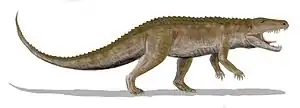
У ранних архозавриформ лобковая и седалищная кости начали удлиняться, передние ноги — укорачиваться, а тяжёлый длинный хвост, практически лишившийся подвижности из-за срастания тазовых костей, превратился в балансир, уравновешивающий массу вытянутого туловища. Задние конечности при ходьбе стали перемещаться в парасагиттальной плоскости, голеностопный сустав получил возможность пружинить при ходьбе, что способствовало более экономному расходу сил, а опора переместилась на средний (третий) палец. Это позволило многим группам архозавриформ перейти к факультативной, или даже габитуальной, бипедальности, подобно современным бегающим птицам, специализировавшись на беге на двух ногах.
Более продвинутые архозавриформы — ранние динозавры, как ящеротазовые, так и птицетазовые, — были активными хищниками, как минимум часть времени передвигавшимися на задних конечностях, что способствовало их быстрому прогрессу, особенно после вымирания большинства цинодонтов на рубеже пермского и триасового периода. В юрском периоде почти все более-менее крупные наземные животные (свыше метра в длину) были динозаврами.
Постепенно часть динозавров переходит на растительную пищу, занимая освободившиеся экологические ниши. Большинство растительноядных видов вновь возвращается к передвижению на четырёх конечностях, эволюция же хищников продолжает идти по «двуногому» пути (у абелизаврид передние конечности оказались практически редуцированны, а у манирапторов — напротив, развились в специализированную хватательную форму). В течение 140 млн лет, вплоть до конца мелового периода, основными наземными хищными позвоночными были бипедальные динозавры.
Бипедальные динозавры были предками птиц и, исходя из принципов кладистической классификации, птицы сами являются современными бипедальными динозаврами. Факультативный бипедализм предполагается и для некоторых сравнительно поздних видов крокодилов.
Среди современных пресмыкающихся (нептичьих завропсид) бипедализм редок. В качестве примера пресмыкающегося с (факультативно) бипедальной локомоцией можно привести плащеносную ящерицу. В отличие от древних двуногих рептилий, у неё сохраняется «растопыренное» расположение конечностей, так что бег на задних ногах в её исполнении выглядит несколько неуклюже.
Птицы
Птицы произошли от более примитивных хищных ящеротазовых динозавров. Бипедализм — изначальная черта птиц, покрытые перьями передние конечности которых превратились в крылья, а вся остальная анатомия адаптировалась к полёту.
Впоследствии часть птиц вновь вернулась к полностью наземному образу жизни, утратив способность летать, и утратила киль и их крылья утратили летательную функцию. Эти группа птиц известна как бескилевые птицы. У многих видов нелетающих птиц развилась способность к быстрому бегу. Так взрослый африканский страус способен развивать скорость до 70 км/ч.
Ещё одна группа птиц — пингвины, также утратила способность к полёту. Крылья этих птиц, ставших полуводными животными, трансформировались в ласты. Передвигаются по твёрдой поверхности они также на двух задних конечностях. При этом позвоночник пингвинов располагается практически вертикально.
Синапсиды
У ранних цинодонтов хвост значительно уменьшается в размерах и перестаёт играть какую-либо роль в локомоции (впоследствии у некоторых форм мог вторично появляться длинный хвост, однако он уже скорее напоминал крысиный и не имел мощной мускулатуры, характерной для хвоста других рептилий), позвоночник приобретает способность к вертикальному изгибу во время бега на четырёх ногах (при одновременном уменьшении способности к поперечному изгибу), лобковая и седалищная кости уменьшаются в размерах, а подвздошная — увеличивается, обеспечивая надёжную опору для сильных мышц конечностей. Эта группа синапсид стала специализироваться к бегу на четырёх ногах, аналогично современным хищным млекопитающим, и достигла большого эволюционного успеха на протяжении пермского периода, но по какой-то причине практически сошла со сцены в мезозое — большинство мезозойских форм цинодонтов, в том числе млекопитающих, имеют очень мелкие размеры.
Млекопитающие
Наибольшее распространение бипедализм получил среди сумчатых млекопитающих (кенгуру, кенгуровые крысы и др.). Судя по некоторым сообщениям очевидцев, недавно вымерший тилацин мог вставать на задние лапы во время преследования добычи, переходя с галопа на прыжки, подобно кенгуру.
Среди плацентарных млекопитающих бипедализм редок и является скорее исключением. Видимо, причиной этого является отсутствие у млекопитающих тяжёлого мясистого хвоста, способного служить при ходьбе на двух ногах в качестве балансира (утраченного ещё их давними предками, зверозубыми рептилиями, в ходе адаптации к бегу на четырёх ногах). Из-за этого большинство бипедальных млекопитающих перемещаются не бегом, а прыжками на задних лапах. Переход предков человека к облигатной бипедальности и быстрому бегу на двух ногах потребовал значительных изменений их анатомии.
В качестве примеров можно привести лептиктидий, тушканчиковых, долгоногов и человека. Многие обезьяны в норме передвигаются на четырёх ногах, однако могут ходить и на двух, в неволе иногда даже переходя к облигатному бипедализму. Например, бонобо используют бипедальную локомоцию при переноске детёнышей или предметов, изредка для добычи пищи. Вставать на задние лапы могут, к примеру, медведи, однако в природе пользуются этим не для передвижения, а для обознения окрестностей.
Ходить на задних лапах могут дрессированные кошки и собаки.
Переход предков человека к хождению на двух задних конечностях способствовал развитию передних конечностей (рук), с помощью которых было удобно пользоваться примитивными орудиями труда, переносить пищу из одного места в другое. Развитие ручного труда, в свою очередь, способствовало развитию головного мозга. Вместе с тем переход гоминид к бипедальности имел и негативный побочный эффект — человек стал фактически единственным млекопитающим, детёныши которого рождаются с не до конца сформированными черепной коробкой и головным мозгом. На самом деле именно «недоразвитый» в момент рождения мозг ребёнка позволяет ему освоить такой объём информации, который позволяет адаптироваться к жизни в сложном человеческом обществе, включая использование членораздельной речи, абстрактного мышления и сложных орудий труда (техники). Если бы человек рождался с более полно сформированным мозгом, как у детёныша обезьяны, он «пропустил» бы в утробе матери много времени, в норме уходящего на обучение, и со временем серьёзно отстал бы от «нормальных» детей в умственном развитии, в частности — скорее всего, не смог бы научиться говорить, как «дети-маугли».
В технике
Ввиду технических сложностей исполнения, механизмы, использующие бипедальный способ локомоции, пока не получили большого распространения. Из используемых на практике механизмов можно назвать шагающие экскаваторы. Также существуют проекты роботов, передвигающихся на двух ногах. Примерами роботов-шагоходов такого типа могут служить ASIMO[1], TOPIO[2], Atlas и другие.
См. также
Ссылки
- Иорданский Н. Н. ГЛАВА 3. МЕЗОЗОЙСКАЯ ЭРА - ВЕК РЕПТИЛИЙ // Эволюция жизни. — М.: Академия, 2001. — 425 с.
- Гилёв А. Н. Латерализация моторных функций конечностей у сумчатых ( Marsupialia ). — С.-Пб.: Санкт - Петербургский Государственный университет., 2010. — 103 с.
- Журавлёв А. Ю. До и после динозавров. — М.: Вече, 2006. — 352 с. — (Великие тайны). — ISBN 5-9533-1258-X.
