Завроподы
Завропо́ды[1][2], или зауропо́ды[3], или ящероно́гие[4] (лат. Sauropoda, буквально: ящероногие динозавры), — большая группа четвероногих растительноядных динозавров из отряда ящеротазовых, обитавших с конца триасового[5] по меловой периоды (около 210—66 миллионов лет назад) на всех материках, включая Антарктиду[6][7][8].
| † Завроподы | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: апатозавр, бронтозавр, диплодок; | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Инфраотряд: † Завроподы |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Sauropoda Marsh, 1878 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 200—66 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
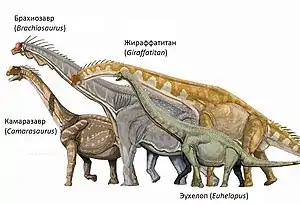
Завроподы насчитывают около 130 видов, до 13 семейств, около 70 родов. Наиболее известными представителями завроподов являются диплодоки, брахиозавры, апатозавры.
Отличительными особенностями их тела была очень длинная шея, длинные хвосты, маленькие головы (по отношению к остальной части их тела) и четыре толстые, похожие на колонны ноги.
Некоторые виды завропод в процессе эволюции достигли огромных размеров. В эту группу входят самые крупные животные, которые когда-либо жили на суше. Хорошо известные роды включают брахиозавра, диплодока, апатозавра, бронтозавра и мамэньсизавра.
Завроподы впервые появились в конце триасового периода[5], где они, по-видимому, входили в группу «Платеозавриды» или были их потомками.
К поздней юре (150 миллионов лет назад) завроподы стали широко распространенными на территории суши животными (особенно диплодоциды и брахиозавриды). В конце мела эти группы были в основном заменены титанозаврами, которые имели почти глобальное распространение.
Как и в случае с другими средними и крупными животными, титанозавры вымерли в процессе мел-палеогенового вымирания.
Название «завроподы» (Sauropoda) было придумано Отниелем Чарлзом Маршем в 1878 году[9].
Завроподы — одна из самых узнаваемых групп динозавров. Они стали неотъемлемой частью массовой культуры благодаря своим внушительным размерам.
Полные завроподобные ископаемые находки встречаются редко. Многие виды, особенно самые крупные, известны только из изолированных и вычлененных костей. У многих почти завершенных образцов отсутствуют головы, кончики хвоста и конечности. Самый крупный завропод титанозавр Бравипароп известен и описан на основании ихнофоссилий (отпечатков ног в виде вереницы следов).
Описание
Завроподы представляли собой гигантских четвероногих динозавров с небольшим по сравнению с общим размером черепом, массивным телом и длинной шеей, доходящей у некоторых видов до 9—11 метров. Ноздри смещены к глазницам. Зубы мелкие, шпателевидные.
Их эволюция демонстрирует феноменальные возможности адаптации к изменяющимся условиям, результатом чего является большое разнообразие этих животных. Размеры и вес гигантов являются предметом спора специалистов и имеют большой разброс оценок.
Череп
Завроподы отличаются относительно небольшой головой, около 1/200 по сравнению с общими размерами. По сравнению с фоссилиями других частей скелета, черепа завропод является довольно редкой находкой для палеонтологов. Известны менее одной трети черепов от общего количества родов завропод, и ещё меньше известны находки полного черепа. Поэтому каждая находка черепа завропода представляет для палеонтологов особый интерес. В результате открытия нового вида абидозавра в 2010 году, с отлично сохранившимся черепом, была подтверждена теория уменьшения величины зубов с ранней юры до мелового периода[10].
Конечности
Вопреки названию («ящероногие»), строением конечностей похожи не на ящериц, а на слонов (сходные адаптации к гигантизму). Более длинные передние, чем задние, конечности многих видов свидетельствуют о том, что пищу эти рептилии собирали преимущественно выше уровня туловища — кормились древесной листвой. Хотя другие виды могли кормиться водной и околоводной растительностью или в кустарниках.
Компьютерное моделирование локомоции завропод представлено в видеофайле 1[11].
Крестцовый мозг
Видоизменённые позвонки крестцового отдела позвоночника образовали подобие черепа. Крестцовый мозг превышал размерами головной в 20 раз.
Хвост
Хвост использовался в брачных ухаживаниях: самцы били хвостами о землю, создавая оглушающий (для слуха современного человека) грохот — самки добивался тот самец, кто издавал самый громкий звук. В точке хвоста, где скорость превышала скорость звука, позвонки для прочности срослись. Удары хвостом могли также использоваться для защиты от хищников.
Морфологические особенности
Многие завроподы, как, например, амаргазавр несли на себе всевозможные шипы, гребни и прочие «причудливые структуры», что визуально увеличивало их размер и, вероятно, служило для общения с сородичами — указывало на видовую принадлежность, могло сделать их внешне крупнее и/или сообщать о достижении рептилией половой зрелости. Для защиты от хищников большая часть «украшений», если не все, была непригодна.
Размеры и вес
Завроподы были гигантскими потомками удивительно маленьких предков — базальных динозавроморфов, таких как Pseudolagosuchus и Маразух из среднего триаса Аргентины, которые весили примерно 1 кг или меньше. Они превратились в ящеротазовых, которые претерпели быстрое увеличение размера на основе набора морфологических признаков, общих для многих представителей рептилий. И это несмотря на то, что более примитивные их представители, такие как Eoraptor, Panphagia, Pantydraco, Saturnalia и Guaibasaurus все еще сохранили свой умеренный размер и, соответственно, вес менее 10 кг[12].
Даже среди этих небольших примитивных динозавроморф имеются представители заметно увеличивших свой размер (скудные фоссилии этого периода делают интерпретацию предположительной). Существует один конкретный пример небольшого производного динозавроморфа: анхизавра, весом менее 50 кг. Он даже ближе к завроподам, чем платеозавр и Riojasaurus, вес которых превышал 1 т[12].
Завроподы были огромными. Их гигантский размер, вероятно, был обусловлен повышенной скоростью роста, которая стала возможной благодаря тахиметаболической эндотермии — физиологического свойства, которым по всей видимости обладали динозавроморфы. После выделения в отдельную группу, завроподы продолжали неуклонно увеличиваться в размерах. В ранней юре они были представлены барапазавром и котазавром. В средней юре — более громадными, например, мамэньсизавром и патагозавром. В ответ на рост завропод, хищные тероподы также увеличили свои размеры, как, например, целофизоид из Германии достиг размера аллозавра[12].
Причины расхождения оценок размеров и масс представителей мезозойской мегафауны заключается в применяемой методике.
Так, например, Кеннет Карпентер (Kenneth Carpenter) по сохранившемуся позвонку Амфицелия altus (см. Рис. 1.) — сверхлегкого диплодока Грега Пола (Gregory Scott Paul) весом 11,5 тонн — на основании принципа актуализма, а также используя аллометрический подход определяет высоту частично сохранившегося позвонка Амфицелия fragillimus — примерно 2,7 м, а затем последовательно и другие параметры: высоту тела в самой высокой точке спины — около 9,25 м, длину скелета — 58 м, а также массу тела — 122 400 кг (см. Рис. 1.)[13].

Майкл Тейлор (Michael P. Taylor) считает, что увеличение позвонка должно сопровождаться увеличением в процентном отношении объёмов пустот в материале кости и уменьшением толщины элементов скелета, что приводит, в свою очередь, к уменьшению ранее определённых линейных размеров. В таком случае частично сохранившийся позвонок Амфицелия fragillimus будет иметь уже 2,3 м в высоту, а в линейном измерении Амфицелия fragillimus уменьшится до длины 49 метров и, скорее всего, окажется весом «всего» в 78,5 тонны[14].
Ситуацию осложняет то, что большинство фоссилий завропод, найденных по всему миру, в том числе на территории стран бывшего СССР, представляют собой крайне фрагментированные фоссилии. Выбор методики, основанный на личных предпочтениях авторов исследований, генерирует, как, например, в случае Amphicoelias fragillimus, расхождение результатов на 35 %[12][14].
По всей видимости, для завропод была характерна экстремально высокая скорость роста. Так, изучение строения костей молодой особи Rapetosaurus krausei, найденных в формации Маэварана на Мадагаскаре, дало основания сделать вывод, что, имея при вылуплении массу около 3,4 кг, уже через несколько недель она достигла массы 40 кг (при высоте в бедре около 35 см)[15].
Ещё одним подтверждением глобального распространения завропод стали зубы, найденные в 90-х годах Аверьяновым А. О. и американскими палеонтологами в пустыне Кызылкум, в урочище Джаракудук, напоминающие по своему устройству зубы титанозавров — гигантских завропод массой в 100 тонн и длиной в 25—30 метров, чьи фоссилии часто находят на юге Аргентины, в Патагонии, и на севере Китая[16].
Преимущества и недостатки большого размера
Одними из причин огромных размеров некоторых завропод являются особенности их питания. Как известно, все завроподы были сухопутными существами и имели скудную, низкокалорийную диету, так как фоссилий из экосистем с постоянно влажным климатом не известно, наоборот, они обитали в засушливых, полупустынных регионах. Для того, чтобы извлечь как можно больше питательных веществ из такого рациона, необходимо как можно дольше задерживать пищу в пищеварительной системе. Таким образом, завроподы были вынуждены увеличивать длину своего кишечника, а следовательно и величину тела[17].
У животного огромных размеров есть и другие преимущества:
- Большой размер завропод был эффективным средством защиты от хищников[18].
- По сравнению с мелкими животными, более крупные виды имеют меньшую площадь поверхности кожи в соотношении с массой тела. Так как вода теряется через кожу, большому животному легче выжить в сухом климате. По этой же причине, если температура окружающей среды понижается, большое животное остывает медленнее, чем маленькое. Если температура повышается, большое животное менее подвержено перегревам. Когда температура воздуха колеблется, температура внутри массивного тела более стабильна и не подвержена колебаниям, чего нельзя сказать о мелких животных. Тело более крупного животного успешнее противостоит экстремальным условиям[18].
- Основная часть энергии, полученной из съеденной животным пищи, идёт на поддержание его жизнедеятельности, а меньшая — на осуществление двигательных функций. 100-килограммовый человек ест почти столько же, сколько 50-килограммовый. Амфицелию требовалось меньше пищи (в пересчете на объём тела), чем мелкому животному[18].
Однако, у крупного размера есть и ряд недостатков. Огромные животные наносят большой ущерб окружающей среде. Они буквально опустошают пастбища и затем перемещаются в новые места[18].
Образ жизни
Завроподы были растительноядными. Скорее всего, завроподы не пережевывали пищу, а глотали камни, чтобы перемолоть её непосредственно в желудке. Долгое время они считались полуводными животными[19], несмотря на полное отсутствие в скелете признаков, специфичных для водных животных[20]. Гипотетическое использование шеи в качестве шноркеля невозможно, так как при этом грудная клетка должна выдерживать давление воды[21].
По ранним версиям длинная шея завропод считалась приспособлением для кормления листьями высоких деревьев. Контраргументом такой гипотезы являются расчёты Р. Сеймура (2009)[22]: чтобы иметь возможность поднимать голову на высоту кроны дерева, артериальное давление в организме должно быть настолько высоким, что на его поддержание уходила бы приблизительно половина всей потребляемой завроподом энергии. Скорее всего, многие завроподы были стадными животными. В пользу этой гипотезы служит тот факт, что чаще всего их остатки палеонтологи находят группами. Завроподы не могли развивать скорость выше 5 км/ч вследствие большого веса, но возможно, что они могли прыгать на малую высоту, издавая колебания почвы и предупреждая сородичей об опасности в виде крупных хищных динозавров, таких как аллозавры. Завроподы могли защищаться от хищников при помощи хвоста, или просто затоптать хищника.
Происхождение и эволюция
Представления о ранней эволюции завропод и увеличении их разнообразия осложняются из-за скудной летописи окаменелостей вплоть до средней юры[23]. Учёные считают, что завроподы (как и другой подотряд динозавров — тероподы) появились на территории Гондваны (например, Antetonitrus, найденный в Южной Африке) и позже расселились по всем континентам. Благодаря ископаемым находкам известно, что завроподы 200 млн лет назад уже обитали на земле и развивались параллельно с прозавроподами[24]. Одними из самых ранних и примитивных завропод верхнетриасовой эпохи являются Isanosaurus, найденный в Таиланде, и вулканодон, найденный в Зимбабве, обитавшие по разные стороны океана Тетис. Это свидетельствует о том, что завроподы имели длинную эволюционную историю в течение верхнетриасовой эпохи.
Около 190 млн лет назад завроподы заселили территорию Индии (Barapasaurus, Kotasaurus), которая в то время была ещё частью континента Гондвана. Примерно в это же время они начали заселять другие территории, в частности территорию современной Южной Африки[24]. Ископаемые остатки говорят о том, что примерно 190—180 млн лет назад завроподы уже обитали в Китае (Zizhongosaurus, Kunmingosaurus), в Европе (Ohmdenosaurus), Марокко (Tazoudasaurus).
На территорию Северной Америки завроподы мигрировали значительно позднее. Известно, что в верхнетриасовую эпоху тероподы уже населяли территорию Северной Америки, однако следов завропод в этот период обнаружено не было. Единых мнений учёных по этому вопросу нет, некоторые высказывают предположение что их, скорее всего, не устраивал климат[25].
Ранняя эволюция завропод характеризовалась увеличением размеров тела, удлинением шеи и переходом от двуногого к четвероногому способу передвижения. Длина шеи завроподов превышает длину шеи всех других известных животных и является отличительным признаком эволюции группы, позволявшая им не только достигать большей высоты просмотра, но и покрывать более широкую зону кормления без необходимости перемещать свои массивные тела[23].
Наибольшего разнообразия завроподы достигли в среднеюрский период, дав множество различных видов. К концу юрского периода видовое разнообразие начинает несколько снижаться. В начале мелового периода (около 145—140 млн лет назад) завроподы уже населяли территорию почти всех континентов — Африки (брахиозавр), Северной Америки (апатозавр, диплодок), Европы (Xenoposeidon) и Азии (Qiaowanlong)[26]. В верхнемеловой эпохе происходит настоящий всплеск (особенно на южных континентах, где конкуренция с орнитоподами была минимальной). Основное разнообразие в этот период представляла группа титанозавров.
Находки
В Австралии в 2004 году были откопаны кости самого крупного завропода, найденного в Австралии, достигавшего 30 метров в длину. Позже учёные отнесли его к группе титанозавров. Учёные утверждают, что этот динозавр — представитель нового вида, которому пока ещё не дали названия[27].
В мартовском номере за 2011 год публикации Бразильской академии наук Anais da Academia Brasileira de Ciências напечатана статья[28], посвящённая находке в 2005 году международной палеонтологической экспедицией в Анголе, примерно в 70 километрах к северу от Луанды, фоссилий правой передней конечности нового крупного (около 13 м в длину) растительноядного динозавра-завропода, получившего наименование Angolatitan adamastor (Angolatitan — ангольский титан; adamastor — мифический гигант Адамастор, упоминаемый Л. Камоэнсом в «Лузиадах»).
Классификация
По данным сайта Fossilworks, на сентябрь 2017 года в инфраотряд включают следующие вымершие таксоны[29]:
- Роды incertae sedis
- Род Amygdalodon Cabrera, 1947
- Род Archaeodontosaurus Buffetaut, 2005
- Род Brontopodus Farlow et al., 1989
- Род Chinshakiangosaurus Yeh, 1975
- Род Chuxiongosaurus Lü et al., 2010
- Род Dachongosaurus Zhao, 1985
- Род Damalasaurus Zhao, 1985
- Род Eosauropus Lockley et al., 2006
- Род Isanosaurus Buffetaut et al., 2000
- Род Kotasaurus Yadagiri, 1988
- Род Lamplughsaura Kutty et al., 2007
- Род Lavinipes Avanzini et al., 2003
- Род Liujianpus Xing et al., 2016
- Род Microdontosaurus Zhao, 1983
- Род Morinosaurus Sauvage, 1874
- Род Ohmdenosaurus Wild, 1978
- Род Oplosaurus Gervais, 1852
- Род Oshanosaurus Zhao, 1985
- Род Parabrontopodus Lockley et al., 1995 (или в семействе Parabrontopodidae)
- Род Polyonyx Santos et al., 2009
- Род Pulanesaura McPhee et al., 2015
- Род Ravatichnus Gabuniya & Kurbatov, 1988
- Род Rotundichnus Hendricks, 1981
- Род Sanpasaurus Young, 1944
- Род Vulcanodon Raath, 1972 (или в семействе вулканодонтид)
- Семейство Faveoloolithidae Zhao & Ding, 1976
- Семейство Parabrontopodidae Apesteguía, 2005
- Род Breviparopus Dutuit & Ouazzou, 1980
- Клада Gravisauria Allain & Aquesbi, 2008
- Семейство Vulcanodontidae Cooper, 1984 — Вулканодонтиды[30]
- Клада Eusauropoda Upchurch, 1995
- Семейство Cetiosauridae Lydekker, 1888 — Цетиозавриды
- Семейство Mamenchisauridae Chao, 1965
- Клада Turiasauria Royo-Torres et al., 2006
- Клада Neosauropoda Wilson & Sereno, 1994
Ещё множество таксонов включают в инфраотряд в статусе nomen dubium, например, следующие роды: Aepisaurus Gervais, 1852, Asiatosaurus Osborn, 1924, Bruhathkayosaurus Yadagiri & Ayyasami, 1987, Campylodoniscus Kuhn, 1961, Chiayusaurus Bohlin, 1953, Chondrosteosaurus Owen, 1876, Gigantosaurus Seeley, 1869, Hamanosauripus Kim, 1986, Iguanodonichnus Casamiquela & Fasola, 1968, Koreanosauripus Kim, 1986, Kunmingosaurus Zhao, 1985, Lancanjiangosaurus Zhao, 1985, Nurosaurus Dong, 1992, Protognathosaurus Olshevsky, 1991, Qinlingosaurus Xie et al., 1996, Ultrasaurus Kim, 1981, Zizhongosaurus Dong et al., 1983[29].
Филогения
Данная кладограмма представляет собой упрощённую схему, предложенную в работе Джеффри Уилсона в 2002 году[31]).
| Sauropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- Завроподы / Алифанов В. Р. // Железное дерево — Излучение. — М. : Большая российская энциклопедия, 2008. — С. 158—159. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 10). — ISBN 978-5-85270-341-5.
- Михайлова, Бондаренко, 2006, с. 467.
- Зауроподы // Евклид — Ибсен. — М. : Советская энциклопедия, 1972. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 9).
- Геологический словарь / Всесоюзный научно-исследовательский геологический институт. — Госгеолтехиздат, 1960. — Т. 2. — С. 445.
- Lallensack et al., 2017.
- Wedel M. J., Cifelli R. L. _Oklahome’s Native Giant (англ.) // Oklahoma Geology Notes. — 2005. — No. 65 (2). — P. 40—57. — ISSN 0030-1735.
- Fernando E. Novas. The Age of Dinosaurs in South America. — 1st ed. — Indiana University Press, 2009. — P. 181. — 452 p. — ISBN 0253352894. — ISBN 978-0-253-35289-7.
- Oklahoma Geological Survey (англ.) // Oklahoma Geology Notes. — 2003. — P. 40. — ISSN 0030-1736.
- Marsh O. C. Principal characters of American Jurassic dinosaurs (англ.) // American Journal of Science. — 1878. — No. 16 (3). — P. 411—416. — ISSN 0002-9599.
- Daniel Chure, Brooks B. Britt, John A. Whitlock, Jeffrey A. Wilson. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition (англ.) // Naturwissenschaften. — 2010. — Iss. 97 (4). — P. 379—391. — doi:10.1007/s00114-010-0650-6. (недоступная ссылка)
- William Irvin Sellers, Lee Margetts, Rodolfo Aníbal Coria, Phillip Lars Manning. March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs (англ.) // PLOS ONE. — 2013. — 30 October (vol. 8, iss. 10). — P. e78733. — ISSN 1932-6203. — doi:10.1371/journal.pone.0078733.
- Sander et al., 2011.
- Carpenter K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878 (англ.) // New Mexico Museum of Natural History and Science Bulletin. — 2006. — No. 36. — ISSN 1524-4156. Архивировано 7 марта 2019 года.
- Taylor М. How big was Amphicoelias fragillimus? I mean, really? (англ.). Sauropod Vertebra Picture of the Week (19 февраля 2010). Дата обращения: 29 июня 2019.
- Kristina Curry Rogers, Megan Whitney, Michael D’Emic, Brian Bagley. Precocity in a tiny titanosaur from the Cretaceous of Madagascar (англ.) // Science. — 2016. — 22 April (vol. 352, iss. 6284). — P. 450—453. — doi:10.1126/science.aaf1509.
- Ученые из РФ и США определили самых крупных динозавров Средней Азии | РИА Новости
- Огромнейший из гигантских. www.dinomillennium.com. Дата обращения: 11 января 2017.
- Я познаю мир: Век динозавров: Энциклопедия. — М.: АСТ, 2002.
- Ефремов И. А. [iae.newmail.ru/science/Pr53-06/index.htm Вопросы изучения динозавров].
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 2. / Пер. с англ. — М.: Мир, 1993. — 283 с.
- Шмидт-Нильсен К. Размеры животных: почему они так важны? / Пер. с англ. — М.: Мир, 1987. — 259 с.
- Seymour R. S. 2009. Sauropods kept thier heads down: Science, v. 323, p. 1671.
- Kevin Leonel Gomez, Jose Luis Carballido, and Diego Pol. 2021. The axial skeleton of Bagualia alba (Dinosauria: Eusauropoda) from the Early Jurassic of Patagonia. Palaeontologia Electronica, 24(3): 37a
- Найдено переходное звено от двуногих к четырёхногим динозаврам. Lenta.ru (11 ноября 2009). Дата обращения: 13 августа 2010.
- Прояснены начальные моменты эволюции динозавров — Наука и техника — История, археология, палеонтология — Палеонтология — Компьюлента Архивная копия от 17 февраля 2010 на Wayback Machine
- Палеонтологи нашли новый вид зауроподов. Lenta.ru (8 сентября 2009). Дата обращения: 13 августа 2010.
- Титанозавры из Австралии
- Octávio Mateus, Louis L. Jacobs, Anne S. Schulp, Michael J. Polcyn, Tatiana S. Tavares, André Buta Neto, Maria Luísa Morais and Miguel T. Antunes. Angolatitan adamastor, a new sauropod dinosaur and the first record from Angola (англ.) // Anais da Academia Brasileira de Ciências (Annals of the Brazilian Academy of Sciences). — 2011. — March (iss. 83 (1)). — P. 1—13. — ISSN 0001-3765.
- †Sauropoda (англ.) информация на сайте Fossilworks. (Дата обращения: 12 октября 2017).
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 118. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Wilson, 2002.
Литература
- Михайлова И. А., Бондаренко О. Б. Палеонтология. — 2 изд., перераб. и доп. — М.: Издательство МГУ, 2006. — 592 с. — ISBN 5-211-04887-3.
- Даррен Нэйш, Пол Барретт. Динозавры: 150 000 000 лет господства на Земле = Dinosaurs: How They Lived and Evolved / переводчик Рыбаков К., научный редактор Аверьянов А., д-р биол. наук. — М.: Альпина нон-фикшн, 2018. — 223 с. — ISBN 978-5-0013-9010-7.
- Jens N. Lallensack, Hendrik Klein, jesper Milàn, Oliver Wings, Octávio Mateus, Lars B. Clemmensen. Sauropodomorph dinosaur trackways from the Fleming Fjord Formation of East Greenland: Evidence for Late Triassic sauropods (англ.) // Acta Palaeontologica Polonica. — 2017. — No. 62 (4). — P. 833—843. — ISSN 1732-2421. — doi:10.4202/app.00374.2017.
- P. Martin Sander, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva‐Maria Griebeler, Hanns‐Christian Gunga, Jürgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas Tütken, Oliver Wings, Ulrich Witzel. Biology of the sauropod dinosaurs: the evolution of gigantism (англ.) // Biological Reviews : journal. — 2011. — 20 January (no. 86 (1)). — P. 117–155. — ISSN 1464-7931. — doi:10.1111/j.1469-185X.2010.00137.x. — PMID 21251189.
- Jeffrey A. Wilson. Sauropod dinosaur phylogeny: critique and cladistic analysis (англ.) // Zoological Journal of the Linnean Society. — 2002. — Iss. 136. — P. 217—276.
| Таксономия |
|---|