Апатозавр
Апатоза́вр[1] (лат. Apatosaurus) — род крупных динозавров-завропод из семейства диплодоцид, живших во времена позднеюрской эпохи (157,3—145,0 млн лет назад) на территории современной Северной Америки. В род включают 2 вида: типовой Apatosaurus ajax, названный и описанный Отниелом Чарлзом Маршем в 1877 году, и Apatosaurus louisae, названный и описанный в 1916 году Уильямом Холландом. Ископаемые остатки найдены в геологической формации Моррисон, расположенной на территории штатов Колорадо, Оклахома, Нью-Мексико, Вайоминг и Юта в Соединённых Штатах Америки. Средняя длина животного составляла 21—22,8 метра, а средняя масса — 16,4—22,4 тонны. Несколько образцов показывают максимальную длину на 11—30 % больше средней и массу 32,7—72,6 тонн.
| † Апатозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Инфраотряд: Надсемейство: Семейство: Подсемейство: Род: † Апатозавр |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Apatosaurus Marsh, 1877 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Виды | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 155,7—145,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||


Шейные позвонки апатозавра короче и тяжелее шейных позвонков родственного диплодока, а кости ног — толще и длиннее, что подразумевает более крепкое телосложение апатозавра. Во время движения хвост животного держался над землёй. Каждая передняя конечность несла один коготь, а каждая задняя — три. Апатозавр питался листвой деревьев, держа голову высоко поднятой. Позвонки этого животного имели обширные полости, заполненные воздушными мешками, облегчавшие вес костей. Как у многие другие диплодоциды, апатозавр, возможно, мог щёлкать кончиком своего длинного хвоста, как кнутом.
До 1909 года череп апатозавра был неизвестен, и скелеты дополняли высоким, коротким и толстокостным черепом камаразавра. После обнаружения скелетного материала второго вида, Apatosaurus louisae, оказалось, что череп апатозавра схож с низким и тонким черепом диплодока. Некоторые более крепкие черепа, ассоциированные с диплодоком, могли на самом деле оказаться черепами апатозавров.
Систематики размещают апатозавра в семействе диплодоцид, в пределах подсемейства Apatosaurinae; это один из наиболее базальных родов. Хотя подсемейство Apatosaurinae было введено в систематику ещё в 1929 году, его должным образом не использовали до тщательного исследования завропод 2015 года. Сюда же включают бронтозавра, а другие роды считают синонимами или переклассифицируют как представителей подсемейства Diplodocinae. Живший в поздней юре апатозавр делил свою среду обитания с такими динозаврами, как аллозавр, камаразавр, диплодок и стегозавр.
Описание
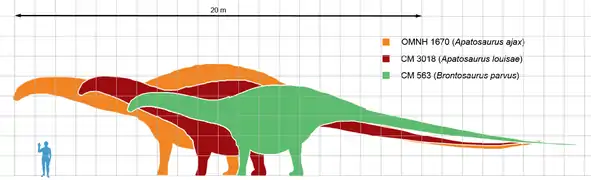
Апатозавр был крупным четвероногим животным с длинной шеей и длинным хвостом, похожим на кнут. Его передние конечности были немного короче задних. Большинство оценок размера основаны на образце CM 3018 — типовом образце A. louisae. В 1936 году путём замера позвоночника длину животного определили как составляющую 21,8 метра[2]. Современные оценки близки к этому значению: 21–22,8 метра в длину с массой от 16,4 до 22,4 тонн[3][4][5]. В исследовании 2015 года, где была оценена масса объёмных моделей Dreadnoughtus, апатозавра и жираффатитана, массу образца CM 3018 оценили в 21,8–38,2 тонны, аналогично массе Dreadnoughtus[6]. Некоторые экземпляры A. ajax (например, OMNH 1670) представляют особей на 11–30 % длиннее, что говорит о том, что их масса была в 2 раза больше массы CM 3018 или 32,7–72,6 тонн, что потенциально может составить конкуренцию самым крупным титанозаврам[7].

Череп апатозавра мал по сравнению с общим размером животного, челюсти оснащены долотоподобными зубами, подходящими для растительной пищи[8]. Форма морды была квадратной, и только у нигерзавра была морда ещё более квадратная[9]. Черепная коробка хорошо сохранилась в экземпляре BYU 17096, который также сохранил большую часть скелета. Филогенетический анализ показал, что черепная коробка апатозавра схожа с таковой у других диплодокоидов[10]. Некоторые черепа найдены вместе с зубами. На тех зубах, которые сохранили открытую эмалевую поверхность, отсутствуют царапины; вместо этого они показывают гладкую структуру и небольшой износ[9].

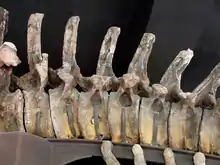
Как и у других завропод, шейные позвонки апатозавра были сильно разнесёнными; они несли невральные дуги с большой впадиной посередине, результатом чего являлась широкая, глубокая шея[8]. Голотип Apatosaurus louisae содержит 15 шейных, 10 грудных, 5 крестцовых и 82 хвостовых позвонка. Число хвостовых позвонков могло варьироваться даже в пределах вида. Шейные позвонки апатозавра и бронтозавра намного толще и крепче таковых у всех прочих диплодоцид; по мнению Чарльза Гилмора, они выглядят больше похожими на шейные позвонки камаразавра[2][11]. Кроме того, они поддерживают шейные рёбра, которые тянутся вниз дальше, чем у диплодоцин. В верхней части шейные позвонки и шейные рёбра сужаются, делая шею животного почти треугольной в поперечном сечении[11]. Первый и второй шейные позвонки Apatosaurus louisae почти слиты. Грудные рёбра были прикреплены к позвонкам неплотно, они свободно сочленены[2]. С каждой стороны тела апатозавр имел по 10 рёбер[12]. Большую шею заполняла обширная система воздушных мешков для снижения массы. У апатозавра, как и у его близкого родственника суперзавра, были высокие невральные дуги, высота которых составляет более половины высоты отдельных позвонков. Форма хвоста необычна для диплодоцид; он сравнительно тонкий из-за быстро уменьшающейся высоты невральных дуг с увеличением расстояния от бёдер. Также у апатозавра были очень длинные рёбра по сравнению с большинством других диплодоцид, из-за чего его грудная клетка была необычайно глубокой[13]. Как и у прочих диплодоцид, конец хвоста был кнутообразным[2].
_(Morrison_Formation%252C_Upper_Jurassic%253B_Carnegie_Quarry%252C_Dinosaur_National_Monument%252C_Utah%252C_USA)_1_(48696375026).jpg.webp)
Кости конечностей апатозавра также очень крепкие[13]. Среди представителей подсемейства Apatosaurinae морфология лопатки Apatosaurus louisae является промежуточной по форме между Apatosaurus ajax и Brontosaurus excelsus. Крепкая плечевая кость апатозавра напоминает таковые у камаразавра и бронтозавра. Однако плечевые кости бронтозавра и Apatosaurus ajax больше похожи друг на друга, чем на Apatosaurus louisae. В 1936 году Чарльз Гилмор отметил, что предыдущие реконструкции передних конечностей апатозавра ошибочно предполагали, что лучевая и локтевая кости могли пересекаться; на самом деле они были параллельными[2]. На каждой передней конечности апатозавра был большой коготь — особенность, характерная для всех завропод, более продвинутых, чем шунозавр[2][14]. Первые три пальца задней конечности несли когти. Фаланговая формула 2-1-1-1-1 означает, что самый внутренний палец на передней конечности имеет две фаланги, а все следующие имеют одну[15]. Единственный коготь, расположенный на каждой передней конечности, слегка изогнут и имеет квадратное сечение на переднем конце. Тазовый пояс включает в себя массивные подвздошные кости, слитые лобковые и седалищные кости. Бедренная кость апатозавра очень крепкая и является одной из самых крепких бедренных костей среди всех известных у завропод. Большая и малая берцовые кости отличаются от более тонких костей диплодока, но почти не отличаются от костей камаразавра. Малоберцовая кость длиннее и тоньше большеберцовой. Ступня апатозавра несёт 3 когтя на самых внутренних пальцах; фаланговая формула 3-4-5-3-2. Первая плюсневая кость — самая крепкая, что является характерной чертой диплодоцид[2][16].
Открытие и название
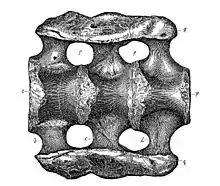
Первые окаменелости апатозавра обнаружил Эдвард Дринкер Коп, а название Apatosaurus ajax ввёл в систематику его конкурент и противник по «костяным войнам» Отниел Чарлз Марш, профессор палеонтологии в Йельском университете, в 1877 году. Вид был описан на основании голотипа YPM 1860, почти полного скелета, обнаруженного в предгорьях Скалистых гор в округе Ганнисон, штат Колорадо (США)[12][17][18]. Родовое название Apatosaurus составлено из греч. apatē (ἀπάτη)/apatēlos (ἀπατηλός) — обман, обманчивый, и sauros (σαῦρος) — ящер[19]. Марш дал животному такое название из-за формы шевронов, не похожих на шевроны других динозавров; вместо этого шевроны апатозавра имели сходство с шевронами мозазавров[17][20]. Во время раскопок и транспортировки кости голотипа смешали с костями другой особи апатозавра, первоначально описанной как Atlantosaurus immanis; как следствие, некоторые элементы нельзя с уверенностью приписать ни одному из образцов[18]. Марш проводил различие между родами Apatosaurus и Atlantosaurus на основании количества крестцовых позвонков (у апатозавра их было 3, у Atlantosaurus — 4). Два года спустя Марш объявил об открытии более крупного и более полного образца в Комо-Блафф, штат Вайоминг. Он дал этому образцу новое название, основанное на разнице его геологического возраста и относительно редких окаменелостях, доступных в то время. Позже было признано, что особенности, которые он использовал для различения родов и видов, на самом деле были широко распространены среди завропод[12][18]. Новый вид он назвал Brontosaurus excelsus[21]. Все экземпляры, которые в настоящее время считаются апатозаврами, происходили из формации Моррисон, где вели раскопки Марш и Коп[22].

Другой экземпляр в Американском музее естественной истории под номером 460, который иногда присваивают апатозавру, считается почти завершенным; отсутствуют только череп, ноги и участки хвоста. Это первый смонтированный скелет зауропода[23]. Образец нашёл в 1898 году к северу от города Медисин-Боу (Вайоминг) американский палеонтолог Уолтер Грейнджер. На извлечение скелета ушло всё лето[24]. К смонтированному скелету были добавлены ступни завропода, обнаруженные в том же карьере, и хвост, позвонки которого были отлиты так, как представлял Марш; при этом позвонков оказалось слишком мало. Кроме того, была сделана скульптурная модель того, как, по мнению музея, мог выглядеть череп этого массивного существа. Это был не тонкий череп, подобный черепу диплодока, который позже был признан более точным, но основывался на «самых больших, самых толстых, самых сильных костях черепа, нижних челюстях и зубных коронках из трёх разных карьеров»[2][12][23][25]. Эти черепа были, вероятно, черепами камаразавров — единственных других завропод, для которых в то время был известен полный черепной материал. За монтажом скелета наблюдал Адам Германн, который не смог найти череп апатозавра. Герману пришлось лепить череп вручную. В своей публикации Осборн заметил, что череп был «в значительной степени предположительным и основанным на черепе Morosaurus» (ныне камаразавра)[26].

В 1903 году американский палеонтолог Элмер Риггс опубликовал исследование, в котором описал хорошо сохранившийся скелет диплодока, найденный в долине Гранд-Ривер близ Фруты, штат Колорадо, экземпляр Филдовского музея P25112. Риггс считал, что по возрасту это месторождение было похоже на месторождение Комо-Блафф в Вайоминге — найденного там бронтозавра описал Марш. Была найдена бо́льшая часть скелета, и после сравнения как с бронтозавром, так и с апатозавром, Риггс понял, что голотип A. ajax принадлежал неполовозрелой особи, и, следовательно, признаки, по которым диагностируется род, были недействительными. Поскольку название «апатозавр» было дано раньше, название «бронтозавр» следует считать младшим синонимом. Из-за этого Риггс рекомбинировал вид Brontosaurus excelsus в Apatosaurus excelsus. На основе сравнения с другими видами, предположительно принадлежащими к апатозаврам, палеонтолог также определил, что образец, находящийся в Филдовском музее, вероятно, является A. excelsus[12].
Несмотря на публикацию Риггса, Генри Осборн, бывший непримиримым противником Марша и его таксонов, отметил скелет апатозавра, находящийся в Американском музее естественной истории, как бронтозавра[26][27]. Из-за этого решения название «бронтозавр» обычно использовалось за пределами научной литературы для того животного, которого Риггс считал апатозавром, а популярность музея повлияла на то, что бронтозавр стал одним из самых известных динозавров, несмотря на то, что его название было недействительным в течение почти всего XX и начала XXI веков[28].
.jpg.webp)
Череп апатозавра был найден только в 1909 году во время экспедиции, возглавляемой Эрлом Дугласом, в так называемом «карьере Карнеги», расположенном на территории Национального памятника Динозавр. Череп был обнаружен на небольшом расстоянии от скелета (образец CM 3018), идентифицированного как новый вид Apatosaurus louisae, названный в честь Луизы Карнеги, жены Эндрю Карнеги, которая финансировала полевые работы. Череп был обозначен индексом CM 11162 и оказался очень похож на череп диплодока[27]. Рядом с образцами CM 11162 и CM 3018 был найден другой, меньший скелет A. louisae[29]. Эрл Дуглас и директор Музея Карнеги Уильям Холланд признали найденный череп принадлежащим образцу апатозавра, хотя другие учёные, в особенности Осборн, отвергли эту идентификацию. Холланд отстаивал свою точку зрения в 1914 году в своём обращении к Палеонтологическому обществу Америки, но, тем не менее, оставил скелет в Музее Карнеги без головы. В то время как некоторые думали, что директор музея таким образом пытается избежать конфликта с Осборном, другие считали, что Холланд ждёт, пока не найдут сочленённый скелет апатозавра с черепом[26]. После смерти Холланда в 1934 году сотрудники музея поместили на скелет череп камаразавра[27].
Пока большинство музеев оснащало скелеты апатозавров слепками или отливками черепов камаразавров, сотрудники Йельского музея Пибоди решили слепить череп на основе нижней челюсти камаразавра с черепом, основанным на иллюстрации Марша 1891 года. Этот череп имел направленные вперёд ноздри — необычные для любого динозавра — и окна, отличающиеся как от окон на иллюстрации, так и от окон других черепов[26].

Черепа апатозавров не упоминались в научной литературе до 1970-х годов, когда Джон Стэнтон Макинтош и Дэвид Берман заново описали черепа диплодока и апатозавра. Они обнаружили, что Холланд, хотя он никогда не публиковал свое мнение, почти наверняка был прав в том, что у апатозавра был череп, похожий на таковой у диплодока. По их мнению, многие черепа, которые, как долго думали, принадлежали диплодокам, могут быть черепами апатозавров. Они заново отнесли несколько черепов к апатозавру на основе связанных и тесно связанных позвонков. Несмотря на то, что авторы исследования поддерживали мнение Холланда, они также отметили, что апатозавр мог обладать черепом, похожим на череп камаразавра. Такой вывод был сделан на основе единственного отдельного зуба, похожего на зуб камаразавра, найденного в том же месте, где за много лет до этого был найден образец апатозавра[25]. 20 октября 1979 года, после публикаций Макинтоша и Бермана, в музее Карнеги к скелету апатозавра был добавлен первый настоящий череп[27]. И только в 2011 году был найден череп апатозавра, сочленённый с шейными позвонками. Было обнаружено, что этот образец, CMC VP 7180, отличается как от черепа, так и от шеи A. louisae, но разделяет многие особенности шейных позвонков с A. ajax[30]. Другим хорошо сохранившимся черепом является образец 17096 Университета Бригама Янга, хорошо сохранившийся скелет с черепом, в том числе, с сохранившейся черепной коробкой. Образец был найден в карьере Кактус Парк в западном Колорадо (США)[10].
Почти все палеонтологи согласились с Риггсом, что таксоны апатозавр и бронтозавр должны классифицироваться как один род. Согласно правилам МКЗН (который регулирует научные названия животных), название апатозавр, которое было опубликовано первым, имеет приоритет в качестве официального названия; название бронтозавр считалось младшим синонимом и поэтому долго не использовалось[31][32][33][34]. Несмотря на это, по крайней мере один палеонтолог — Роберт Т. Бэккер — в 1990-х годах утверждал, что A. ajax и A. excelsus («Brontosaurus» excelsus) на самом деле достаточно различны, и что последний заслуживает выделения в отдельный род[35].
В 2015 году Эмануэль Чопп, Октавио Матеус и Роджер Бенсон опубликовали статью о систематике диплодокоидов и предположили, что роды можно диагностировать по тринадцати различным признакам, а виды — по шести. Минимальное число для разделения по родам выбрано на основании того факта, что A. ajax и A. louisae различаются двенадцаться характеристиками, а Diplodocus carnegiei и D. hallorum — одиннадцатью. Таким образом, было выбрано тринадцать отличительных черт для подтверждения разделения родов. Шесть отличительных признаков для видового разделения учёные выбрали путём подсчёта количества отличающихся признаков в отдельных экземплярах, которые однозначно представляют один вид, с одним отличительным признаком у D. carnegiei и A. louisae, и по пяти признакам у B. excelsus. Поэтому авторы статьи утверждали, что Apatosaurus excelsus, первоначально классифицированный как Brontosaurus excelsus, имел достаточно морфологических отличий от других видов апатозавров, чтобы оправдать свою повторную классификацию в качестве отдельного рода. Такой вывод был сделан по результатам сравнения 477 морфологических признаков у 81 экземпляра динозавров. Одним из многих заметных отличий являлась более широкая и предположительно более сильная шея видов апатозавра по сравнению с B. excelsus. Другие виды, ранее относимые к апатозавру, такие как Elosaurus parvus и Eobrontosaurus yahnahpin, также были реклассифицированы как бронтозавр. Некоторые особенности, предлагаемые для отделения бронтозавра от апатозавра, включают: задние грудные позвонки с центрумом более длинным, чем широким; акромиальный край лопатки имеет округлое расширение; отношение длины к ширине астрагала 0,55 или более[18]. Эксперт по завроподам Мишель Даниэль д`Эмик отметил, что выбранные критерии были в некоторой степени произвольными и что они потребовали бы повторного отказа от названия бронтозавр, если в результате новых анализов будут получены другие результаты[36]. Специалист по вымершим млекопитающим Дональд Протеро раскритиковал реакцию средств массовой информации на это исследование, назвав её поверхностной и преждевременной и добавив, что он будет заключать название «бронтозавр» в кавычки и не будет относиться к нему как к валидному роду[37].
Валидные виды



Многие виды апатозавра определены по весьма скудному материалу. Отниэл Марш назвал столько видов, сколько мог, однако многие из них были основаны на фрагментарных и недиагностируемых остатках. В 2005 году Пол Апчёрч и его коллеги опубликовали исследование, в котором анализировались виды и видовые связи апатозавра. Они обнаружили, что самым основным видом был A. louisae, за которым следуют FMNH P25112, а затем — политомия из A. ajax, A. parvus и A. excelsus[16]. В 2015 году их анализ был пересмотрен и расширен многими дополнительными видами, которые несколько по-разному разрешили филогенетические взаимоотношения апатозавра, а также поддержали отделение бронтозавра от апатозавра[18].
- Apatosaurus ajax был назван Маршем в 1877 году в честь Аякса — героя из греческой мифологии[38]. В качестве голотипа Марш назначил неполный скелет неполовозрелой особи, образец YPM 1860. Этот вид менее изучен, чем бронтозавр и A. louisae, в первую очередь, из-за неполноты голотипа. В 2005 году было обнаружено, что, помимо голотипа, к A. ajax можно отнести многие экземпляры: YPM 1840, NSMT-PV 20375, YPM 1861 и AMNH 460. Образцы датируют промежутком времени от позднего киммериджа до раннего титона[16]. Исследование 2015 года отнесло к этому виду только голотип YPM 1860, а образец AMNH 460 был восстановлен либо как бронтозавр, либо как собственный таксон. Тем не менее, YPM 1861 и NSMT-PV 20375 отличались только по нескольким характеристикам и не могут отличаться как род или вид от A. ajax. Экземпляр YPM 1861 является голотипом «Atlantosaurus» immanis, что означает, что он может быть младшим синонимом A. ajax[18].
- Apatosaurus louisae ввёл в систематику Холланд в 1916 году на основании неполного скелета, найденного в штате Юта (США)[39]. Голотипом является образец CM 3018, с отнесёнными образцами CM 3378, CM 11162 и LACM 52844. Первые два экземпляра представляют собой позвоночные столбы; последние два — череп и почти полный скелет соответственно. Все экземпляры A. louisae были найдены в киммериджских отложениях на территории Национального памятника Динозавр[16]. В исследовании 2015 года Э. Чопп и его коллеги обнаружили, что вид Apatosaurus laticollis имеет очень много сходных черт с образцом CM 3018, что означает, что первый, вероятно, является младшим синонимом A. louisae[18].
Приведённая ниже кладограмма является результатом филогенетического анализа, проведённого в 2015 году командой палеонтологов под руководством Э. Чоппа. Авторы проанализировали большинство образцов джиплодоцид отдельно друг от друга, чтобы определить, какой экземпляр принадлежал к какому виду и роду[18].
| Apatosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Переназначенные виды

- Apatosaurus grandis был назван О. Маршем в 1877 году в той же статье, в которой он описал A. ajax. Краткое описание было снабжено иллюстрацией и диагнозом[2]. Годом позже Марш упомянул, что вид был отнесён к апатозавру временно, и причислил его к новому роду Morosaurus[40]. Поскольку Morosaurus считается синонимом камаразавра, Camarasaurus grandis является самым старым из названных видов этого рода[41].
- Apatosaurus excelsus был первоначально типовым видом бронтозавра, впервые названным Маршем в 1879 году. В 1903 году Элмер Риггс реклассифицировал название бронтозавр как синоним апатозавра, перенеся вид B. excelsus в A. excelsus. В 2015 году Чопп, Матеус и Бенсон утверждали, что этот вид имеет достаточно различий, чтобы поместить его в отдельный род, поэтому они реклассифицировали его обратно в бронтозавра[18].
- Apatosaurus parvus, впервые описанный в 1902 году по скелету неполовозрелой особи как Elosaurus Петерсоном и Гилмором, был был реклассифицирован как апатозавр в 1994 году, а затем как бронтозавр — в 2015 году. Множество других, более зрелых экземпляров отнесены к нему после исследования 2015 года[18].
- Apatosaurus minimus в 1904 году описал Г. Осборн как образец бронтозавра. В 1917 году Генри Мук отнёс его к отдельному виду апатозавра, A. minimus. Таксон основан на образце, состоящем из пары подвздошных костей, сочленённых с крестцом[2][42][43]. В 2012 году Майк Тэйлор и Мэтт Уэдел опубликовали краткую аннотацию, описывающую материал A. minimus, и обнаружили, что его сложно отнести как к диплодокоидам, так и к макронариям. Одно время вид помещали в род сальтазавр, поскольку образец представлял некоторую форму со сходящимися чертами из многих групп[43]. Обширное исследование 2015 года подтвердило расположение вида в пределах семейства Camarasauridae, однако, авторы работы оговорились, что позиция таксона сильно изменчива, и позиции более явной они не нашли[18].
- Apatosaurus alenquerensis назвал в 1957 году французские палеонтологи Альбер-Фели де Лаппарен и Жорж Збивески на основе посткраниального материала из Португалии. В 1990 году этот материал отнесли к камаразавру, но в 1998 году ему было присвоено собственное название Lourinhasaurus[16]. Выделение в отдельный род подтвердило исследование 2015 года, где было установлено, что Lourinhasaurus является сестринским таксоном для камаразавра и других камаразаврид[18].
- Apatosaurus yahnahpin назвали и описали Джеймс Филла и Патрик Редман в 1994 году. В 1998 году Р. Бэккер сделал A. yahnahpin типовым видом для нового рода Eobrontosaurus[35], а в 2015 году Чопп и его коллеги классифицировали животное как Brontosaurus yahnahpin[18].
Систематика

Апатозавр является представителем семейства диплодоцид — группы гигантских завроподовых динозавров. В это семейство входят одни из самых длинных существ, когда-либо ходивших по земле, в том числе диплодок, суперзавр и барозавр. Апатозавра иногда включают в подсемейство Apatosaurinae, которое также может содержать Suuwassea, суперзавра и бронтозавра[13][44][45]. О. Ч. Марш описал апатозавра вместе с Atlantosaurus в составе ныне не существующей группы Atlantosauridae[12][17]. В 1878 году Марш поднял это семейство в ранге до подотряда, включив в него, помимо апатозавра и Atlantosaurus, диплодока и Morosaurus (=Camarasaurus). Он классифицировал эту группу в составе группы Sauropoda, которую ввёл в систематику в этой же научной работе. В 1903 году Элмер С. Риггс сказал, что название Sauropoda будет младшим синонимом более ранних названий; он сгруппировал апатозавра в пределах группы Opisthocoelia[12]. В 2011 году Джон Уитлок опубликовал исследование, в котором апатозавр был назван более базальным диплодоцидом, менее базальным, чем суперзавр[46].
Ниже приводится кладограмма для семейства диплодоцид, построенная в 2015 году по результатам филогенетического анализа, проведённого группой палеонтологов под руководством Э. Чоппа[18].
| Diplodocidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
В течение всего XIX и начала XX веков считалось, что такие завроподы, как апатозавр, были слишком массивными, чтобы выдержать собственный вес на суше. Предполагалось, что они жили частично погруженными в воду, возможно, в болота. Более поздние результаты не подтверждают эту гипотезу; считается, что завроподы были полностью наземными животными[47]. Исследование черепов диплодоцид показало, что квадратная морда, большое количество ямок и мелкие субпараллельные царапины на зубах апатозавра позволяют предположить, что он был неразборчивым собирателем, питавшимся на уровне земли[9]. Он мог есть папоротники, цикадовые, семенные папоротники, хвощи и водоросли[48]. Некоторые исследователи предположили, что апатозавры паслись по берегам рек, поедая подводные растения[49].
Проведённое в 2015 году исследование шей апатозавра и бронтозавра выявило множество различий между ними и другими диплодоцидами. Возможно, данные различия указывают на использование этими животными шей при внутривидовых боях[11]. Были предложены различные варианты использования единственного когтя на передних конечностях зауропод. Одно из предположений состоит в том, что они использовались для защиты, но их форма и размер делают это маловероятным. Наиболее вероятным использованием когтей было захватывание с их помощью таких предметов, как стволы деревьев, при кормлении[14].
Следы крупных завропод, таких, как апатозавр, показывают, что эти динозавры могли проходить по 25—40 километров в день, двигаясь со скоростью 20—30 километров в час[8]. Сравнительно медленное передвижение завропод может быть связано с их минимальной мускулатурой или с отдачей от шагов[50]. Следы, оставленные неполовозрелыми завроподами, заставили некоторых учёных утверждать, что эти животные были способны к бипедальности, однако, эти выводы оспариваются[51][52].
Положение шеи

Диплодоцид, таких как апатозавр, часто изображают с высоко поднятыми шеями, позволявшими им общипывать высокие деревья. В некоторых исследованиях утверждается, что шеи диплодоцид были менее гибкими, чем считалось ранее, потому что структура шейных позвонков не позволила бы шее вытягиваться высоко вверх, и что завроподы, такие как апатозавр, были приспособлены к поеданию растительности с уровня земли[48][49][53].
Проведённые Майклом Тейлором исследования показали, что все четвероногие, по-видимому, держат шеи на максимально возможном вертикальном разгибании, когда они находятся в нормальном положении; учёный утверждает, что то же самое будет справедливо и для завропод, за исключением любых неизвестных, уникальных характеристик, которые отличают анатомию мягких тканей их шеи от других животных. Апатозавр, как и диплодок, держал бы свою шею под наклоном вверх, а в положении покоя голова была бы направлена вниз[54][55]. Кент Стивенс и Майкл Пэрриш утверждают, что шея апатозавра имела больший диапазон движений и могла сгибаться в U-образную форму при виде сбоку[48]. Такой диапазон движения шеи позволил бы животному питаться на уровне собственных ступней[49].
Мэттью Кобли с соавторами оспаривают эти утверждения, заявляя, что крупные мышцы и хрящи ограничивали бы движения шеи. В свою очередь, они утверждают, что диапазон движений шей завропод, таких как диплодок, был меньше, чем считалось ранее, и животным, возможно, приходилось перемещать всё своё тело, чтобы добраться до областей, где они могли бы объедать растительность. Таким образом, диплодоциды тратили больше времени на поиск пищи, чтобы удовлетворить свои минимальные потребности в энергии[56][57]. Выводы Кобли оспаривает Тейлор, который проанализировал количество и расположение межпозвонковых хрящей, чтобы определить гибкость шей апатозавра и диплодока. Он обнаружил, что шея апатозавра была очень гибкой[54].
Физиология
Учитывая огромную массу тел и длинные шеи завропод, физиологи строят предположения, какой была дыхательная система этих животных. Выдвигались гипотезы, что, как и у крокодилов, у апатозавров не было диафрагмы, и потому количество мёртвого пространства (объём неиспользованного воздуха, остающегося во рту, трахее и бронхах после каждого вдоха) оценивался примерно в 0,184 м³ для образца массой 30 тонн. Группа исследователей под руководством Ф. Паладино оценила дыхательный объём (количество воздуха, перемещаемого внутрь или наружу во время одного вдоха) равный 0,904 м³, при условии, что дыхательная система этих динозавров была подобна птичьей, 0,225 м³ — если подобна таковой у млекопитающих, и 0,019 м³ — если у рептилий[58].
Исходя из этих расчётов, дыхательная система апатозавра, скорее всего, напоминала птичью, с воздушными мешками и проточными лёгкими. Для дыхательной системы птичьего типа потребовался бы объём лёгких, равный 0,6 м³ (600 литров), против 2,95 м³ (2950 л) для дыхательной системы как у млекопитающих, которая превышает допустимое пространство внутри грудной клетки. Общий объём грудной клетки апатозавра оценили в 1,7 м³ (1700 литров), что позволяло вместить четырёхкамерное сердце объёмом 0,5 м³ (500 литров) и лёгкие объёмом 0,9 м³ (900 литров)[58]. Доказательством наличия у завропод и других динозавров дыхательной системы птичьего типа служит пневматизация позвонков. Хотя пневматизация снижает общий вес животного, считается, что в полостях располагались воздушные мешки, как у птиц[59].
В 1991 году Джеймс Спотила с соавторами пришёл к выводу, что большой размер завропод не позволил бы им поддерживать высокую скорость метаболизма, потому что они не смогли бы выделять достаточно тепла[60]. Они предположили, что у завропод была «рептильная» дыхательная система. Палеонолог Мэттью Уэдел утверждал, что «птичья» дыхательная система позволила бы этим животным сбрасывать больше тепла[59]. Некоторые учёные утверждают, что у сердца завропода были бы проблемы с поддержанием достаточного артериального давления, чтобы насыщать мозг кислородом[47]. Другие предполагают, что почти горизонтальная осанка головы и шеи устранила бы проблему подачи крови в мозг, потому что таким образом давление не было бы высоким[48].
В 1987 году Джеймс Фарлоу подсчитал, что пищеварительная система 35-тонного апатозавра вмещала бы 5,7 тонн перебродившего содержимого[61]. Предположив, что апатозавр имел дыхательную систему птиц и метаболизм рептилий, Фрэнк Паладино с соавторами подсчитали, что животное должно было потреблять всего около 262 литров воды в день[58].
Модели роста

Микроскопическое исследование костей апатозавра и бронтозавра в 1999 году показало, что эти животные быстро росли в молодости и примерно за 10 лет достигли размеров, близких к взрослым[62]. В 2008 году Томас Леман и Холли Вудворд опубликовали исследование скорости роста завропод. Они показали с использованием линий роста и соотношения длины тела к массе, что апатозавры вырастали до 25 тонн за 15 лет, с пиком роста в 5000 килограммов за один год. Альтернативный метод, использующий длину конечностей и массу тела, показал, что апатозавр рос на 520 килограммов в год и достигал своей полной массы прежде, чем ему исполнялось 70 лет[63]. Однако, эти исследования были признаны ненадёжными, поскольку сами расчёты в статье приведены не были, а также из-за того, что более старые линии роста стирались в результате ремоделирования кости[64]. Одним из первых идентифицированных факторов роста апатозавра было количество крестцовых позвонков, которое увеличивалось до пяти единиц к моменту взросления животного. Впервые этот фактор был отмечен в 1903 году, а затем в 1936 году[2].
Гистология длинных костей позволила исследователям оценить возраст, которого достигала конкретная особь. Ева Грибелер с соавторами в 2013 году изучила гистологические данные длинных костей и пришла к выводу, что образец апатозавра SMA 0014 при жизни весил 20 206 килограммов, достиг половой зрелости в 21 год и умер в возрасте 28 лет. Та же модель роста показала, что другой образец апатозавра, BYU 601–17328, весил 18 178 килограммов, достиг половой зрелости в 19 лет и умер в возрасте 31 года[64].
Детёныши
По сравнению с другими завроподами, известно большое количество ископаемого материала, принадлежавшего неполовозрелым особям апатозавра. Множество скелетов, размещённых в Оклахомском музее естественной истории, принадлежали детёнышам апатозавра неустановленного вида. Этот материал включает частичные плечевые и тазовые пояса, некоторые позвонки и кости конечностей. Ювенильный материал музея относится как минимум к двум разным возрастным группам и, судя по перекрывающимся костям, вероятно, происходит от более чем трёх особей. Образцы демонстрируют особенности, которые отличают апатозавров от их родственников, и, таким образом, вероятно, принадлежат к этому роду[16][65]. У молодых завропод шея и хвост были пропорционально короче, а также присутствовало более выраженное несоответствие между передними и задними конечностями, чем у взрослых представителей группы[66].
Хвост
В статье, опубликованной в 1997 году под авторством изобретателя Натана Мирволда и палеонтолога Филипа Карри, сообщалось об исследовании механики хвостов апатозавров. Мирволд провёл компьютерную симуляцию хвоста, который у диплодоцид, таких как апатозавр, представлял собой очень длинную сужающуюся структуру, напоминающую кнут. Это компьютерное моделирование показало, что диплодоциды были способны издавать щёлкающий звук, похожий на звук хлыста, мощностью более 200 децибел, что сравнимо с громкостью выстрела из пушки[67].
На хвосте одного из образцов апатозавра обнаружена патология, вызванная дефектом роста. Два хвостовых позвонка плавно срастаются по всей суставной поверхности кости, включая дуги нервных отростков. Этот дефект мог быть вызван отсутствием или ингибированием вещества, образующего межпозвоночные диски или суставы[68]. Было высказано предположение, что кнуты могли использоваться в бою и защите, однако хвосты диплодоцид были довольно лёгкими и узкими по сравнению с хвостами шунозавров и мамэньсизаврид, и, таким образом, ранение хвостом другого животного серьёзно повредило бы сам хвост[67].
Палеоэкология

Формация Моррисон представляет собой последовательность мелководных морских и аллювиальных отложений, которые, согласно радиометрическому датированию, датируются периодом от 156,3 миллионов лет в нижней части[69] до 146,8 млн лет в верхней части[70], что позволяет отнести её к позднему оксфордскому, кимериджскому и нижнетитонскому ярусам верхней юры. Считается, что это образование происходит из полузасушливой среды с чётко выраженными влажными и сухими сезонами. Бассейн реки Моррисон, где жили динозавры, простирался от Нью-Мексико до Альберты и Саскачевана; он образовался, когда Передовой хребет Скалистых гор только начал подниматься на западе. Отложения из обращённых на восток водосборных бассейнов переносились ручьями и реками и откладывались в заболоченных низинах, озёрах, руслах рек и поймах[71].

В формации Моррисон апатозавр был вторым по распространённости завроподом после камаразавра[41]. Возможно, апатозавры вели более уединённый образ жизни, чем прочие динозавры формации[72]. Ископаемые остатки этого рода были обнаружены только в отложениях верхних уровней формации. Вид Apatosaurus ajax известен исключительно из верхнего слоя стратиграфического горизонта Браши-Бэйсин, который датируют примерно 152—151 миллионами лет назад. Остатки A. louisae встречаются редко, они известны только на одном участке в верхней части Браши-Бэйсин; их датируют верхним киммериджским ярусом, около 151 млн лет назад. Другие остатки апатозавра известны из пород того же возраста или немного моложе, но они не были идентифицированы как какие-либо конкретные виды[73] и, таким образом, могут принадлежать бронтозаврам[18].
Формация Моррисон фиксирует время, когда в местной окружающей среде доминировали гигантские динозавры-завроподы[41]. Динозавры, чьи фоссилии найдены здесь, включают теропод аллозавра, цератозавра, орнитолеста, Saurophaganax и торвозавра; завропод бронтозавра, брахиозавра, камаразавра и диплодока; птицетазовых камптозавра, дриозавра и стегозавра[74]. По большей части, остатки апатозавров находят в тех же отложениях, что и остатки аллозавров, камаразавров, диплодоков и стегозавров[72]. Окаменелые остатки аллозавров составляют 70—75 % остатков теропод; этот хищник находился на верхнем трофическом уровне пищевой цепи Моррисона[75]. Многие динозавры формации Моррисон принадлежат к тем же родам, что и динозавры, встречающиеся в породах португальской формации Лоринья — в основном это аллозавры, цератозавры и торвозавры — или имеют близких аналогов — брахиозавров и Lusotitan, камптозавров и Draconyx, а также апатозавров и Dinheirosaurus[22]. Другими известными позвоночными, делившими с апатозаврами среду обитания, являются лучепёрые рыбы, лягушки, саламандры, черепахи, сфенодонты, ящерицы, наземные и водные крокодиломорфы, а также несколько видов птерозавров. Для этой местности также характерны раковины двустворчатых моллюсков и водных улиток. О флоре того периода свидетельствуют окаменелости зелёных водорослей, грибов, мхов, хвощей, саговников, гинкго и нескольких семейств хвойных деревьев. Растительность варьировалась от лесов из древовидных папоротников с подлеском из обычных папоротников (галерейные леса), до папоротниковых саванн с редкими деревьями, такими как хвойное дерево, похожее на араукарию — Brachyphyllum[76].
Примечания
- Апатозавры / Алифанов В. Р. // Анкилоз — Банка. — М. : Большая российская энциклопедия, 2005. — С. 104. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 2). — ISBN 5-85270-330-3.
- Gilmore C. W. Osteology of Apatosaurus, with special references to specimens in the Carnegie Museum (англ.) // Memoirs of the Carnegie Museum. — 1936. — Vol. 11, no. 4. — P. 1–136.
- Seebacher F. A new method to calculate allometric length-mass relationships of dinosaurs (англ.) // Journal of Vertebrate Paleontology. — 2001. — Vol. 22, no. 1. — P. 51—60. — doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Mazzetta G. V., Christiansen P., Farina R. A. Giants and bizarres: body size of some southern South American Cretaceous dinosaurs (англ.) // Historical Biology. — 2004. — Vol. 16, no. 2—4. — P. 71—83. — doi:10.1080/08912960410001715132.
- Henderson D. M. Burly Gaits: Centers of mass, stability, and the trackways of sauropod dinosaurs (англ.) // Journal of Vertebrate Paleontology. — 2006. — Vol. 26, no. 4. — P. 907—921. — doi:10.1671/0272-4634(2006)26[907:BGCOMS]2.0.CO;2.
- Bates K. T., Falkingham P. L., Macaulay S., Brassey C., Maidment S. C. R. Downsizing a giant: re-evaluating Dreadnoughtus body mass (англ.) // Biology Letters. — 2015. — Vol. 11, no. 6. — P. 20150215. — doi:10.1098/rsbl.2015.0215.
- Wedel M. A giant, skeletally immature individual of Apatosaurus from the Morrison Formation of Oklahoma (англ.) // 61st Symposium on Vertebrate Palaeontology and Comparative Anatomy - Programme and Abstracts. — 2013. — P. 40.
- Fastovsky D. E., Weishampel D. B. Dinosaurs: A Concise Natural History (англ.). — Cambridge University Press, 2009. — P. 165–200. — ISBN 978-0-521-88996-4.
- Whitlock J. A. Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses (англ.) // PLoS ONE. — 2011. — Vol. 6, no. 4. — P. e18304. — doi:10.1371/journal.pone.0018304.
- Balanoff A. M., Bever G. S., Ikejiri T. The Braincase of Apatosaurus (Dinosauria: Sauropoda) Based on Computed Tomography of a New Specimen with Comments on Variation and Evolution in Sauropod Neuroanatomy (англ.) // American Museum Novitates. — 2010. — Vol. 3677. — P. 1–32. — doi:10.1206/591.1.
- Taylor M. P., Wedel M. J., Naish D., Engh B. Were the necks of Apatosaurus and Brontosaurus adapted for combat? (англ.) // PeerJ : PrePrints. — 2015. — Vol. 3. — P. e1663. — doi:10.7287/peerj.preprints.1347v1.
- Riggs E. S. Structure and Relationships of Opisthocoelian Dinosaurs. Part I, Apatosaurus Marsh (англ.) // Publications of the Field Columbian Museum Geographical Series. — 1903. — Vol. 2, no. 4. — P. 165–196.
- Lovelace D. M., Hartman S. A., Wahl W. R. Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny (англ.) // Arquivos do Museu Nacional. — 2007. — Vol. 65, no. 4. — P. 527—544. — ISSN 0365-4508.
- Upchurch P. Manus claw function in sauropod dinosaurs (англ.) // Gaia. — 1994. — Vol. 10. — P. 161—171. — ISSN 0871-5424.
- Martin A. J. Introduction to the Study of Dinosaurs (Second ed.) (англ.). — Blackwell Publishing, 2006. — P. 560. — ISBN 978-1-4051-3413-2.
- Upchurch P., Tomida Y., Barrett P. M. A new specimen of Apatosaurus ajax (Sauropoda: Diplodocidae) from the Morrison Formation (Upper Jurassic) of Wyoming, USA (англ.) // National Science Museum Monographs. — 2005. — Vol. 26, no. 118. — P. 1–156. — ISSN 1342-9574.
- Marsh O. C. Notice of New Dinosaurian Reptiles from the Jurassic formation (англ.) // American Journal of Science. — 1877. — Vol. 14, no. 84. — P. 514–516. — doi:10.2475/ajs.s3-14.84.514.
- Tschopp E., Mateus O. V., Benson R. B. J. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda) (англ.) // PeerJ. — 2015. — Vol. 3. — P. e857.
- Liddell G. H., Scott R. A Greek-English Lexicon (англ.). — Harper & Brothers, 1882. — 1774 p.
- Holtz T. R. Jr. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages (англ.). — Random House, 2008. — 432 p. — ISBN 978-0-375-82419-7.
- Marsh O. C. Notice of new Jurassic dinosaurs (англ.) // American Journal of Science. — 1879. — Vol. 18, no. 108. — P. 501–505. — doi:10.2475/ajs.s3-18.108.501.
- Mateus O. Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison // Paleontology and Geology of the Upper Jurassic Morrison Formation (англ.) / Foster J. R., Lucas S. G.. — New Mexico Museum of Natural History and Science Bulletin, 2005. — P. 223–231.
- Bakker R. T. The Bite of the Bronto (англ.) // Earth. — 1994. — Vol. 3, no. 6. — P. 26—33.
- Matthew W. D. The mounted skeleton of Brontosaurus (англ.) // The American Museum Journal. — 1905. — Vol. 5, no. 2. — P. 63–70.
- McIntosh, J.S.; Berman, D.S. Description of the Palate and Lower Jaw of the Sauropod Dinosaur Diplodocus (Reptilia: Saurischia) with Remarks on the Nature of the Skull of Apatosaurus (англ.) // Journal of Paleontology. — 1975. — Vol. 49, no. 1. — P. 187—199.
- Miller B. Bully for Camarasaurus (англ.) (30 октября 2014). Дата обращения: 16 июня 2020.
- Keith M. Parsons. The Wrongheaded Dinosaur (англ.). Carnegie Magazine (1997). Дата обращения: 22 июня 2020.
- Crafton D. C. Before Mickey: The Animated Film 1898–1928 (англ.). — MIT Press, 1982. — 1–57 p. — ISBN 978-0-262-03083-0.
- Glut D. F. Dinosaurs: The Encyclopedia (англ.). — Jefferson, N. C.: McFarland, 1997. — P. 150—163. — ISBN 978-0-7864-7222-2.
- Barrett P. M., Storrs G. W., Young M. T., Witmer L. M. A new skull of Apatosaurus and its taxonomic and palaeobiological implications (англ.) // Symposium of Vertebrate Palaeontology & Comparative Anatomy Abstracts of Presentations. — 2011. — P. 5.
- Taylor M. P. Sauropod dinosaur research: a historical review (англ.) // Geological Society, London, Special Publications. — 2010. — Vol. 343, no. 1. — P. 361–386. — doi:10.1144/SP343.22.
- Brinkman P. Bully for Apatosaurus (англ.) // Endeavour. — 2006. — Vol. 30, no. 4. — P. 126–130. — doi:10.1016/j.endeavour.2006.10.004.
- Upchurch P., Barrett P. M., Dodson P. Sauropoda // The Dinosauria / Weishampel D. B., Osmólska H., Dodson P.. — University of California Press, 2004. — С. 259—322. — ISBN 978-0-520-25408-4.
- McIntosh J. S. Remarks on the North American sauropod Apatosaurus Marsh (англ.) // Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota Short Papers. — 1995. — P. 119–123.
- Bakker R. T. Dinosaur mid-life crisis: the Jurassic-Cretaceous transition in Wyoming and Colorado // Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin (англ.) / Lucas S. G., Kirkland J. I., Estep J. W.. — Albuquerque, 1998. — Vol. 14. — P. 67–77.
- Michael D'Emic. Not So Fast, Brontosaurus. Time.com. Дата обращения: 28 июня 2020.
- Donald Protero. Is "Brontosaurus" Back? Not So Fast!. Skeptic.com. Дата обращения: 28 июня 2020.
- Sauropodomorpha // Dinosaur Systematics, Approaches and Perspectives (англ.) / Carpenter K., Currie P. J.. — Cambridge: Cambridge University Press, 1990. — 318 p. — ISBN 978-0-521-36672-4.
- Carpenter K., McIntosh J. Upper Jurassic sauropod babies from the Morrison Formation // Dinosaur Eggs and Babies (англ.) / Carpenter K., Hirsch K. F., Horner J. R.. — Cambridge: Cambridge University Press. — P. 265—278. — ISBN 978-0-521-56723-7.
- Marsh O. C. Principal Characters of American Jurassic Dinosaurs (англ.) // American Journal of Science. — 1878. — Vol. 16, no. 95. — P. 412–414.
- Foster J. Jurassic West: The Dinosaurs of the Morrison Formation and Their World (англ.). — Indiana University Press, 2007. — P. 273–329. — ISBN 978-0-253-34870-8.
- Taylor M. P. “Apatosaurus” minimus sacrum/ilia, right lateral view. Sauropod Vertebra Picture of the Week. Дата обращения: 29 июня 2020.
- Taylor M. P., Wedel M. J. Re-evaluating "Apatosaurus" minimus, a bizarre Morrison Formation sauropod with diplodocoid and macronarian features (англ.) // SVPCA 2012 Programme and Abstracts. — 2012. — Vol. 23.
- Taylor M. P., Naish D. The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda) (англ.) // PaleoBios. — 2005. — Vol. 25, no. 2. — P. 1—7.
- Harris J. D. The significance of Suuwassea emiliae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution (англ.) // Journal of Systematic Palaeontology. — 2006. — Vol. 4, no. 2. — P. 185–198. — doi:10.1017/S1477201906001805.
- Whitlock J. A. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda) (англ.) // Zoological Journal of the Linnean Society. — 2011. — Vol. 161, no. 4. — P. 872–915. — doi:10.1111/j.1096-3642.2010.00665.x.
- Pierson D. J. The Physiology of Dinosaurs: Circulatory and Respiratory Function in the Largest Animals Ever to Walk the Earth (англ.) // Respiratory Care. — 2009. — Vol. 54, no. 7. — P. 887—911. — doi:10.4187/002013209793800286.
- Stevens K. A., Parrish J. M. Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs (англ.) // Science. — 1999. — Vol. 284, no. 5415. — P. 798–800. — doi:10.1126%2Fscience.284.5415.798.
- Stevens K. A., Parrish J. M. Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs // Thunder Lizards: The Sauropodomorph Dinosaurs (англ.) / Carpenter K., Tidswell V.. — Indiana University Press, 2005. — P. 212—232. — ISBN 978-0-253-34542-4.
- Sellers W. I., Margetts L., Coria R. A., Manning P. L. March of the Titans: The Locomotor Capabilities of Sauropod Dinosaurs (англ.) // PLOS ONE. — 2012. — Vol. 8, no. 10. — P. e78733. — doi:10.1371/journal.pone.0078733.
- Switek B. Did Wee Little Sauropods Stand Up to Run? (англ.). Smithsonian.com. Smithsonian Magazine (2 ноября 2010). Дата обращения: 23 июля 2020.
- Tracks of a running bipedal baby brontosaur? Baby sauropod footprints discovered in Colorado (англ.). ScienceDaily. The Geological Society of America (1 ноября 2010). Дата обращения: 23 июля 2020.
- Upchurch P. Neck Posture of Sauropod Dinosaurs (англ.) // Science. — 2000. — Vol. 287, no. 5453. — P. 547b. — doi:10.1126/science.287.5453.547b.
- Taylor M. P. Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs (англ.) // PeerJ. — 2014. — Vol. 2. — P. e712. — doi:10.7717/peerj.712.
- Taylor M. P., Wedel M. J., Naish D. Head and neck posture in sauropod dinosaurs inferred from extant animals (англ.) // Acta Palaeontologica Polonica. — 2009. — Vol. 54, no. 2. — P. 213—220. — doi:10.4202/app.2009.0007.
- Cobley M. J., Rayfield E. J., Barrett P. M. Inter-Vertebral Flexibility of the Ostrich Neck: Implications for Estimating Sauropod Neck Flexibility (англ.) // PLoS One. — 2013. — Vol. 8, no. 8. — P. e72187. — doi:10.1371/journal.pone.0072187.
- Tia Ghose. Ouch! Long-Necked Dinosaurs Had Stiff Necks. Livescience.com (15 августа 2013). Дата обращения: 23 июля 2020.
- Paladino F. V., Spotila J. R., Dodson P. A Blueprint for Giants: Modeling the Physiology of Large Dinosaurs // The Complete Dinosaur (англ.) / Farlow J. O., Brett-Surman M. K.. — Indiana University Press, 1997. — P. 491–504. — ISBN 978-0-253-33349-0.
- Wedel M. J. Vertebral Pneumaticity, Air Sacs, and the Physiology of Sauropod Dinosaurs (англ.) // Paleobiology. — 2003. — Vol. 29, no. 2. — P. 243–255. — doi:10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2.
- Spotila J. R., O'Connor M. P., Dodson P. R., Paladino F. V. Hot and cold running dinosaurs. Metabolism, body temperature, and migration (англ.) // Modern Geology. — 1991. — Vol. 16. — P. 203—227.
- Farlow J. A. Speculations About the Diet and Physiology of Herbivorous Dinosaurs (англ.) // Paleobiology. — 1987. — Vol. 13, no. 1. — P. 60–72. — doi:10.1017/S0094837300008587.
- Curry K. A. Ontogenetic histology of Apatosaurus (Dinosauria: Sauropoda): new insights on growth rates and longevity (англ.) // Journal of Vertebrate Paleontology. — 1999. — Vol. 19, no. 4. — P. 654—665. — doi:10.1080/02724634.1999.10011179.
- Lehman T. M., Woodward H. N. Modelling growth rates for sauropod dinosaurs (англ.) // Paleobiology. — 2008. — Vol. 34, no. 2. — P. 264—281. — doi:10.1666/0094-8373(2008)034[0264:MGRFSD]2.0.CO;2.
- Griebeler E.M., Klein N., Sander P. M. Aging, Maturation and Growth of Sauropodomorph Dinosaurs as Deduced from Growth Curves Using Long Bone Histological Data: An Assessment of Methodological Constraints and Solutions (англ.) // PLOS One. — 2013. — Vol. 8, no. 6. — P. e67012. — doi:10.1371/journal.pone.0067012.
- Carpenter K., McIntosh J. S. Dinosaur Eggs and Babies (англ.) / Carpenter K., Hirsch K. F., Horner J. R.. — Cambridge University Press, 1994. — P. 265—274. — ISBN 978-0-521-44342-5.
- Wedel M. Get down, get fuzzy, speculative juvenile Apatosaurus! (англ.) (27 февраля 2013). Дата обращения: 31 октября 2020.
- Myhrvold N. P., Currie P. J. Supersonic sauropods? tail dynamics in the diplodocids (англ.) // Paleobiology. — 1994. — Vol. 23, no. 4. — P. 393—409. — doi:10.1017/S0094837300019801.
- Lovelace D. M. Developmental Failure of Segmentation in a Caudal Vertebra of Apatosaurus (Sauropoda) (англ.) // The Anatomical Record. — 2014. — Vol. 297, no. 7. — P. 1262—1269. — doi:10.1002/ar.22887.
- Trujillo K. C., Chamberlain K. R., Strickland A. Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations (англ.) // Geological Society of America Abstracts with Programs. — 2006. — Vol. 38, no. 6. — P. 7.
- Bilbey S. A. Cleveland-Lloyd Dinosaur Quarry — age, stratigraphy and depositional environments // The Morrison Formation: An Interdisciplinary Study (англ.) / Carpenter K., Chure D., Kirkland J. I.. — Taylor and Francis Group, 1998. — P. 87—120. — ISBN 0026-7775.
- Russell D. A. An Odyssey in Time: Dinosaurs of North America (англ.). — Minocqua, Wisconsin: NorthWord Press, 1989. — P. 64—70. — ISBN 978-1-55971-038-1.
- Dodson P., Behrensmeyer A. K., Bakker R. T., McIntosh J. S. Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation (англ.) // Paleobiology. — 1980. — Vol. 6, no. 2. — P. 208—232. — doi:10.1017/S0094837300025768.
- Turner C. E., Peterson F. Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A. // Vertebrate Paleontology in Utah (англ.) / Gillette D. D.. — Utah Geological Survey Miscellaneous Publication, 1999. — P. 77—114.
- Chure D. J., Litwin R., Hasiotis S. T., Evanoff E., Carpenter K. The fauna and flora of the Morrison Formation: 2006 // Paleontology and Geology of the Upper Jurassic Morrison Formation (англ.) / Foster J. R., Lucas S. G.. — New Mexico Museum of Natural History and Science Bulletin, 2006. — Vol. 36. — P. 233—248.
- Foster J. R. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. (англ.) : New Mexico Museum of Natural History and Science Bulletin. — 2003. — Vol. 23. — P. 29.
- Carpenter K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus // Paleontology and Geology of the Upper Jurassic Morrison Formation (англ.) / Foster J. R., Lucas S. G.. — New Mexico Museum of Natural History and Science Bulletin, 2006. — Vol. 36. — P. 131—138.