Цератозавр
Цератоза́вр[1][2] (лат. Ceratosaurus, от κερας/κερατος [keras/keratos] — «рогатый», и σαυρος [sauros] — «ящерица, рептилия») — монотипический род хищных динозавров подотряда теропод, живших в позднем юрском периоде (киммериджский — титонский века, 155,7—145 млн лет назад). Единственным видом является Ceratosaurus nasicornis[3].
| † Цератозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Инфраотряд: Клада: † Неоцератозавры Семейство: Род: † Цератозавр |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Ceratosaurus Marsh, 1884 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
† Ceratosaurus nasicornis Marsh, 1884
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 155,7—145,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||


Типовой экземпляр, найденный в формации Моррисон в США, назвал и описал в 1884 году известный американский палеонтолог Отниел Чарлз Марш. Впоследствии в формации Моррисон было обнаружено множество окаменелых остатков цератозавров, в том числе подростковых особей. Образцы, отнесённые к этому таксону, также были найдены на территории Танзании и Португалии.
Цератозавр был крупным бипедальным динозавром с мощными задними конечностями и редуцированными передними. Длину скелета самого крупного из найденных экземпляров цератозавра оценивают приблизительно в 7 м. Масса, по разным оценкам, сильно варьирует: от 275 до 980 кг у разных экземпляров.
Отличительной особенностью анатомии цератозавра был рельефный рог на носовых костях черепа, из-за которого ящер получил своё название, а также два роговых выступа над глазами. Больше половины зубов верхней челюсти были очень длинными и прочными; зубы нижней челюсти были меньше по размеру и менее крепкими. Вдоль средней линии спины шкуру цератозавров покрывал ряд остеодерм.
Цератозавры обитали возле рек, озёр и болот, нередко заходя в воду, и охотились на водных животных: рыб, крокодиломорфов и черепах. Однако, они могли разнообразить свой рацион мясом более крупных наземных травоядных, участвуя при этом в конкурентной борьбе с аллозаврами.
История открытия

Первый образец цератозавра, ставший впоследствии типовым экземпляром USNM 4737, нашёл фермер Маршалл Паркер Фелч в 1883—1884 годах[4]. Образец представлял собой почти полный сочленённый скелет с черепом, располагавшийся в исходном анатомическом положении. Окаменелости находились в твёрдом песчанике; череп и позвоночник сильно деформировались во время фоссилизации[5]. Место находки, расположенное на территории Гарден Парка к северу от Каньон-Сити и известное как «Карьер Фелча 1» (Felch Quarry 1), считается одним из самых богатых на окаменелости участков формации Моррисон. Ещё до открытия цератозавра здесь были обнаружены и извлечены многочисленные остатки динозавров и прочих вымерших рептилий, в том числе, почти полный экземпляр аллозавра. Найденный образец отправили в Музей Естественной истории Пибоди в Нью-Хейвене, штат Коннектикут, где его изучил палеонтолог Отниел Чарльз Марш, который и описал динозавра в 1884 году под новым именем Ceratosaurus nasicornis[6][7]. Учитывая превосходную сохранность образца, новый род стал самым известным в то время тероподом, обнаруженным в Америке. Во время подготовки черепа, под наблюдением Марша был изготовлен слепок внутренней части черепной коробки, который позволил учёному сделать вывод, что «мозг был среднего размера, но в сравнении значительно больше, чем мозг травоядных динозавров»[5][7]. Тем не менее, кости черепа были прочно скреплены вместе, так что точность этого заключения не была подтверждена поздними исследованиями. В 1898—1899 годах образец был передан Национальному музею естественной истории в Вашингтоне вместе с другими окаменелостями, описанными Маршем. Упаковка и транспортировка из Нью-Хейвена в Вашингтон нанесли некоторый ущерб образцу цератозавра. Также оказалось, что только небольшая часть ископаемого материала была подготовлена для исследования. Работы по препарированию продолжались с 1911 по 1918 годы[8]. В 1920 году палеонтолог Чарльз Гилмор опубликовал обширное повторное описание этого экземпляра, а также других образцов теропод из Нью-Хейвена, включая почти полный экземпляр аллозавра (упомянутого Гилмором как антродем), извлечённый из того же карьера, что и цератозавр[5].

В статье 1892 года Марш опубликовал первую реконструкцию цератозавра, где указал размеры животного: 6,7 метра в длину и 3,7 метра в высоту[9]. Как отмечал в 1920 году Гилмор, туловище на иллюстрации Марша получилось слишком длинным, поскольку позвоночник включал по меньшей мере 6 лишних грудных позвонков. Ошибка Марша повторилась в нескольких последующих публикациях, включая первую реконструкцию внешнего вида, которую выполнил в 1899 году Фрэнк Бонд под руководством Чарльза Найта и которая не была опубликована до 1920 года. Более точную реконструкцию внешнего вида выполнил в 1901 году художник Дж. М. Глисон, также под руководством Найта. Типовой образец был смонтирован Гилмором в 1910—1911 годах и экспонировался в Национальном музее естественной истории в Вашингтоне. Самые ранние реконструкции показывают цератозавра в вертикальной позе, волочащим хвост по земле[5]. Смонтированный Гилмором скелет, напротив, изображал животное в более продвинутой позиции[10]: на основании кости бедра, найденной под углом к голени, он собрал скелет в виде бегущего животного в более горизонтальной, чем вертикальной позиции, и с хвостом, который не контактировал с землёй. Из-за сильного уплощения костей Гилмор установил образец не как отдельно стоящий скелет, а в виде барельефа в искусственной стене[5].
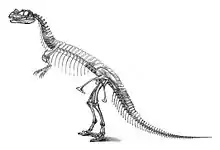
После находки типового образца довольно долго не было сделано ни одного значительного открытия цератозавров. Лишь в начале 1960-х годов команда палеонтологов под руководством Джеймса Мэдсена обнаружила неполный разъединённый скелет с черепом (образец UMNH 5278) в Динозавровом карьере Кливленд-Ллойда в штате Юта. Эта находка представляет собой один из крупнейших обнаруженных скелетов цератозавров[11]. Второй, сочленённый образец, включающий череп (MWC 1), обнаружил Тор Эриксон, сын палеонтолога Ланса Эриксона, в 1976 году возле города Фрута, штат Колорадо[12]. Это достаточно полный экземпляр, у которого отсутствуют только нижняя челюсть, кости предплечий и гастралии. Скелет принадлежал крупной, но не полностью взрослой особи, о чём свидетельствуют несрощенные кости черепа. Оба экземпляра описали Джеймс Мэдсен и Сэмьюэл Пол Уэллс, в монографии 2000 года, где образец из Юты упоминается как Ceratosaurus dentisulcatus, а образец из Колорадо — как Ceratosaurus magnicornis[13]. Видовое название dentisulcatus относится к параллельным канавкам, имеющимся на внутренних сторонах премаксиллярных зубов и первым трём зубам нижней челюсти в этом образце; magnicornis указывает на большой рог на носу[14]. Однако, в последующих публикациях валидность обоих видов была поставлена под сомнение: Бритт и др. в 2000 году утверждали, что типовой экземпляр Ceratosaurus nasicornis на самом деле принадлежал неполовозрелой особи, а два более крупных вида представляют собой взрослых особей одного вида[15]. Оливер Раухут в 2003 году, а также Мэттью Каррано и Скотт Сэмпсон в 2008 году посчитали анатомические различия у описанных Мэдсеном и Уэллсом видов, в качестве онтогенетических или индивидуальных вариаций[16][17].
Ещё один экземпляр (BYUVP 12893) был обнаружен в 1992 году в районе Агат Бэйсин, на юго-востоке от населённого пункта Мур, штат Юта. Этот образец всё ещё не описан. Он считается самым крупным образцом цератозавра из всех найденных и включает в себя полный череп с остатками рога, семь грудных позвонков и прекрасно сохранившиеся сочленённые тазовые кости и крестец[18][19].

В 1999 году палеонтолог Брукс Бритт сообщил об открытии первого скелета цератозавра, принадлежавшего подростковой особи. Обнаруженный в Боун Кэбин Кворри в штате Вайоминг, он на 34 % меньше образца голотипа и состоит из полного черепа и примерно 30 % скелета, включая полный таз[20].
Помимо этих находок, были зарегистрированы фрагментарные остатки цератозавров, обнаруженные в нескольких зонах раскопок формации Моррисон, включая некоторые из основных ископаемых участков формации[21]. Национальный памятник Динозавр, штат Юта, дал изолированную правую предчелюстную кость; из Комо Блафф в Вайоминге сообщалось о находке большого скапулокоракоида. Ещё один экземпляр происходит из Драй Меза Кворри, штат Колорадо, и включает левый скапулокоракоид, а также фрагменты позвонков и костей конечностей. Из Майгэт Мур Кворри, Колорадо, род известен по ископаемым зубам[13].
Находки за пределами Северной Америки
В ходе Германской экспедиции 1909—1913 годов, проведённой Берлинским музеем естествознания, было выявлено наличие остатков разнообразной динозавровой палеофауны в формации Тендагуру, в Германской Восточной Африке (современная Танзания)[22]. Несмотря на то, что эта формация считается самым важным африканским месторождением окаменелостей динозавров[22], крупные тероподы здесь известны только по нескольким фрагментарным образцам[23]. В 1920 году немецкий палеонтолог Вернер Яненш отнёс несколько грудных позвонков из раскопа «TL» к неопределённому виду цератозавра. В 1952 году Яненш назвал новый вид цератозавров — Ceratosaurus roechlingi на основе фрагментарных остатков из раскопа «Mw». Этот экземпляр принадлежал особи, значительно превышающей по размеру голотип цератозавра[23]. Мэдсен и Уэллс в своей монографии 2000 года подтвердили принадлежность этих остатков цератозавру, и отнесли к нему же ископаемые остатки зубов, которые сам Яненш описал под биноменом Labrosaurus (?) stechowi[13]. Многие авторы ставят вопрос о причислении к цератозавру любых ископаемых остатков теропод из Тендагуру, отмечая, что ни один из образцов не показывает признаки, по которым диагностируется этот род[17][24][25][26].
В 1990 году Т. Роу и Ж. Готье описали второй вид цератозавра из формации Тендагуру — Ceratosaurus ingens, который предположительно выделил в 1920 году Яненш на основе 25 очень больших изолированных зубов длиной до 15 см[23][24]. Однако, Яненш фактически отнёс этот вид не к цератозавру, а к мегалозавру; этот биномен может быть просто ошибкой копирования[13][23]. В настоящее время Ceratosaurus ingens считается сомнительным видом в составе клады Tetanurae[27], а Labrosaurus (?) stechowi, возможно, представляет собой род, тесно связанный с цератозаврами[28].
В 2000 и 2006 годах команда палеонтологов под руководством Октавио Матеуша описала находку в формации Лориньян, расположенной в центральной части Португалии, в качестве нового экземпляра цератозавра. Образец состоит из правой бедренной кости, левой большеберцовой и нескольких разрозненных зубов, извлечённых из скальных пород в муниципалитетах Лориньян и Торриш-Ведраш[29][30]. В 2006 году Матеуш и др. отнесли образцы к виду Ceratosaurus dentisulcatus[30]. В своём обзоре 2008 года Каррано и Сэмпсон подтвердили принадлежность окаменелостей к роду цератозавры, но отметили, что причисление образцов к новому виду невозможно[17]. Элисабете Малафайа и её коллеги в 2015 году поставили под вопрос принадлежность португальских образцов к виду C. dentisulcatus и соотнесли их с видом C. nasicornis[31].
Описание
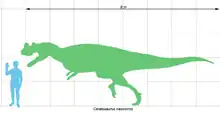
Строение тела цератозавра было типичным для крупного теропода[32]. Будучи бипедальным, он передвигался на мощных задних лапах, в то время как его передние конечности были маленькими. Типовой образец принадлежал особи длиной в 5,3 метра; неясно, было ли это животное полностью выросшим[5][25]. В 1884 году Марш предположил, что типовой экземпляр цератозавра весил примерно в два раза больше, чем аллозавр такой же длины[7]. По более поздним оценкам от других авторов, масса типового образца цератозавра могли достигать 418, 524 или 670 килограммов[33]. Два скелета, которые Джеймс Мэдсен и Сэмьюэл Уэллс отнесли к видам C. magnicornis и C. dentisulcatus, были значительно больше типового экземпляра[13][34]. Длину более крупного из них, C. dentisuclatus, Мэдсен неофициально оценил в 8,8 метра[12]. Однако, в 2016 году американский учёный и писатель Грегори С. Пол оценил длину этого образца в 7 метров[35]. В 1988 году он оценивал массу C. dentisulcatus в 980 кг[36]. В 2007 году Джон Фостер дал значительно более низкую оценку массы: 275 кг для C. magnicornis и 452 кг для C. dentisulcatus[37].
Точное количество позвонков неизвестно из-за пропусков в позвоночнике типового образца. Крестец состоял из 6 слитых позвонков. По меньшей мере, 20 позвонков образовывали шейный и грудной отделы позвоночника и не менее 50 — хвостовой. Хвост составлял около половины длины тела; Гилмор оценил его длину у типового экземпляра в 2,84 метра[38]. Как и у других теропод, хвост уравновешивал тело и содержал массивную хвостобедренную мышцу, musculus caudofemoralis, которая отвечала за толчок при движении вперёд, отводя бедро животного назад при своём сокращении. Хвост цератозавра характерен сравнительно высокими поперечными отростками позвонков и удлинёнными шевронами[25].

Типовой образец был найден с сочленёнными костями левой передней конечности, включающей неполную кисть. Несмотря на то, что кости разделили при препарировании, был сделан слепок окаменелости, чтобы описать и задокументировать первоначальное местоположение костей. Кости запястий не были найдены ни в типовом образце, ни в других экземплярах, что позволило учёным высказать предположение, что цератозавры утратили эту часть своего скелета. В статье 2016 года Мэттью Каррано и Джона Шуанье проанализировали слепок и заключили, что на присутствие запястья указывает наличие промежутка между костями предплечья и пястья, а также поверхностная структура этого промежутка, которая отличается от таковой в окружающей породе[39].
Кисть сохраняла четыре пальца, и последний, четвёртый палец был редуцирован, по размеру напоминая таковой у других базальных теропод. I и IV пястные кости были уменьшены, а II кость была немного длиннее III. Пясть и особенно первые фаланги были пропорционально очень короткими, в отличие от таковых у большинства базальных теропод. В типовом экземпляре сохранились только первые фаланги второго, третьего и четвёртого пальцев соответственно; общее же количество фаланг и когтей неизвестно. Анатомия I пястной кости указывает на то, что изначально на этом пальце фаланги присутствовали. Ступня состояла из трёх опорных пальцев (II—IV). Первый палец, который у теропод обычно редуцирован и не касается земли, в типовом образце не сохранился[40]. В первоначальном описании 1884 года Марш предположил, что этот палец у цератозавров был полностью утерян[7], но Гилмор в своей монографии 1920 года отметил область прикрепления на II плюсневой кости, демонстрирующую наличие этого пальца[41].
Остеодермы
Уникальной среди других теропод особенностью является наличие у цератозавра небольших удлинённых остеодерм неправильной формы, расположенных вдоль средней линии спины. Такие остеодермы были обнаружены над поперечными отростками 4 и 5 шейных позвонков, а также над 4—10 хвостовыми позвонками. Вероятно, они образовывали сплошной ряд, который тянулся от основания черепа до большей части хвоста. Согласно Гилмору, их положение в каменной матрице указывает на точное расположение у живого животного. Остеодермы над хвостом были найдены отделёнными от поперечных отростков на 25—38 мм, возможно, с учётом присутствия между ними кожи и мышц, тогда как шейные остеодермы были намного ближе к позвоночнику. Найденная костная пластинка размером 58 на 70 мм указывает на присутствие дополнительных остеодерм, расположенных в других местах, кроме средней линии спины. Однако, точное их местоположение неизвестно[42]. Образец C. dentisulcatus был также обнаружен с рядом остеодерм, форма которых описана как «аморфная». Остеодермы были обнаружены в пределах 5 метров от скелета и не были напрямую связаны ни с одним из позвонков, поэтому их расположение на теле не может быть выведено из этого экземпляра[43].
Череп

Череп был очень большим по отношению к остальному скелету[9]. В типовом образце его длина составляет 625 мм[33], в то время, как длина достаточно полного черепа C. magnicornis оценивается в 600 мм от кончика морды до затылочного мыщелка, при общей ширине в 160 мм и ширине носа в 60 мм[44]. Задняя часть черепа состоит из арок, на удивление тонких для такой большой структуры; неизвестен ни один другой теропод, чей череп был бы построен настолько же непрочно. Тем не менее, челюсти цератозавра были крепкими и высокими и поддерживали пропорционально большие зубы[45].
Рог

Самой отличительной частью головы был рельефный рог, расположенный на средней линии черепа за ноздрями и образованный слитыми окостеневшими выступами левой и правой носовых костей[5]. По окаменелым остаткам известно только костное ядро рога — у живого животного это ядро поддерживало кератиновую оболочку. В типовом образце гладкая часть у основания рога имеет наибольшую поперечную ширину 12 мм, которая резко увеличивается до 20 мм в том месте, где начинается шероховатость сосудистых канавок. Рог имеет наибольшую длину у основания в 130 мм и максимальную высоту в 70 мм над носовыми костями[46]. Сразу за рогом носовые кости образуют овальную бороздку; эти две анатомические особенности черепа — рог и бороздка — служат признаками, отличающими цератозавра от родственных таксонов[18]. В дополнение к большому рогу на носу, цератозавр обладал меньшими, полукруглыми, роговидными гребнями перед каждым глазом, напоминающими таковые у аллозавра; эти гребни были образованы парными слёзными костями[37]. У неполовозрелых особей все три рога были меньше, и две половины ядра носового рога ещё не были окостеневшими[20].
Зубы
Верхнечелюстные кости типового экземпляра содержали по 15 зубов в форме лезвия с каждой стороны. Первые 8 зубов были очень длинными и крепкими, но, начиная с 9, зубы постепенно уменьшались в размерах. Что типично для теропод, зубы были оснащены пилообразной насечкой; типовой образец содержал около 10 зубцов на 5 мм[47]. Образец C. magnicornis содержит только 11—12 зубов, а образец C. dentisulcatus — в каждой половине верхней челюсти, которые у последнего более массивные и более изогнутые[48]. Во всех образцах зубные коронки верхних челюстей были исключительно длинными, длиной до 9,3 см у образца C. dentisulcatus, что равно минимальной высоте нижней челюсти. В типовом образце длина верхних зубных коронок (7 см) даже превосходит минимальную высоту нижней челюсти (6,3 см) — среди других теропод эта особенность известна только у возможного родственного цератозавру рода Genyodectes[49]. Парная предчелюстная кость, образующая кончик рыла, содержала всего три зуба на каждой половине, меньше, чем у большинства других теропод[25]. Каждая половина нижнечелюстной кости содержала 15 зубов, которые плохо сохранились в типовом экземпляре. Оба образца C. magnicornis и C. dentisulcatus показывают только 11 зубов на каждой половинке нижней челюсти, которые, как показывает последний экземпляр, были намного более прямыми и менее прочными, чем в верхней челюсти[13].
Систематика

В первоначальном описании цератозавра Отниел Марш отметил ряд характеристик, неизвестных на то время у теропод[50]. Двумя из этих особенностей являются слитые кости таза и окостеневшая плюсна, которые, однако, характерны для современных птиц. По словам Марша, это наглядно демонстрирует тесную связь между птицами и динозаврами[7]. Чтобы отделить род от аллозавра, мегалозавра и «целурозавра», Марш сделал цератозавра единственным представителем нового инфраотряда Ceratosauria. За целое столетие, прошедшее с тех пор, как Марш основал инфраотряд, лишь немногие его представители были описаны по очень скудным и фрагментарным остаткам. Группа Ceratosauria широко не использовалась, а самого цератозавра иногда считали примитивным карнозавром. Строгий консенсус был достигнут при создании кладистического анализа в 1980-х годах. Американский палеонтолог Жак Готье в 1986 году заявил о тесной связи надсемейства Coelophysoidea с родом цератозавры, хотя в большинстве современных анализов эта группа находится вне инфраотряда Ceratosauria. В 1985 году обнаруженный незадолго до этого южноамериканский род абелизавр признан тесно связанным с цератозаврами. С тех пор было описано много таксонов, находящихся с цератозаврами в родственной связи[50].
Инфраотряд Ceratosauria отделился от эволюционной линии, ведущей к современным птицам, довольно рано, а потому считается базальным в пределах клады тероподы[51]. В инфраотряд включают группу производных таксонов, таких, как Carnotaurus и Noasaurus, помещённых внутрь группы Abelisauroidea, а также несколько базальных представителей: Elaphrosaurus, Deltadromeus и самого цератозавра. Позиция рода цератозавры внутри базального инфраотряда Ceratosauria находится в стадии обсуждения. При некоторых филогенетических анализах цератозавр размещается в качестве терминального таксона (то есть располагается на конце ветви в кладограмме) среди базальных представителей семейства, и является сестринским таксоном для Abelisauroidea[17][52]. Впрочем, Оливер Раухут в 2004 году предложил считать цератозавра сестринским таксоном Genyodectes, поскольку для обоих родов характерны исключительно длинные зубы в верхней челюсти. Раухут сгруппировал цератозавра и Genyodectes внутри семейства Ceratosauridae[49], и такой систематики придерживались авторы последующих работ[53][54]. Однако, в 2015 году, группа аргентинских исследователей под руководством Федерико Джанекини вернула род цератозавры в инфраотряд Ceratosauria[55].
Найденный в Англии череп из среднеюрских отложений, по-видимому, несёт рог на носу, похожий на рог цератозавра. В 1926 году немецкий палеонтолог Фридрих фон Хюне описал этот череп и назвал новый род Proceratosaurus (что означает «до цератозавра»), посчитав его предком позднеюрского цератозавра[56]. Сегодня Proceratosaurus считается базальным представителем надсемейства Tyrannosauroidea, более обособленной группы тероподовых динозавров[57]; следовательно, носовой рог эволюционировал независимо у обоих таксонов[17]. В 2010 году Раухут и его коллеги отнесли Proceratosaurus к собственному семейству Proceratosauridae. Помимо этого, авторы отметили, что носовой рог сохранился не полностью, что могло указывать на наличие у динозавра более обширного гребня, что уже наблюдалось у прочих процератозаврид, таких, как Guanlong[57].
Палеобиология
Экология и питание

В Северной Америке все окаменелые остатки цератозавров были обнаружены в формации Моррисон, расположенной в западной части Соединённых Штатов Америки. Здесь окаменелости часто встречаются в сочетании с остатками других крупных теропод, включая мегалозаврида[2] торвозавра и аллозаврида аллозавра. В местности Гарден Парк в штате Колорадо также найдены остатки цератозавра и образцы, приписываемые аллозавру. Локация Драй Меза Кворри в Колорадо, а также Кливленд-Ллойд Кворри и Национальный памятник Динозавр в штате Юта изобилуют остатками по крайней мере трёх крупных теропод: цератозавра, аллозавра и Torvosaurus[58][59]. Аналогично, Комо-Блафф и соседние локации в штате Вайоминг содержат остатки цератозавров, аллозавров и по крайней мере одного крупного мегалазаврида[60].
Некоторые исследователи предпринимали попытки объяснить, как эти живущие в одно время виды могли снижать межвидовую конкуренцию. В 1998 году Дональд Хендерсон проанализировал цератозавра и две морфы аллозавра — морфу с укороченной мордой, высоким и широким черепом и короткими загнутыми назад зубами, и морфу, характеризующуюся более длинной мордой, низким черепом и длинным, прямыми зубами. В экологии общеизвестным является факт: чем большим будет сходство между симпатричными видами в морфологии, физиологии и поведении, тем интенсивнее будет конкуренция[61]. Хендерсон пришёл к выводу, что морфа аллозавров с короткими черепами занимала иную экологическую нишу, чем морфа с длинными черепами и цератозавры. Более короткий череп этой морфы имел бы меньшие изгибающие моменты при кусании и повышенную силу укуса, который по динамике был похож на таковой у кошек. Напротив, укус цератозавров и длинноголовых аллозавров сравним по динамике с укусом собак: более длинные зубы использовались как клыки для нанесения быстрых режущих укусов, при этом сила укуса рассредотачивалась на меньшей площади из-за узкой формы черепа. С другой стороны, Хендерсон обнаружил большое сходство между цератозавром и морфой аллозавров с длинным черепом и сделал вывод, что обе формы участвовали в прямой конкуренции друг с другом. Таким образом, популяция цератозавров, возможно, была вытеснена из ареала, в котором доминировали длинноголовые аллозавры. Действительно, остатки цератозавров встречаются в Кливленд-Ллойд Кворри реже остатков аллозавра с длинным черепом, в то время как в Драй Меза Кворри образцы цератозавров находят так же часто, как и окаменелости короткоголовой морфы аллозавров. Кроме того, цератозавры могли избегать конкуренции, предпочитая различные части добычи; эволюция их чрезвычайно вытянутых зубов могла представлять собой прямой результат конкуренции с аллозаврами. В таком случае, при поедании туши оба вида могли предпочитать различные её части. Удлинённые зубы также могли служить для распознавания представителей своего вида или выполнять другие социальные функции. Кроме того, Хендерсон утверждает, что большие размеры этих теропод (6—8 м) сами по себе приводили к снижению конкуренции, поскольку с увеличением размера хищника увеличивается и размер массы жертвы[58].

В исследовании 2004 года Роберт Бэккер и Грей Бир предположили, что цератозавр в основном специализировался на водной добыче, такой, как двоякодышащие рыбы, крокодиломорфы и черепахи. Как показывает статистический анализ утерянных животными зубов из 50 различных локаций в Комо-Блафф и вокруг него, зубы цератозавров и мегалозаврид наиболее распространены вокруг водных источников, таких, как поймы рек, берега озёр и болота. Однако, остатки цератозавров также иногда находят и в сухопутных местах обитания. С другой стороны, окаменелости аллозавров одинаково распространены как в водных, так и в сухопутных ареалах. Исходя из этого, Бэккер и Бир пришли к выводу, что цератозавры и мегалозавриды охотились возле водных объектов, нередко залезая в воду, при этом цератозавры могли дополнительно разнообразить свой рацион мясом крупных динозавров, обитающих вдалеке от воды. Исследователи также отметили длинные, низкие и гибкие тела цератозавров и мегалозаврид. По сравнению с другими тероподами формации Моррисон, цератозавры обладали более высокими поперечными отростками передних хвостовых позвонков, которые располагались строго вертикально, а не под наклоном к спине. Вместе с длинными шевронами на нижней стороне хвоста эти анатомические особенности указывают на наличие уплощённого с боков хвоста, напоминающего крокодилий, который, возможно, был нужен для плавания. Напротив, тела аллозваврид были более короткими, более высокими и более крепкими, с длинными ногами. Они были приспособлены для быстрого бега на открытой местности и для охоты на крупных травоядных динозавров, таких как зауроподы и стегозавры, но, как предположили Бэккер и Бир, временами переключались на ловлю водной или околоводной добычи, когда крупные травоядные отсутствовали[60].
Крупные тероподы иногда и сами становились пищей для других плотоядных динозавров. Об этом свидетельствует найденная лобковая кость аллозавра со следами зубов другого теропода, вероятно, другого аллозавра, цератозавра или Torvosaurus. Расположение кости в теле (вдоль нижнего края туловища, частично закрытая ногами), а также тот факт, что лобковая кость является одной из самых тяжёлых и малосъедобных в скелете теропод, указывает на то, что аллозавр был уже некоторое время мёртв до момента поедания[62].
Функции носового рога и передних конечностей
_-_AMNH_-_DSC06292.JPG.webp)
В своей монографии 1884 года Отниел Марш назвал рог цератозавра «самым мощным оружием» для наступательных и оборонительных целей, и Гилмор (1920) согласился с этим определением[63][46]. Однако, в настоящее время их интерпретация считается маловероятной[12]. Британский палеонтолог Дэвид Норман в 1985 году предположил, что рог, вероятно, не был предназначен для защиты от других хищников, а вместо этого мог использоваться во внутривидовых сражениях между мужскими особями цератозавров за право размножения[64]. Грегори С. Пол поддержал это предположение и изобразил двух цератозавров, участвующих в несмертельном поединке[65]. Т. Роу и Ж. Готье (1990) пошли дальше и предположили, что рог выполнял демонстрационную функцию и не играл никакой роли в физическом противостоянии[24]. В этом случае рог, вероятно, мог быть ярко раскрашен[37].
Сильно укороченные пястные кости и фаланги цератозавра наводят на мысль о сохранении хватательной функции кисти у базальных теропод. Среди представителей инфраотряда цератозавров ещё более экстремальное уменьшение кисти наблюдается у семейства абелизаврид, где передние конечности превратились в рудименты[66]. В статье 2016 года Мэттью Каррано и Джона Шуанье подчеркнули большое морфологическое сходство кисти цератозавра с кистью других базальных теропод и предположили, что она всё ещё выполняла хватательную функцию, несмотря на сильную редукцию. Хотя в образцах сохранились только первые фаланги пальцев, вторые фаланги сохраняли подвижность — на это указывают хорошо развитые суставные поверхности и пальцы цератозавров, вероятно, сохраняли аналогичную свободу движения, как у прочих базальных теропод. И, как у других теропод, кроме абелизаврид, первый палец слегка загибался вовнутрь[39].
Слитая плюсна и палеопатология
В типовом экземпляре Ceratosaurus nasicornis, USMN 4735, левые кости плюсны — со второй по четвёртую — были слиты вместе[67]. В 1884 году Марш посвятил короткую статью этой особенности, которая в то время была неизвестна у динозавров. Учёный отметил тесное сходство этой анатомической особенности с таковой у современных птиц[7]. Наличие этой черты у цератозавра было поставлено под сомнение в 1890 году, когда Георг Баур предположил, что слияние плюсневых костей в образце было результатом сросшегося перелома. Тем не менее, примеры слитых костей плюсны у динозавров, которые не относятся к патологическому происхождению, были позже описаны, включая таксоны, более базальные, чем цератозавр[67]. Генри Осборн в 1920 году заключил, что аномальный рост костей не проявлен и, хоть окостенение и необычно, но, вероятно, оно не является патологией[41]. Р. Раткевич в 1976 году выдвинул предположение, что слияние плюсны ограничило бы у динозавра способность к движению, но это опроверг Г. Пол в 1988 году, который отметил, что аналогичная особенность наблюдается у многих быстро движущихся современных животных, включая наземных птиц и копытных[36]. Анализ Даррена Танке и Брюса Ротшильда, опубликованный в 1999 году, предполагает, что слияние плюсневых костей всё-таки было патологическим, подтвердив вывод, сделанный Бауром[67].
Другие сообщения о патологиях в окаменелостях цератозавров включают перелом в отнесённой к таксону кости стопы[68], а также сломанный зуб неизвестного вида цератозавров, который показывает признаки дальнейшего износа, полученные после того, как он сломался[67].
Палеоэкология и палеобиогеография

Формация Моррисон, где были найдены все североамериканские образцы цератозавра, представляет из себя последовательность морских и речных аллювиальных отложений, возраст которых, согласно радиометрическому датированию, колеблется от 156,3 млн лет в нижней части[69] до 146,8 млн лет назад в верхней[70], что соответствует позднему оксфордскому, киммериджскому и раннему титонскому векам юрского периода. Считается, что местность, которая теперь является формацией Моррисон, прежде представляла из себя полузасушливую среду с явно выраженными сухими и влажными сезонами. Бассейн Моррисон простирался от современных Нью-Мексико до Альберты и Саскачевана и сформировался, когда предшественник Передового хребта Скалистых гор начал двигаться на запад. Отложения с восточного края водосбора переносились речными потоками и осаждались в болотистых низинах, озёрах, речных руслах и поймах[71]. Формация Моррисон по возрасту схожа с формациями Лориньян в Поргугалии и Тендагуру в Танзании[72].

Формация Моррисон сохранила среду того времени, когда на Земле доминировали гигантские зауроподы[21]. Здесь найдены ископаемые остатки таких зауропод, как апатозавр, камаразавр, брахиозавр и диплодок. Помимо остатков зауропод, обнаружены окаменелости, принадлежавшие тероподам орнитолесту, аллозавру, Koparion, Stokesosaurus и Torvosaurus. Из птицетазовых здесь обнаружены остатки Camptosaurus, Dryosaurus, Othnielia, Gargoyleosaurus и стегозавра[73]. Кости диплодоков обычно находятся на тех же участках, что и апатозавр, камаразавр, аллозавр и стегозавр[74]. Аллозавр, остатки которого составляют от 70 до 75 % всех окаменелостей теропод, находился на вершине пищевой цепи, будучи сверххищником[75]. Многие динозавры формации Моррисон представлены теми же таксонами, что и в португальской формации Лориньян (аллозавр, цератозавр, стегозавр, Torvosaurus) или имеют близкие аналоги (брахиозавр и Lusotitan, Camptosaurus и Draconyx)[72]. Другими позвоночными, разделявшими с динозаврами среду обитания, были лучепёрые рыбы, лягушки, саламандры, черепахи (Dorsetochelys), сфенодонты, ящерицы, наземные и водные крокодиломорфы, такие, как Hoplosuchus, а также некоторые птерозавры, например, Harpactognathus и Mesadactylus. В отложениях формации широко распространены раковины двустворчатых моллюсков и водных улиток. Флора этого периода показана наличием окаменелых зелёных водорослей, грибов, мхов, хвощей, цикадовых, гинкго и нескольких семейств хвойных деревьев. Растительность варьировалась от галерейных лесов древовидных папоротников, растущих по берегам рек, до заросшими папоротниками саванн со случайными одиночными деревьями, такими, как похожий на араукарию хвойный Brachyphyllum[76].
Найденный неполный экземпляр цератозавра указывает на присутствие этого таксона в горизонте Порто-Ново в формации Лориньян, что свидетельствует о португальской тероподовой фауне, сходной с североамериканской фауной формации Моррисон. Помимо цератозавра, исследователи отметили присутствие в португальских породах остатков аллозавра и Torvosaurus, которые известны и в формации Моррисон, в то время, как Lourinhanosaurus найден только в Португалии. Травоядные динозавры из горизонта Порто-Ново включают в себя, среди прочих, зауропод Dinheirosaurus и Zby, а также стегозавра Miragaia[29][30][77]. В позднеюрскую эпоху Европу от североамериканского континента отделял ещё узкий Атлантический океан, и Португалия, как часть Пиренейского полуострова, была отделена от других частей Европы. По словам Октавио Матеаша и его коллег, сходство между тероподовой фауной Португалии и Северной Америки предполагает наличие сухопутного моста, допускающего обмен фауной[29][30]. Однако Элисабете Малафайа и её команда обсуждают более сложный сценарий, поскольку другие группы пресмыкающихся, такие, как зауроподы, черепахи и крокодилы, явно демонстрируют видовое различие в Португалии и Северной Америке. Таким образом, начальное разделение этих фаун могло привести к обмену в некоторому аллопатрическому видообразованию в других группах[31].
В популярной культуре
Благодаря запоминающемуся рогу цератозавр является одним из самых известных динозавров и регулярно появляется в популярной литературе[12]. Он также появляется в нескольких фильмах, самым первым из которых является немое кино Brute Force 1914 года режиссёра Дэвида Гриффита, где была показана полноразмерная модель цератозавра, который нападал на пещерных людей[78].
Примечания
- Гиляров, 1986, с. 701.
- Нэйш, Барретт, 2019, pp. 47.
- Ceratosaurus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 12 февраля 2018)
- Brinkman, 2010.
- Gilmore, 1920.
- Gilmore, 1920, pp. 7, 114.
- Marsh, 1884.
- Gilmore, 1920, pp. 2, 14.
- Marsh, 1892.
- Paul, 1988, pp. 276.
- Madsen, Welles, 2000, pp. 21.
- Glut, 1997.
- Madsen, Welles, 2000.
- Madsen, Welles, 2000, pp. 2, 21.
- Britt et al., 2000, pp. 32A.
- Rauhut, 2003, pp. 25.
- Carrano, Sampson, 2008.
- Carrano, Sampson, 2008, pp. 192.
- Madsen, Welles, 2000, pp. 36.
- Britt et al., 1999.
- Foster, Appendix, 2007.
- Zils, 1995.
- Janensch, 1925.
- Rowe, Gauthier, 1990.
- Tykoski, Rowe, 2004.
- Rauhut, 2003.
- Tetanurae (англ.) информация на сайте Paleobiology Database. (Дата обращения: 14 февраля 2018)
- Rauhut, 2011.
- Mateus, 2000.
- Mateus, 2006.
- Malafaia, 2015.
- Marsh, 1892, pp. 343—349.
- Therrien, 2007.
- Tykoski, Rowe, 2004, pp. 66.
- Paul, 2016, pp. 84.
- Paul, 1988.
- Foster, Gargantuan to Minuscule, 2007.
- Gilmore, 1920, pp. 115.
- Carrano, Choiniere, 2016.
- Gilmore, 1920, pp. 104, 112.
- Gilmore, 1920, pp. 112.
- Gilmore, 1920, pp. 113—114.
- Madsen, Welles, 2000, pp. 32.
- Madsen, Welles, 2000, pp. 3.
- Paul, 1988, pp. 277.
- Gilmore, 1920, pp. 82.
- Gilmore, 1920, pp. 92.
- Madsen, Welles, 2000, pp. 3, 27.
- Rauhut, 2004.
- Carrano, Sampson, 2008, pp. 185.
- Hendrickx, 2015.
- Xu et al., 2009.
- Pol, Rauhut, 2012.
- Wang et al., 2017, pp. 147.
- Gianechini et al., 2015.
- Huene, 1926.
- Rauhut et al., 2010.
- Henderson, 1998.
- Foster, 2003, pp. 29.
- Bakker, Bir, 2004.
- Henderson, 1998, pp. 220.
- Chure, 1998.
- Marsh, 1884, pp. 331.
- Norman, 1985.
- Paul, 1988, pp. 274—279.
- Senter, 2009.
- Molnar, 2001.
- Rothschild, 2001.
- Trujillo, 2006.
- Bilbey, 1998.
- Russell, 1989.
- Mateus O., 2006.
- Chure et al., 2006.
- Dodson, 1980.
- Foster, 2003.
- Carpenter, 2006.
- Mateus, 2014.
- Glut, Brett-Surman, 1997.
Литература
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986. — 831 с. — 100 000 экз.
- Нэйш Д., Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. Александр Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2019. — 223 с. — ISBN 978-5-91671-940-6.
- Bakker R. T., Bir G. Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness // Feathered Dragons: Studies on the Transition from Dinosaurs to Birds / P. J. Currie, E. B. Koppelhus, M. A. Shugar, J. L. Wright. — Bloomington and Indianapolis: Indiana University Press, 2004. — P. 301—342. — ISBN 978-0-253-34373-4.
- Bilbey S. A. Cleveland-Lloyd Dinosaur Quarry – age, stratigraphy and depositional environments // The Morrison Formation: An Interdisciplinary Study / K. Carpenter, D. Chure, J. I. Kirkland. — Taylor and Francis Group, 1998. — P. 87—120. — (Modern Geology 22). — ISBN 0026-7775.
- Brinkman P. D. The Second Jurassic Dinosaur Rush. Museums and Paleontology in America at the Turn of the Twentieth Century. — University of Chicago Press, 2010. — P. 10. — ISBN 978-0-226-07472-6.
- Britt B. B., Miles C. A., Cloward K. C., Madsen J. H. A juvenile Ceratosaurus (Theropoda, Dinosauria) from Bone Cabin Quarry West (Upper Jurassic, Morrison Formation), Wyoming (англ.) // Journal of Vertebrate Paleontology. — 1999. — Vol. 19 (Supplement to No 3). — P. 33A.
- Britt B. B., Chure D. J., Holtz T. R. Jr., Miles C. A., Stadtman K. L. A reanalysis of the phylogenetic affinties of Ceratosaurus (Theropoda, Dinosauria) based on new specimens from Utah, Colorado, and Wyoming (англ.) // Journal of Vertebrate Paleontology. — 2000. — Vol. 20 (suppl.). — P. 32A. — doi:10.1080/02724634.2000.10010765.
- Carpenter K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus // Paleontology and Geology of the Upper Jurassic Morrison Formation / Foster J. R., Lucas S. G.. — Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 2006. — P. 131—138.
- Carrano M. T., Sampson S. D. The Phylogeny of Ceratosauria (Dinosauria: Theropoda) (англ.) // Journal of Systematic Palaeontology. — 2008. — Vol. 6, no. 2. — P. 183—236. — doi:10.1017/S1477201907002246.
- Carrano M. T., Choiniere J. New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution (англ.) // Journal of Vertebrate Paleontology. — 2016. — Vol. 36, no. 2. — doi:10.1080/02724634.2015.1054497.
- Chure D. J. Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic (англ.) // Gaia. — 1998. — December (vol. 15). — P. 227—232. — ISSN 0871-5424. Архивировано 19 июля 2011 года.
- Chure D. J., Litwin R., Hasiotis S. T., Evanoff E., Carpenter K. The fauna and flora of the Morrison Formation: 2006 // Paleontology and Geology of the Upper Jurassic Morrison Formation / Foster J. R., Lucas S. G.. — Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 2006. — P. 233—248.
- Dodson P., Behrensmeyer A. K., Bakker R. T., McIntosh J. S. Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation (англ.) // Paleobiology. — 1980. — Vol. 6, no. 2. — P. 208—232.
- Foster J. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. // New Mexico Museum of Natural History and Science Bulletin, 23. — Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 2003. — P. 29.
- Foster J. Appendix // Jurassic West: The Dinosaurs of the Morrison Formation and Their World. — Indiana University Press, 2007. — P. 327—329.
- Foster J. Gargantuan to Minuscule: The Morrison Menagerie, Part II // Jurassic West: The Dinosaurs of the Morrison Formation and Their World. — Indiana University Press, 2007. — P. 162—242. — ISBN 0-253-34870-6.
- Gianechini F. A., Apestteguia S., Landini W., Finotti F., Valieri R. J., Zandonai F. New abelisaurid remains from the Anacleto Formation (Upper Cretaceous), Patagonia, Argentina (англ.) // Cretaceous Research. — 2015. — Vol. 54. — P. 1—16. — doi:10.1016/j.cretres.2014.11.009.
- Gilmore C. W. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus (англ.) // Bulletin of the United States National Museum. — 1920. — Vol. 110, no. 110. — P. 1—154. — doi:10.5479/si.03629236.110.i.
- Glut D. F. Ceratosaurus // Dinosaurs: The Encyclopedia. — McFarland & Company, 1997. — P. 266—270. — ISBN 0-89950-917-7.
- Glut D. F., Brett-Surman M. K. Dinosaurs and the media // The Complete Dinosaur. — Bloomington and Indianapolis: Indiana University Press, 1997. — P. 675—706. — ISBN 0-253-33349-0.
- Henderson D. M. Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods (англ.) // Gaia. — 1998. — Vol. 15. — P. 219—226. — ISSN 0871-5424.
- Hendrickx C., Hartman S. A., Mateus O. An overview of non-avian theropod discoveries and classification (англ.) // PalArch's Journal of Vertebrate Palaeontology. — 2015. — Vol. 12, no. 1. — P. 1—73. — ISSN 1567-2158.
- Huene F. v. On several known and unknown reptiles of the order Saurischia from England and France (англ.) // Annals and Magazine of Natural History. — 1926. — Vol. 9, no. 17. — P. 473—489.
- Janensch W. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas (нем.) // Palaeontographica. — 1925. — Bd. Supplement VIII. — S. 1—100.
- Marsh O. C. Principal characters of American Jurassic dinosaurs, part VIII: The order Theropoda (англ.) // American Journal of Science. — 1884. — Vol. 24, no. 160. — P. 329—340. — doi:10.2475/ajs.s3-27.160.329.
- Marsh O. C. Restorations of Claosaurus and Ceratosaurus (англ.) // American Journal of Science. — 1892. — Vol. 44, no. 262. — P. 343—349. — doi:10.2475/ajs.s3-44.262.343.
- Madsen J. H., Welles S. P. Ceratosaurus (Dinosauria, Theropoda): A Revised Osteology. — Utah Geological Survey, 2000. — 80 p. — ISBN 1-55791-380-3.
- Malafaia E., Ortega F., Escaso F., Silva B. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal (англ.) // Historical Biology. — 2015. — Vol. 27, no. 7. — P. 938—946. — doi:10.1080/08912963.2014.915820.
- Mateus O., Antunes M. T. Ceratosaurus (Dinosauria: Theropoda) in the Late Jurassic of Portugal (англ.) // 31st International Geological Congress. — 2000. — Vol. Abstract Volume.
- Mateus O., Walen A., Antunes M. T. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus (англ.) // New Mexico Museum of Natural History and Science Bulletin. — 2006. — Vol. 36.
- Mateus O. Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison // Paleontology and Geology of the Upper Jurassic Morrison Formation / Foster J. R., Lucas S. G.. — Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 2006. — P. 223—231.
- Mateus O., Mannion Ph. D., Upchurch P. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal (англ.) // Journal of Vertebrate Paleontology. — 2014. — Vol. 34, no. 3. — P. 618—634. — doi:10.1080/02724634.2013.822875.
- Molnar R. E. Theropod paleopathology: a literature survey // Mesozoic Vertebrate Life / D. H. Tanke, K. Carpenter. — Bloomington: Indiana University Press, 2001. — P. 337—363. — 577 p. — ISBN 978-0-253-33907-2.
- Norman D. B. Carnosaurs // The Illustrated Encyclopedia of Dinosaurs. — Salamander Books Ltd, 1985. — P. 62—67. — ISBN 0-517-46890-5.
- Paul G. S. Ceratosaurs // Predatory Dinosaurs of the World. — NY: Simon & Schuster, 1988. — P. 274—279. — ISBN 0-671-61946-2.
- Paul G. S. The Princeton Field Guide to Dinosaurs (2nd ed.). — Princeton: Princeton University Press, 2016. — 362 p. — ISBN 978-0-691-16766-4. Архивная копия от 2 июля 2018 на Wayback Machine
- Pol D., Rauhut O. W. M. A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs (англ.) // Proceedings of the Royal Society B: Biological Sciences. — 2012. — Vol. 279, no. 1804. — P. 3170—3175. — doi:10.1098/rspb.2012.0660.
- Russell D. A. An Odyssey in Time: Dinosaurs of North America. — Minocqua, Wisconsin: NorthWord Press, 1989. — P. 64—70. — ISBN 978-1-55971-038-1.
- Rauhut O. W. M. The Interrelationships and Evolution of Basal Theropod Dinosaurs (англ.) // Special Papers in Palaeontology. — 2003. — Vol. 69.
- Rauhut O. W. M. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia (англ.) // Journal of Vertebrate Paleontology. — 2004. — Vol. 24, no. 4. — P. 894—902. — doi:10.1671/0272-4634(2004)024[0894:paaogs]2.0.co;2.
- Rauhut O. W. M., Milner A. C., Moore-Fay S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England (англ.) // Zoological Journal of the Linnean Society. — 2010. — Vol. 158, no. 1. — P. 155—195. — doi:10.1111/j.1096-3642.2009.00591.x.
- Rauhut O. W. M. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania) (англ.) // Special Papers in Palaeontology. — 2011. — Vol. 86. — P. 195—239.
- Rothschild B., Tanke D. H., Ford T. L. Theropod stress fractures and tendon avulsions as a clue to activity // Mesozoic Vertebrate Life / D. H. Tanke, K. Carpenter. — Bloomington: Indiana University Press, 2001. — P. 331—336. — ISBN 978-0-253-33907-2.
- Rowe T., Gauthier J. Ceratosauria // The Dinosauria / Weishampel D. B., Dodson P., Osmólska H.. — University of California Press, 1990. — P. 151—168. — ISBN 0-520-06726-6.
- Senter P. Vestigial skeletal structures in dinosaurs (англ.) // Journal of Zoology. — 2009. — Vol. 280. — P. 60—71. — doi:10.1111/j.1469-7998.2009.00640.x.
- Therrien F., Henderson D. M. My theropod is bigger than yours... or not: estimating body size from skull length in theropods (англ.) // Journal of Vertebrate Paleontology. — 2007. — Vol. 27, no. 1. — P. 108—115. — doi:10.1671/0272-4634(2007)27[108:mtibty]2.0.co;2.
- Trujillo K. C., Chamberlain K. R., Strickland A. Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations (англ.) // Geological Society of America Abstracts with Programs. — 2006. — Vol. 38, no. 6. — P. 7.
- Tykoski R. S., Rowe T. Ceratosauria // The Dinosauria: Second Edition / David B. Weishampel, Peter Dodson, Halszka Osmólska. — University of California Press, 2004. — P. 47—70. — ISBN 0-520-24209-2.
- Wang S., Stiegler J., Amiot R., Wang X., Du G.-H., Clark J. M., Xu X. Extreme ontogenetic changes in a ceratosaurian theropod (англ.) // Cell Biology. — 2017. — Vol. 27. — P. 144—148. — doi:10.1016/j.cub.2016.10.043.
- Xu X., Clark J. M., Mo J., Choiniere J., Forster C. A., Erickson G. M., Hone D. W., Sullivan C., Eberth D. A., Nesbitt S., Zhao Q. A Jurassic ceratosaur from China helps clarify avian digital homologies (англ.) // Nature. — 2009. — Vol. 459, no. 7249. — P. 940—944. — doi:10.1038/nature08124.
- Zils Ch. W., Moritz A., Saanane Ch. Tendaguru, the most famous dinosaur locality of Africa. Review, survey and future prospects (англ.) // Documenta naturae. — 1995. — Vol. 97. — P. 1—41.
Ссылки
- The Good, Bad and Ugly: Dismantling Historic Fossil Displays, Part Two (недоступная ссылка). Digging the Fossil Record: Paleobiology at the Smithsonian. Smithsonian National Museum of Natural History. Дата обращения: 13 февраля 2018. Архивировано 10 октября 2017 года.
- David Koch donates $35 million to National Museum of Natural History for dinosaur hall. Washington Post. Дата обращения: 13 февраля 2018.