Целурозавры
Целуроза́вры[2] (лат. Coelurosauria) — самая многочисленная группа тероподовых динозавров, объединяющая все таксоны, более близкие к птицам, чем к карнозаврам[3]. Возникают в палеонтологической летописи в средней юре. К целурозаврам относятся тираннозавроиды (Tyrannosauroidea), компсогнатиды (Compsognathidae), орнитомимозавры (Ornithomimosauria) и манирапторы (Maniraptora)[4][5][6][7][8]; в состав последних, помимо нескольких вымерших таксонов, входят птицы (Aves) — единственная современная группа целурозавров[9][5][6][7][10].
| Целурозавры | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 По часовой стрелке, начиная сверху слева: овирапторид GIN 100/42 (читипати?), синозавроптерикс, Nothronychus mckinleyi, тираннозавр, бэмбираптор, домовый воробей, струтиомим и микрораптор гуи | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Клада: Клада: Целурозавры |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Coelurosauria von Huene, 1914 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
по The Theropod Database[1]:
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 167,7 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Отличительные черты
Согласно исследованию А. Кау 2018 года, целурозавры отличимы от других теропод по следующим апоморфиям[6]:
- отчётливый задневентральный отросток слёзной кости;
- дистальная поверхность лобковой ножки субпрямоугольная;
- задняя (постериорная) часть лобковой ножки удлинённая;
- дистальная половина диафиза плюсневой кости IV соприкасается с плюсневой костью III.
Кроме того, для целурозавров характерны следующие признаки[4]:
- мозг не менее чем в два раза больше, чем у других теропод при тех же размерах тела;
- длинные стройные трёхпалые кисти;
- шевроны имеют форму каяка, что делает дистальную часть хвоста особенно тонкой;
- длинные узкие плюсневые кости.
Эволюционная история

Первые остатки целурозавров найдены в отложениях, датируемых средней юрой. Древнейшими родами, достоверно относящимися к целурозаврам, являются тираннозавроиды процератозавр[11] и килеск[12][13], жившие на территории Азии около 167,7—164,7 млн лет назад (окаменелости первого обнаружены в Англии, второго — в Сибири)[14][15].
Хорошо сохранившимся материалом представлен примитивный целурозавр Bicentenaria[16], живший на территории нынешней Аргентины в позднем («среднем») мелу, около 99,7—94,3 млн лет назад[17]. Это был небольшой стройный динозавр, достигавший 2—4 м в длину, который, судя по тонким конечностям и узкому динамичному стабилизирующему хвосту, был весьма проворными. Исходя из наличия узких хватательных передних конечностей, можно предположить, что Bicentenaria и прочие примитивные целурозавры специализировались на ловле небольшой добычи[4]. Кроме того, к числу базальных целурозавров относят Zuolong[18] и, в некоторых классификациях, Aorun[19] (по другой версии — примитивный альваресзаврид[20]) из среднеюрских отложений Китая, а также раннемеловых Aratasaurus (Бразилия)[21] и Vayuraptor (Таиланд)[22].
Хотя древнейшие целурозавры были небольшими активными хищниками, их потомки быстро разделились на множество морфологически и экологически отличных подгрупп. Зубы базальных целурозавров зифодонтные, т. е. уплощены с боков и имеют режущие пильчатые края. У продвинутых целурозавров — манирапториформ (орнитомимозавры и манирапторы) — зубы приобретают листовидную форму (англ. phyllodont teeth), либо утрачиваются вовсе, часто отсутствуют хватательные когти, а череп обладает сравнительно небольшими размерами. Судя по этим признакам, манирапториформы отходили от строго плотоядной диеты своих предков и родственников[4][23].

Эволюционная история перьев во многом неясна. Отпечатки примитивных нитеобразных перьев обнаружены у некоторых птерозавров, птицетазовых и многих теропод. Таким образом, согласно филогенетическому брекетингу, протоперья могли появиться у орнитодир ещё в раннем триасе и имелись уже у ближайшего общего предка птерозавров и динозавров. С другой стороны, известны многочисленные отпечатки чешуи динозавров; если у их предка были протоперья, то, возможно, они были вторично утрачены в некоторых группах. Наибольшее разнообразие перьев наблюдается у целурозавров. У базальных целурозавров, таких как тираннозавроиды ютираннус[25] и дилун[26], а также компсогнатиды синозавроптерикс[27] и "Ubirajara"[28], имелись примитивные нитеподобные перья. У манирапторов появляются контурные перья, как у современных птиц[24]. Известно множество отпечатков чешуи тираннозаврид, что, в сочетании с наличием перьев у более базальных тираннозавроид, свидетельствует в пользу того, что тираннозавриды вторично утратили перьевой покров (возможно, из-за увеличения размеров тела)[29]. Не исключено, что у некоторых других крупных целурозавров, таких как дейнохейриды и теризинозавриды, также почти или полностью отсутствовал перьевой покров, поскольку он был лишними с точки зрения их теплоэнергетики, но проверить данное утверждение в настоящее время невозможно из-за отсутствия отпечатков покровов этих животных[30].
Основные группы целурозавров возникли в результате сравнительно быстрой дивергенции. В ходе такой эволюционной радиации появились и первые авиалы (птицы в широком смысле). Один из наиболее известных базальных авиал — археоптерикс — жил в поздней юре. Как показывают некоторые недавние анализы, к авиалам может также относиться семейство Anchiornithidae, которое, предположительно, занимает более базальную позицию, чем археоптерикс[31][32][33][34][7]; представители этого семейства существовали в средней юре, около 160 млн лет назад[35]. Все целурозавры, кроме веерохвостых птиц (т. е. птиц в узком смысле), вымерли в ходе мел-палеогенового вымирания, произошедшего 66 млн лет назад[31].
Классификация
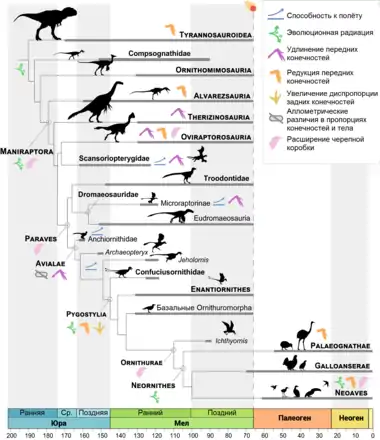
Ж. А. Готье (1986) определил кладу Coelurosauria как объединяющую птиц и все таксоны тетануров, более близкие к птицам, чем к карнозаврам[3]. Уточнённую версию этого определения дали Т. Р. Хольц, Р. Е. Мольнар и Ф. Д. Карри (2004); в качестве спецификатора птиц исследователи выбрали домового воробья (Passer domesticus), тогда как на роль представителя карнозавров был выбран аллозавр (Allosaurus fragilis)[36]. Почти все целурозавры, кроме наиболее базальных родов, входят в состав клады Tyrannoraptora, определяемой как наименее инклюзивная клада, включающая тираннозавра и современных птиц[37] (домового воробья[36]).
Спорное положение в пределах группы тетануров занимает клада Megaraptora, в разных исследованиях относимая к аллозавроидам (карнозаврам) семейства Neovenatoridae[38][39][40], к тираннозавроидам[41][42] или к базальным целурозаврам (вне клады Tyrannoraptora)[43][44]. В составе Tyrannoraptora выделяют следующие основные подгруппы: тираннозавроиды, компсогнатиды, орнитомимозавры и манирапторы[5][7].
Тираннозавроиды просуществовали со средней юры до конца мела, став наиболее успешной группой базальных целурозавров. На протяжении большей части своей эволюционной истории тираннозавроиды были в основном небольшими оперёнными плотоядными животными (на фоне своих мелких родственников выделяются ютираннус[25] и синотираннус[45], достигавшие 7,5—8,5 м в длину[46]). Только после того как ниши высших хищников Северной Америки и Азии, которые прежде занимали кархародонтозавриды и спинозавриды, оказались освобождены, продвинутые тираннозавроиды семейства тираннозаврид сумели стать успешными сверххищниками. Тираннозавриды жили 80—66 млн лет назад[4].
Компсогнатиды — это мелкие целурозавры, вероятно, более продвинутые (в плане близости к птицам), чем тираннозавроиды[5][6][7], хотя и весьма примитивные по своей морфологии. Компсогнатиды обитали на территории Азии, Европы и Южной Америки с верхней юры по ранний мел, в длину достигали от 1 м (компсогнат) до 2,5 м (синокаллиоптерикс)[4].
Клада манирапториформ подразделяется на орнитомимозавров и манирапторов. Орнитомимозавры — страусоподобные растительноядные или всеядные динозавры мелового периода, наиболее заметными чертами которых были удлинённая шея с небольшой головой на ней и, у продвинутых представителей, беззубый клюв[4]. Манирапторов обычно подразделяют на клады Alvarezsauria, Therizinosauria (теризинозавры), Oviraptorosauria (овирапторозавры), Scansoriopterygidae (скансориоптеригиды), Deinonychosauria (дейнонихозавры; иногда считаются парафилетической группой[5]) и Avialae (авиалы)[9]; последний таксон включает около 10 912 современных видов птиц[47] (в составе кроновой группы Aves[48][31] / Neornithes[7]), что делает его самой успешной группой целурозавров и динозавров в целом[31][9].
Филогения
Кладограмма согласно филогенетическому анализу 2020 года (Sayao et al., 2020)[21]:
| Avetheropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
.png.webp)

Примечания
- Mortimer M. Coelurosauria (англ.). The Theropod Database. Дата обращения: 26 сентября 2021.
- Основы палеонтологии : Справочник для палеонтологов и геологов СССР : в 15 т. / гл. ред. Ю. А. Орлов. — М. : Наука, 1964. — Т. 12 : Земноводные, пресмыкающиеся и птицы / под ред. А. К. Рождественского, Л. П. Татаринова. — С. 530. — 724 с. — 3000 экз.
- Gauthier J. A. Saurischian monophyly and the origin of birds (англ.) // The Origin of Birds and the Evolution of Flight / In K. Padian. — San Francisco, CA: Memoirs of the California Academy of Sciences, 1986. — Vol. 8. — P. 1—55.
- Theropoda II, Coelurosauria: Tyrant Kings and Lesser Royals (англ.). GEOL 104 Dinosaurs: A Natural History. Дата обращения: 26 сентября 2021. Архивировано 15 января 2021 года.
- Hendrickx C., Hartman S. A., Mateus O. An Overview of Non-Avian Theropod Discoveries and Classification (англ.) // PalArch’s Journal of Vertebrate Palaeontology : journal. — 2015. — Vol. 12, iss. 1. — P. 1—73.
- Cau A. The assembly of the avian body plan: a 160-million-year long process (англ.) // Bollettino della Società Paleontologica Italiana : journal. — 2018. — Vol. 57, iss. 1. — P. 1–25. — ISSN 0375-7633. — doi:10.4435/BSPI.2018.01.
- Nebreda S. M., Fernández M. H., Marugán-Lobón J. ‘Dinosaur-bird’ macroevolution, locomotor modules and the origins of flight (англ.) // Journal of Iberian Geology. — 2021. — Vol. 47, iss. 3. — P. 565—574. — ISSN 1886-7995. — doi:10.1007/s41513-021-00170-3.
- Нэйш и Барретт, 2019, с. 50—52.
- Theropoda III, Maniraptora: The Feathered Dinosaurs (англ.). GEOL 104 Dinosaurs: A Natural History. Дата обращения: 26 сентября 2021. Архивировано 26 сентября 2021 года.
- Нэйш и Барретт, 2019, с. 52—57.
- Rauhut O. W. M., Milner A. C., Moore-Fay S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England (англ.) // Zoological Journal of the Linnean Society. — 2010. — Vol. 158, iss. 1. — P. 155—195. — ISSN 0024-4082. — doi:10.1111/j.1096-3642.2009.00591.x.
- Averianov A. O., Krasnolutskii S. A., Ivantsov S. V. A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia (англ.) // Proceedings of the Zoological Institute. — 2010. — Vol. 314, no. 1. — P. 42—57.
- Averianov A. O., Osochnikova A., Skutschas P., Krasnolutskii S., Schellhorn R., Schultz J. A., Martin T. New data on the tyrannosauroid dinosaur Kileskus from the Middle Jurassic of Siberia, Russia (англ.) // Historical Biology : journal. — 2021. — Vol. 33, iss. 7. — P. 897—903. — ISSN 0891-2963. — doi:10.1080/08912963.2019.1666839.
- Proceratosaurus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 21 сентября 2021).
- Kileskus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 21 сентября 2021).
- Novas F., Ezcurra M., Agnolin F., Pol D., Ortiz R. New Patagonian Cretaceous theropod sheds light about the early radiation of Coelurosauria (англ.) // Revista del Museo Argentino de Ciencias Naturales nueva serie : journal. — 2012. — Vol. 14, iss. 1. — P. 57—81. — ISSN 1853-0400. — doi:10.22179/revmacn.14.372.
- Bicentenaria (англ.) информация на сайте Paleobiology Database. (Дата обращения: 21 сентября 2021).
- Choiniere J. N., Clark J. M., Forster C. A., Xu X. A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People's Republic of China (англ.) // Journal of Vertebrate Paleontology. — 2010. — Vol. 30, iss. 6. — P. 1773—1796. — ISSN 0272-4634. — doi:10.1080/02724634.2010.520779.
- Choiniere J. N., Clark J. M., Forster C. M., Norella M. A., Eberth D. A., Erickson G. M., Chu H., Xu X. A juvenile specimen of a new coelurosaur (Dinosauria: Theropoda) from the Middle–Late Jurassic Shishugou Formation of Xinjiang, People's Republic of China (англ.) // Journal of Systematic Palaeontology. — 2014. — Vol. 12, iss. 2. — P. 177—215. — ISSN 1477-2019. — doi:10.1080/14772019.2013.781067. Архивировано 11 января 2019 года.
- Xu X., Choiniere J., Tan Q., Benson R. B. J., Clark J., Sullivan C., Zhao Q., Han F., Ma Q., He Y., Wang S., Xing H., Tan L. Two Early Cretaceous Fossils Document Transitional Stages in Alvarezsaurian Dinosaur Evolution (англ.) // Current Biology : journal. — 2018. — Vol. 28, iss. 17. — P. 2853—2860.e3. — ISSN 0960-9822. — doi:10.1016/j.cub.2018.07.057.
- Sayao J. M., Saraiva A. Á. F., Brum A. S., Bantim R. A. M., de Andrade R. C. L. P., Cheng X., de Lima F. J., de Paula Silva H., Kellner A. W. A. The first theropod dinosaur (Coelurosauria, Theropoda) from the base of the Romualdo Formation (Albian), Araripe Basin, Northeast Brazil (англ.) // Scientific Reports : journal. — 2020. — Vol. 10, iss. 1. — P. 10892. — ISSN 2045-2322. — doi:10.1038/s41598-020-67822-9.
- Samathi A., Chanthasit P., Sander P. Two new basal coelurosaurian theropod dinosaurs from the Early Cretaceous Sao Khua Formation of Thailand (англ.) // Acta Palaeontologica Polonica : journal. — 2019. — Vol. 64. — P. 239—260. — ISSN 0567-7920. — doi:10.4202/app.00540.2018.
- Zanno L. E., Makovicky P. J. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution (англ.) // Proceedings of the National Academy of Sciences : journal. — 2011. — Vol. 108, iss. 1. — P. 232—237. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.1011924108.
- Benton M. J., Dhouailly D., Jiang B., McNamara M. The Early Origin of Feathers (англ.) // Trends in Ecology & Evolution : journal. — 2019. — Vol. 34, iss. 9. — P. 856—869. — ISSN 0169-5347. — doi:10.1016/j.tree.2019.04.018. — PMID 31164250.
- Xu X., Wang K., Zhang K., Ma Q., Xing L., Sullivan C., Hu D., Cheng S., Wang S. A gigantic feathered dinosaur from the Lower Cretaceous of China (англ.) // Nature : journal. — 2012. — Vol. 484, iss. 7392. — P. 92—95. — ISSN 1476-4687. — doi:10.1038/nature10906.
- Xu X., Norell M. A., Kuang X., Wang X., Zhao Q., Jia C. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids (англ.) // Nature : journal. — 2004. — Vol. 431, iss. 7009. — P. 680—684. — ISSN 1476-4687. — doi:10.1038/nature02855. — PMID 15470426. Архивировано 17 августа 2016 года.
- Currie P. J., Chen P.-J. Anatomy of Sinosauropteryx prima from Liaoning, northeastern China (англ.) // Canadian Journal of Earth Sciences. — 2001. — Vol. 38, no. 12. — P. 1705—1727. — ISSN 1480-3313. — doi:10.1139/e01-050.
- Smyth R. S. H., Martill D. M., Frey E., Rivera-Silva H. E., Lenz N. TEMPORARY REMOVAL: A maned theropod dinosaur from Gondwana with elaborate integumentary structures (англ.) // Cretaceous Research : journal. — 2020 online. — No. 104686. — P. 1—20. — doi:10.1016/j.cretres.2020.104686.
- Bell P. R., Campione N. E., Persons W. S., Currie P. J., Larson P. L., Tanke D. H., Bakker R. T. Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution (англ.) // Biology Letters : journal. — 2017. — Vol. 13, iss. 6. — P. 20170092. — ISSN 1744-957X. — doi:10.1098/rsbl.2017.0092.
- Марк Уиттон. Пух гигантов под микроскопом: насколько достоверны пушистые реконструкции гигантских вымерших животных?. PaleoNews (15 мая 2020). Дата обращения: 26 сентября 2021. Архивировано 26 сентября 2021 года.
- Theropoda IV, Avialae: Rise of the Birds (англ.). GEOL 104 Dinosaurs: A Natural History. Дата обращения: 26 сентября 2021. Архивировано 26 сентября 2021 года.
- Cau A., Beyrand V., Voeten D. F. A. E., et al. Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs (англ.) // Nature : journal. — 2017. — Vol. 552, iss. 7685. — P. 395—399. — ISSN 1476-4687. — doi:10.1038/nature24679. — . — PMID 29211712.
- Rauhut O. W. M., Tischlinger H., Foth C. A non-archaeopterygid avialan theropod from the Late Jurassic of southern Germany (англ.) // eLife : journal. — 2019. — Vol. 8. — P. e43789. — ISSN 2050-084X. — doi:10.7554/eLife.43789. — PMID 31084702.
- Pei R., Pittman M., Goloboff P. A., et al. Potential for Powered Flight Neared by Most Close Avialan Relatives, but Few Crossed Its Thresholds (англ.) // Current Biology : journal. — 2020. — Vol. 30, iss. 20. — P. 4033—4046.e8. — ISSN 0960-9822. — doi:10.1016/j.cub.2020.06.105.
- Chu Z., He H., Ramezani J., Bowring S. A., Hu D. High-precision U-Pb geochronology of the Jurassic Yanliao Biota from Jianchang (western Liaoning Province, China): Age constraints on the rise of feathered dinosaurs and eutherian mammals (англ.) // Geochemistry, Geophysics, Geosystems. — 2016. — Vol. 17, iss. 10. — P. 3983—3992. — ISSN 1525-2027. — doi:10.1002/2016GC006529.
- Weishampel, Dodson & Osmolska, 2004, chpt. 4: "Basal Tetanurae" by T. R. Holtz Jr., R. E. Molnar, P. J. Currie, pp. 71—110.
- Sereno P. C. The Evolution of Dinosaurs (англ.) // Science : journal. — 1999. — Vol. 284, no. 5423. — P. 2137—2147. — ISSN 0036-8075. — doi:10.1126/science.284.5423.2137. — PMID 10381873.
- Benson R. B. J., Carrano M. T., Brusatte S. L. A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic (англ.) // Naturwissenschaften : journal. — 2010. — Vol. 97, iss. 1. — P. 71—78. — ISSN 1432-1904. — doi:10.1007/s00114-009-0614-x. — . — PMID 19826771.
- Carrano M. T., Benson R. B. J., Sampson S. D. The phylogeny of Tetanurae (Dinosauria: Theropoda) (англ.) // Journal of Systematic Palaeontology : journal. — 2012. — Vol. 10, iss. 2. — P. 211—300. — ISSN 1477-2019. — doi:10.1080/14772019.2011.630927.
- Zanno L. E., Makovicky P. J. Neovenatorid theropods are apex predators in the Late Cretaceous of North America (англ.) // Nature Communications : journal. — 2013. — Vol. 4, iss. 1. — P. 2827. — ISSN 2041-1723. — doi:10.1038/ncomms3827.
- Novas F. E., Agnolín F. L., Ezcurra M. D., Canale J. I., Porfiri J. D. Megaraptorans as members of an unexpected evolutionary radiation of tyrant-reptiles in Gondwana (англ.) // Ameghiniana : journal. — 2012. — Vol. 49 (Suppl), no. 4. — P. R33.
- Porfiri J. D., Novas F. E., Calvo J. O., Agnolín F. L., Ezcurra M. D., Cerda I. A. Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation (англ.) // Cretaceous Research : journal. — 2014. — Vol. 51. — P. 35—55. — ISSN 0195-6671. — doi:10.1016/j.cretres.2014.04.007.
- Porfiri J. D., Juárez Valieri R. D., Santos D. D. D., Lamanna M. C. A new megaraptoran theropod dinosaur from the Upper Cretaceous Bajo de la Carpa Formation of northwestern Patagonia (англ.) // Cretaceous Research : journal. — 2018. — Vol. 89. — P. 302—319. — ISSN 0195-6671. — doi:10.1016/j.cretres.2018.03.014.
- Delcourt R., Grillo O. N. Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology : journal. — 2018. — Vol. 511. — P. 379—387. — ISSN 0031-0182. — doi:10.1016/j.palaeo.2018.09.003. — .
- Ji Q., Ji S.-A., Zhang L.-J. First large tyrannosauroid theropod from the Early Cretaceous Jehol Biota in northeastern China (англ.) // Geological Bulletin of China : journal. — 2009. — Vol. 28, no. 10. — P. 1369—1374. Архивировано 1 октября 2018 года.
- Molina-Pérez & Larramendi, 2019, pp. 264—265.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Welcome (англ.). IOC World Bird List (v11.2) (15 июля 2021). doi:10.14344/IOC.ML.11.2.
- In de Queiroz, Cantino & Gauthier, 2020, Aves C. Linnaeus 1758 [J. A. Clarke, D. P. Mindell, K. de Queiroz, M. Hanson, M. A. Norell & J. A. Gautheir], converted clade name, pp. 1247—1253.
Литература
На русском языке
- Нэйш Д., Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. А. О. Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2019. — 223 с. — ISBN 978-5-91671-940-6.
На английском языке
- The Dinosauria (англ.) / D. B. Weishampel, P. Dodson, H. Osmolska, eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p. — ISBN 978-0-520-25408-4.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K., Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
- Molina-Pérez R., Larramendi A. Dinosaur Facts and Figures: The Theropods and Other Dinosauriformes (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press, 2019. — 288 p. — ISBN 978-0-691-18031-1.