Синозавроптерикс
Синозавропте́рикс[2][3] (лат. Sinosauropteryx, что означает «китайский крылатый ящер», кит. 中华龙鸟, пин. Zhōnghuá lóng niǎo, буквально: китайское крыло дракона) — монотипический род тероподовых динозавров из семейства компсогнатид, живших в раннемеловую эпоху (аптский век, 124,6—122 млн лет назад) на территории, которую сейчас занимает Китай. Описанный в 1996 году, стал первым таксоном за пределами клады авиал с доказательствами наличия перьевого покрова. При жизни эти динозавры были покрыты очень простыми нитевидными перьями. Отвечающие за окрас структуры также сохранились в некоторых перьях, поэтому синозавроптерикс стал первым динозавром с достоверно установленной расцветкой. Некоторые споры вызвала альтернативная интерпретация нитевидных вкраплений в качестве остатков коллагеновых волокон, однако такая интерпретация не была широко принята.
| † Синозавроптерикс | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Образец голотипа с отпечатками волокон, Музей Внутренней Монголии | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Клада: Клада: Клада: Семейство: Род: † Синозавроптерикс |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Sinosauropteryx Ji & Ji, 1996 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 124,6—122,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
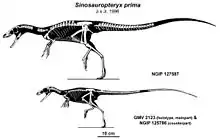
Синозавроптерикс был мелким тероподом с необычайно длинным хвостом и короткими передними конечностями. Самый крупный известный образец достигал длины 1,07 метра и имел предполагаемую массу в 0,55 килограмма. Этот динозавр являлся близким родственником похожего, но более раннего компсогната; оба рода включают в состав семейства компсогнатид. Известен единственный вид — Sinosauropteryx prima. Описаны три образца. Третий образец, пока соотнесённый с этим родом, может являться либо другим видом синозавроптерикса, либо другим родом динозавров.
Синозавроптерикс является одним из первых динозавров, найденных в формации Исянь (провинция Ляонин) и был представителем биоты Жэхэ. Хорошо сохранившиеся окаменелости этого вида иллюстрируют многие аспекты его палеобиологии, такие как рацион и размножение.
История открытия и изучения

Первый ископаемый образец динозавра, позднее названного Sinosauropteryx prima, обнаружили в августе 1996 китайские фермеры. Окаменелость располагалась на двух плитках, которые они продали двум разным научным организациям: Национальному геологическому музею в Пекине и Нанкинскому институту геологии и палеонтологии. Директор пекинского музея Цзи Цян подтвердил важность находки после встречи с канадским палеонтологом Филипом Карри и художником-иллюстратором Майклом Скрепником, которые узнали об уникальной окаменелости случайно во время знакомства с коллекцией музея в октябре 1996 года. Карри сразу понял исключительное значение образца. Как писала газета «The New York Times», цитируя Карри: «Когда я увидел плитку из окаменевшего ила, смешанную с вулканическим пеплом, в которой было заключено существо, я был поражён»[4].
Поначалу китайские власти запретили фотографировать образец. Тем не менее, Карри принёс фотографию на собрание Общества палеонтологии позвоночных, состоявшееся в Американском музее естественной истории в Нью-Йорке, что привело к бурному обсуждению нового открытия собравшимися палеонтологами. Сообщалось, что новость об открытии оперённого динозавра привела палеонтолога Джона Острома, который в 1970-е годы впервые высказал мысль о происхождении птиц от динозавров, «в состояние шока»[4]. Позднее Остром вошёл в состав международной команды, собравшейся в Пекине для изучения окаменелостей; другими членами команды были Алан Браш, эксперт по перьям, Ларри Мартин, эксперт по ископаемым птицам, и Петер Велльнхофер, эксперт по археоптериксу[5].
В 1998 году голотип GMV 2123 был описан китайскими палеонтологами Цян Цзи и Шуан Цзи как новый вид Sinosauropteryx prima. Родовое название образовано от слов лат. Sino — китайский, греч. σαῦρος — рептилия, и πτέρυξ — крыло, с отсылкой, что найденный образец, по мнению авторов описания, принадлежит переходной форме между тероподами и птицами. Видовое название в переводе с латинского означает «первая»[6].
С видом Sinosauropteryx prima были соотнесены три образца: голотип (и его контр-плита NIGP 127586), NIGP 127587 и D 2141[7]. Все окаменелости были найдены в разрезах Цяншангоу или Даванчжанцзы формации Исянь (уезды Бэйпяо и Линъюань, провинция Ляонин, Китай), которые датируют нижним аптским ярусом меловой системы, 124,6—122 млн лет назад[8].
Несмотря на перья, палеонтологи не посчитали синозавроптерикса птицей. Филогенетически род только отдалённо связан с кладой Aves, которая обычно определялась как Archaeopteryx lithographica плюс современные птицы. Авторы описания синозавроптерикса, тем не менее, использовали апоморфное определение класса птиц, согласно которому любое животное с перьями считается птицей. Исследователи утверждали, что нитчатые структуры динозавра представляли собой настоящие перья со стержнями и бородками, и потому синозавроптерикса следует считать настоящей птицей[9]. Они классифицировали новые род и вид в составе нового отряда Sinosauropterygiformes, семейства Sinosauropterygidae, в подклассе Sauriurae[6]. Такое филогенетическое размещение не было принято научным сообществом. В 2003 году немецкий палеонтолог Оливер Раухут по результатам филогенетического анализа отнёс динозавра к подсемейству Compsognathinae[10]:151. В следующем, 2004 году Фил Сентер сначала разместил синозавроптерикса среди базальных архозавров[11], но уже в следующей работе того же года отнёс его к целурозаврам[12]. С таким размещением впоследствии согласился и Раухут, разместив синозавроптерикса в политомии с целюром[13]. Тем не менее, в последующей статье 2011 года Филип Карри и Пей-Цзи Чен относят синозавроптерикса к семейству компсогнатид[14]:1708 — группе мелких длиннохвостых целурозавров, известных из верхнеюрских — нижнемеловых отложений Западной Европы и Китая[2].
Идентификация волокон
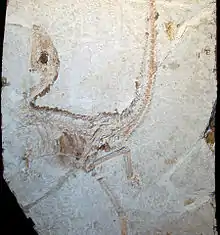
Споры по поводу нитевидных волокон, сохранившихся в первом экземпляре синозавроптерикса, начались почти сразу, как только международная команда учёных начала изучать пекинский образец под микроскопом. Результаты их исследований, сообщённые во время пресс-конференции в Филадельфийской академии естественных наук 24 апреля 1997 года, были неубедительными. Палеонтологи согласились, что структуры, сохранённые на образце, не были полноценными перьями, и предложили продолжить исследования[15]. Американский палеонтолог Алан Федуччиа утверждал в журнале «Aubodon», что нитевидные структуры на спине динозавра были жёстким «гребешком», и что палеонтологи принимали желаемое за действительное, приравнивая его к перьям[16]. В поздних публикациях некоторые учёные не соглашались с идентификацией структур в качестве нитевидных перьев[14][17].
Авторы некоторых публикаций последовали доводам Федуччии, который рассматривал нитевидные структуры образца в качестве коллагеновых волокон, а не примитивных перьев. Поскольку эти структуры явно располагались снаружи тела динозавра, было выдвинуто мнение, что волокна образовывали гребень на спине животного и нижней части хвоста, подобно таковому у некоторых современных водных ящериц[18][19]. Отсутствие перьев опровергло бы предположение, что синозавроптерикс является самым базальным известным оперённым тероподом, а также вызвало бы вопросы о нынешней теории происхождения перьев. Это поставило бы под сомнение гипотезу, согласно которой первые перья развились не для полёта, а для тепловой изоляции, а также то, что перья впервые появились в относительно базальных линиях динозавров, которые позже превратились в современных птиц[20].
Однако большинство учёных не согласилось с идентификацией структур в качестве коллагеновых волокон. Примечательно, что команда учёных, сообщивших о наличии пигментных клеток в структурах, утверждала: их присутствие уже доказывает, что структуры были перьями, а не коллагеном, поскольку коллаген не содержит пигмента[21][22]. Американский палеонтолог Грэгори С. Пол заявил, что сторонники гипотезы коллагеновых волокон рассматривают волокна не как составляющую тела динозавра, а в качестве дефекта препарации: линия разлома плиты и матовый герметик были ошибочно приняты за контур тела[23].
Гипотеза о наличии у древних рептилий коллагеновых волокон была тщательно проанализирована и опровергнута в научной работе 2017 года командой палеонтологов под руководством Ф. Смитвика. Покров синозавроптерикса сравнили с менее спорными доказательствами наличия коллагеновых волокон, сохранённых в образце ихтиозавра Stenopterygius. Несмотря на то, что сторонники коллагеновых волокон утверждали, что центральные стержни предполагаемых перьев теропода на самом деле были примером стержнеобразных коллагеновых нитей, изображение под сильным увеличением показало, что это не так. Предполагаемые стержни коллагена в образце ихтиозавра фактически оказались царапинами и трещинами, образовавшимися во время препарирования. Напротив, стержни на образце синозавроптерикса явились обычным примером фоссилизированных структур. Сторонники коллагена также утверждали, что на плитке с экземпляром теропода находились сферические образования, похожие на те, которые иногда встречаются в разлагающемся коллагене современных морских млекопитающих. Тем не менее, это заявление также не нашло подтверждения — команда Смитвика не обнаружила ни одной сферической структуры на образцах. Авторы работы предположили, что некоторые области окаменелости отбрасывали тени, которые на фотографиях с низким разрешением были похожи на сферические образования[24].
В результате исследования выяснилось, что другие предполагаемые фрагменты коллагеновых волокон, расположенные в области хвоста, оказались царапинами, аналогичными царапинам на других частях каменной плитки. Участки кости с неравномерной поверхностью считались доказательством того, что некоторые нити коллагена были разрушены больше, чем другие. Однако, в исследовании утверждалось, что после дальнейшей препарации эти неравномерные участки оказались всего лишь слоем осадка другого цвета. Коллагеновый «ореол», идентифицированный Федуччией, также был определён как осадок, окружающий один из экземпляров. Смитвик и его команда сделали вывод: покров, сохранённый на образцах синозавроптерикса, очень похож на таковой у птиц, чьи остатки найдены в той же формации[24].
Описание
Синозавроптерикс был мелким бипедальным тероподом, примечательным короткими передними конечностями, длинным первым пальцем кисти и очень длинным хвостом. Таксон основан на одних из самых маленьких из известных образцах нептичьих динозавров с голотипом всего 68 сантиметров длиной, включая хвост[25]:148. При этом данная особь была относительно молодой[14]. Самый крупный известный образец достигал длины 1,07 метра и имел предполагаемую массу в 0,55 килограмма[26].
Анатомически синозавроптерикс был очень схож с компсогнатом, отличаясь от него лишь пропорциями. Череп синозавроптерикса был на 15 % длиннее бедренной кости, в отличие от компсогната, у которого череп и бедро были почти равны по длине. Длина передней конечности синозавроптерикса (плечевой и локтевой кости) составляли только 30 % длины его ног (бедра и голени), по сравнению с 40 % у компсогната. Кроме того, у синозавроптерикса было несколько особенностей, уникальных среди всех других теропод. Его хвост состоял из 64 позвонков. Такое необычайно высокое количество позвонков сделало его хвост самым длинным по отношению к длине тела среди всех известных теропод[25]. Кисти этого динозавра также были длинными по сравнению с передними конечностями: они составляли от 84 % до 91 % длины остальной части лапы и половину длины стопы. Первый и второй пальцы были примерно одинаковой длины, первый палец был оснащён большим когтем[14]; при этом он был толще и длиннее, чем одна из костей предплечья[25]. Зубной аппарат синозавроптерикса был гетеродонтным: зубы на кончике верхней челюсти были тонкими и без зазубрин, а те, что располагались за ними, имели зазубрены и были сжаты с боков. Зубы нижней челюсти были аналогично дифференцированы[14].
Пигментированная область в брюшной полости голотипа рассматривалась как остатки внутренних органов[25]. Команда учёных под руководством Джона Рубена интерпретировала её как печень, которую они описали как часть крокодилоподобной дыхательной системы — «печёночный поршень»[27]. В более позднем исследовании, согласившись с тем, что пигментированная область изначально представляла собой нечто, расположенное внутри тела динозавра, учёные Филип Карри и Чен Пэй-цзи не нашли никакой определённой структуры. Авторы работы отметили, что при фоссилизации остатков любые органы будут искажены процессами, которые сплющивают скелет в двухмерный отпечаток[14]. Тёмный пигмент присутствует также в глазницах голотипа и другого образца[25].
Оперение

Все описанные образцы сохраняют покровные структуры, которые большинство палеонтологов интерпретирует как очень примитивный тип перьев. Эти короткие, пухоподобные нитевидные структуры расположены вдоль задней части черепа, конечностей, шеи, спины, верхней и нижней частей хвоста. Клочки перьев были обнаружены по бокам головы и тела, и авторы описания предположили, что плотность перьев на спине и случайное расположение клоков на боках указывает на то, что при жизни эти динозавры были полностью оперёнными, а большинство перьев на боках и брюхе отделилось в процессе разложения[25].
Нитевидные структуры сохранились в промежутках между костями, что, по мнению некоторых исследователей, соответствует ожидаемому количеству кожи и мышечной ткани, которые присутствовали на теле при жизни животного. Ближе всего к костям перья расположены на черепе и на конце хвоста, где было минимальное количество мышц, а дальше всего — в районе грудных позвонков, где мышц было больше. Это указывает на то, что нити размещались с внешней стороны тела и не являлись подкожными структурами[25].
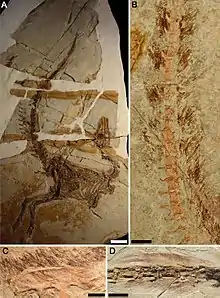
Нити показывают случайную ориентацию, местами они перекрывают друг друга и спутываются, что интерпретируется как свидетельство того, что они были мягкими и гибкими при жизни динозавра. Микроскопическое исследование показывает, что отдельные нити кажутся тёмными вдоль краёв и светлыми внутри — это свидетельствует о том, что внутри они были полыми, как и современные перья. По сравнению с шерстью современных млекопитающих, эти нити в целом были довольно грубыми для такого мелкого животного, причём каждая отдельная нить намного больше и толще, чем волоски млекопитающих такого же размера[25].
Длина волокон различна. В типовом образце самые короткие волокна расположены перед глазами — их длина составляет 13 миллиметров. При продвижении дальше по телу длина волокон быстро увеличиваются до достижения максимального размера в 35 миллиметров над лопатками. Длина остаётся одинаковой до середины спины, вплоть до бёдер, где нити снова удлиняются и достигают максимальной длины в 40 мм в середине хвоста. Волокна на нижней стороне хвоста в целом короче, чем на верхней стороне, и их длина уменьшается быстрее. К 25 хвостовому позвонку нити на нижней стороне достигают длины всего 35 мм. Самые длинные волокна, располагавшиеся на предплечьях, достигают 14 миллиметров[25].
Хотя волокна расположены слишком плотно, чтобы можно было изолировать одно из них для изучения, в некоторых исследованиях предполагают наличие двух разных типов нитей (толстых и тонких), чередующихся друг с другом. Толстые нити выглядят более жёсткими, чем тонкие, при этом тонкие волокна имеют тенденцию располагаться параллельно друг к другу, но под углом к толстым нитям. Такие свойства волокон предполагают наличие у каждого из них стержня с бородками, похожими на таковые в перьях современных птиц, но более примитивные[14]:1721. В целом, эти структуры напоминают пуховые перья некоторых современных птиц; те же структуры были найдены и в других ископаемых исяньской формации, включая конфуциусорниса[25].
Однако, проведённое в 2018 году исследование показало, что толстые нити могут быть пучками переплетённых тонких нитей. Такая гипотеза подтверждается наблюдением, что тонкие нити имеют тенденцию к выравниванию параллельно друг другу и толстым волокнам, а не к разветвлению, как указывали другие учёные. Некоторые из толстых нитей довольно длинные и заканчиваются небольшими пучками тонких нитей. Пухообразное оперение обычно выглядит наоборот: короткий центральный стержень и длинные пучки волокон. Кроме того, толстые нити не содержат следов фосфата кальция — минерала, из которого состоят перья современных птиц. Большое число изгибов также делает маловероятным наличие крепкого стержня. Таким образом, гипотеза, что толстые нити представляют собой просто пучки тонких нитей, является более правдоподобной, чем идея о том, что они представляют собой вариант пуховых перьев, которые развили морфологию, противоположную морфологии перьев птиц и пернатых теропод. В целом, авторы работы предпочли гипотезу о том, что перья синозавроптерикса были простыми одинарными нитями, хотя и предположили, что иногда эти нити соединялись у основания в пучки как предшественники настоящих пуховых перьев[28].
Несмотря на то, что синозавроптерикс имел перьеподобные структуры, он не состоял в близком родстве с более древним археоптериксом[25]. Существовало множество других групп динозавров, которые были более тесно связаны с археоптериксом, чем синозавроптерикс, включая дейнонихозавров, овирапторозавров или теризинозавроидов. Это указывает на то, что перья, возможно, были характерной чертой анатомии для многих тероподовых динозавров, а не только для явно похожих на птиц, и весьма вероятно, что у таких не родственных птицам животных, как компсогнатиды, были перья[29].
Расцветка

Синозавроптерикс стал первым динозавром, чью прижизненную расцветку исследователи описали на основе физических доказательств. Некоторые окаменелости этого теропода показывают чередование светлых и тёмных полос, сохранившихся на хвосте. Первоначально Чен и её коллеги интерпретировали эту полосатую структуру как артефакт, образовавшийся при расколе куска породы на плиту и контрплиту, между которыми находилась окаменелость[25]. Однако, в своей презентации 2002 года, подготовленной для Общества палеонтологии позвоночных, британский палеонтолог Ник Лонгрич выдвинул гипотезу, что экземпляры синозавроптерикса фактически сохраняют остатки расцветки, которую эти животные демонстрировали при жизни. Учёный утверждал, что тёмные полоски на хвосте были слишком равномерно распределены, чтобы быть остатками от раскола плиты, и что они представляют собой окаменелый пигмент, ранее присутствовавший в перьях. Кроме того, присутствие тёмных волокон вдоль верхней части тела динозавра также может отражать такой цветовой паттерн защитной окраски, при котором спина синозавроптерикса была тёмного цвета, а брюхо — более светлого, с полосками или пятнами на хвосте[30].
Выводы Лонгрича были поддержаны в научной статье, впервые опубликованной онлайн в журнале «Nature» в январе 2010 года. Фучэн Чжан и его коллеги исследовали окаменевшие перья нескольких динозавров и ранних птиц и обнаружили доказательства того, что они сохраняют меланосомы — клетки, которые придают перьям современных птиц их цвет. Среди исследованных образцов был ранее не описанный экземпляр синозавроптерикса под инвентарным номером IVPP V14202. Изучив структуру и расположение меланосом, Чжан и его коллеги смогли подтвердить наличие светлых и тёмных полос в перьях хвоста динозавра. Кроме того, команда учёных сделала сравнение найденных меланосом с меланосомами современных птиц, чтобы определить основной цвет оперения. По присутствию сферических меланосом, которые содержат красный пигмент, они пришли к выводу, что более тёмные перья синозавроптерикса были каштанового или красновато-коричневого цвета[21]. Более подробное исследование окраски динозавра показало, что у него была «бандитская маска», как у енота, и двусторонний паттерн окраски тела, указывающий на открытую среду обитания, что говорит о том, что в биоте Жэхэ существовало сразу несколько сред обитания[31].
Систематика


Существует только один названный вид синозавроптерикса — Sinosauropteryx prima. Возможный второй вид представлен образцом GMV 2124 (NGMC 2124), который был описан в качестве третьего, более крупного экземпляра этого динозавра[9]. Тем не менее, в 2002 году, во время презентации для Общества палеонтологии позвоночных, Ник Лонгрич продемонстрировал, что данный образец отличается в нескольких анатомических аспектах от других образцов, например, размером тела, пропорционально длинными голенями и более коротким хвостом. Лонгрич предположил, что образец GMV 2124 принадлежал целурозавру семейства компсогнатид, в то время как собственно синозавроптерикс являлся более примитивным целурозавром или даже базальным карнозавром[30]. В 2007 году А. Гишлик и Ж. Готье согласились, что экземпляр мог быть новым таксоном, и предварительно переклассифицировали его как Sinosauropteryx? sp., при этом предположив, что он мог представлять собой новый род[32]. В том же 2007 году С. Цзи и его коллеги также заключили, что образец GMV 2124 принадлежит новому роду, основываясь на длине хвоста и пропорциях задних конечностей[7].
Приведённая ниже кладограмма показывает филогенетическое размещение синозавроптерикса в пределах группы целурозавров, согласно работе 2012 года, выполненной Филом Сентером и его командой[33].
| Coelurosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Отличительные признаки
В 1996 году авторы первоописания перечислили множество признаков, позволяющих отличить синозавроптерикса от других динозавров и птиц. Они описали его как небольшую примитивную птицу с относительно высоким черепом, тупым рострумом и слегка высокой премаксиллой. Предглазничное окно имеет форму овала, но не увеличено, супрангулярная кость узкая и удлинённая, нижнечелюстная кость крепкая, а зубы — острые и чрезвычайно хорошо развиты. Хвост насчитывает более 50 хвостовых позвонков, составляющих больше 60 % длины тела. Передние конечности чрезвычайно маленькие, с короткими и толстыми плечевыми костями. Лобковая кость удлинена, её дистальный конец чрезвычайно завышен, а таз широк. Задние конечности длинные и крепкие, большеберцовая кость немного длиннее бедренной кости, предплюсна не слита, а кости плюсны относительно крепкие, с не слитыми проксимальными концами. Перья короткие, маленькие и однородные, большое их количество украшает вершину черепа, шейный и спинной отделы, а также верхнюю и нижнюю стороны хвоста[6].
Палеобиология
Рацион

Образец NIGP 127587 сохранился с остатками ящерицы в области пищеварительного тракта, что указывает на то, что мелкие проворные позвоночные составляли часть рациона синозавроптерикса. Множество ящериц этого типа было обнаружено в тех же породах, что и S. prima[25]. С большой долей вероятности эти рептилии принадлежали к роду Dalinghosaurus — быстро бегающим ящерицам, приспособленным к жизни на открытых пространствах, как и сам синозавроптерикс[31].
Образец, возможно принадлежавший синозавроптериксу (GMV 2124, Sinosauropteryx? sp.) содержит три челюстные кости млекопитающих в районе пищеварительной системы. В 2006 году палеонтологи идентифицировали две из этих челюстей как принадлежащие симметродонту Zhangheotherium, а третью — принадлежащую животному из отряда многобугорчатых Sinobaatar, указав, что два этих рода млекопитающих были частью рациона динозавра. Известно, что на лодыжках у Zhangheotherium были шпоры, как у современного утконоса; таким образом, синозавроптерикс, возможно, питался ядовитыми млекопитающими[34].
Размножение
Тот же экземпляр S. prima, который сохранил остатки ящерицы среди содержимого своего желудка (NIGP 127587), содержал несколько крошечных яиц в животе. Два яйца сохранились прямо перед «ботинком» лобковой кости, а ещё несколько располагались под динозавром, на плите. Маловероятно, чтобы яйца были съедены животным, поскольку они находятся не в той полости тела, где расположен пищеварительный аппарат, а также потому, что скорлупа яиц осталась нетронутой. Более вероятно, это неотложенные яйца самого синозавроптерикса. Каждое яйцо имеет длину 36 миллиметров и ширину 26 миллиметров. Присутствие двух развитых яиц предполагает наличие у синозавроптерикса парных яйцеводов и откладывание яиц парами, как у других теропод[25].
Палеоэкология
Синозавроптерикс, как динозавр, найденный в формации Исянь, являлся представителем биоты Жэхэ — совокупности организмов, обнаруженных в формации Исянь и перекрывающей её формации Цзюфотан[8]. Исяньская формация состоит в основном из вулканических пород, таких как андезит и базальт. Между вулканическими слоями находятся несколько слоёв осадочных пород, представляющих озёрные отложения[25]. В пресноводных озёрных наносах сохранилось множество остатков растений, беспозвоночных и позвоночных животных. Озёра располагались среди обширных лесов голосеменных и ранних цветковых деревьев. Здесь в изобилии водились остракоды и насекомые, двустворчатые моллюски и брюхоногие. Млекопитающие и птицы также хорошо известны из этих формаций. Эта местность подвергалась периодическим катастрофам, которые заканчивались массовой гибелью животных: извержениям вулканов, лесным пожарам и выбросу ядовитых газов, выходящих из озёр[8]. Климат был интерпретирован как умеренный, с выраженными засушливыми и дождливыми сезонами[35]. Средняя годовая температура равнялась примерно 10 °C, что указывает на умеренный климат с непривычно холодными зимами для в целом тёплой мезозойской эры, возможно, из-за расположения этой территории в более высоких широтах, чем сейчас[36].
Примечания
- Sinosauropteryx prima (англ.) информация на сайте Paleobiology Database. (Дата обращения: 2 декабря 2018).
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 76, 104. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Нэйш Д., Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. Александр Аверьянов. — М.: Альпина нон-фикшн, 2019. — С. 120, 139, 187. — 223 с. — ISBN 978-5-91671-940-6.
- Browne M. W. Feathery Fossil Hints Dinosaur-Bird Link. New York Times (19 October 1996). Дата обращения: 13 ноября 2018.
- Feduccia A. The Origin and Evolution of Birds (Second ed.). — Yale University Press, 1999. — P. 375. — ISBN 978-0-300-07861-9.
- Ji Q., Ji S. On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds (англ.) // Chinese Geology. — 1996. — Vol. 10, no. 233. — P. 30—33.
- Ji S., Gao C., Liu J., Meng Q., Ji Q. New material of Sinosauropteryx (Theropoda: Compsognathidae) from western Liaoning, China (англ.) // Acta Geologica Sinica (English Edition). — 2007. — Vol. 81, no. 2. — P. 177—182. — doi:10.1111/j.1755-6724.2007.tb00942.x.
- Zhou Z. Evolutionary radiation of the Jehol Biota: chronological and ecological perspectives (англ.) // Geological Journal. — 2006. — Vol. 41, no. 3—4. — P. 377—393. — doi:10.1002/gj.1045.
- Ji Q., Ji S. Advances in Sinosauropteryx research (англ.) // Chinese Geology. — 1997. — Vol. 7. — P. 30—32.
- Rauhut O. W. M. The interrelationships and evolution of basal theropod dinosaurs (англ.) // Special Papers in Palaeontology. — 2003. — Vol. 69. — P. 1—213.
- Senter P. Phylogeny of Drepanosauridae (Reptilia: Diapsida) (англ.) // Journal of Systematic Palaeontology. — 2004. — Vol. 2, no. 3. — P. 257—268. — doi:10.1017/S1477201904001427.
- Senter P., Barsbold R., Britt B. B., Burnham D. A. Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda) (англ.) // Bulletin of the Gunma Museum of Natural History. — 2004. — Vol. 8. — P. 1—20.
- Rauhut O. W. M., Milner A. C., Moore-Fay S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England (англ.) // Zoological Journal of the Linnean Society. — 2010. — Vol. 158, no. 1. — P. 155—195. — doi:10.1111/j.1096-3642.2009.00591.x.
- Currie P. J., Chen P.-j. Anatomy of Sinosauropteryx prima from Liaoning, northeastern China (англ.) // Canadian Journal of Earth Sciences. — 2011. — February (vol. 38, no. 12). — P. 1705—1727. — doi:10.1139/cjes-38-12-1705.
- Stieg B. Debate rages over birds' relation to dinosaurs (англ.) // Knight-Ridder Newspapers. — 1997. — 23 April.
- Morell V. The origin of birds: the dinosaur debate (англ.) // Audubon Magazine. — 1997. — March/April. — P. 36—45.
- Martin L., Czerkas S. A. The fossil record of feather evolution in the Mesozoic (англ.) // American Zoologist. — 2000. — Vol. 40, no. 4. — P. 687—694. — doi:10.1668/0003-1569(2000)040[0687:TFROFE]2.0.CO;2.
- Ruben J. A., Jones T. D. Selective factors associated with the origin of fur and feathers (англ.) // American Zoologist. — 2000. — Vol. 40, no. 4. — P. 585—596. — doi:10.1093/icb/40.4.585.
- Lingham-Soliar T., Feduccia A., Wang X. A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres (англ.) // Proceedings of the Royal Society B. — 2007. — Vol. 274, no. 1620. — P. 1823—1829. — doi:10.1098/rspb.2007.0352.
- Sanderson K. Bald dino casts doubt on feather theory (англ.) // Nature. — 2007. — 23 May. — doi:10.1038/news070521-6.
- Zhang F., Kearns S. L., Orr P. J., Benton M. J., Zhou Z., Johnson D., Xu X., Wang X. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds (англ.) // Nature. — 2010. — Vol. 463, no. 7284. — P. 1075—1078. — doi:10.1038/nature08740.
- Sloan C. Dinosaur True Colors Revealed for First Time. National Geographic (27 January 2010). Дата обращения: 14 ноября 2018.
- Paul G. S. Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. — Baltimore, Maryland: Johns Hopkins University Press, 2002. — P. 66. — 472 p. — ISBN 978-0-8018-6763-7.
- Smithwick F. M., Mayr G., Saitta E. T., Benton M. J., Vinther J. On the purported presence of fossilized collagen fibres in an ichthyosaur and a theropod dinosaur (англ.) // Palaeontology. — 2017. — Vol. 60, no. 3. — P. 409—422. — doi:10.1111/pala.12292.
- Chen P., Dong Z., Zhen S. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China (англ.) // Nature. — 1998. — Vol. 391, no. 8. — P. 147—152. — doi:10.1038/34356.
- Therrien F., Henderson D. M. My theropod is bigger than yours... or not: estimating body size from skull length in theropods (англ.) // Journal of Vertebrate Paleontology. — 2007. — Vol. 27, no. 1. — P. 108—115. — doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- Ruben J. A., Jones T. D., Geist N. R., Hillenius W. J. Lung structure and ventilation in theropod dinosaurs and early birds (англ.) // Science. — 1997. — Vol. 278, no. 5341. — P. 1267—1270. — doi:10.1126/science.278.5341.1267.
- Saitta E. T., Gelernte R., Vinther J. Additional information on the primitive contour and wing feathering of paravian dinosaurs (англ.) // Palaeontology. — 2018. — Vol. 61, no. 2. — P. 273—288. — doi:10.1111/pala.12342.
- Padian K. Basal Avialae // The Dinosauria (Second ed.) / Weishampel D. B., Peter Dodson, Osmólska H.. — Berkeley: University of California Press, 2004. — ISBN 978-0-520-24209-8.
- Longrich N. Systematics of Sinosauropteryx (англ.) // Journal of Vertebrate Paleontology. — 2002. — Vol. 22 (supplement to 3). — P. 80A.
- Smithwick F.M., Nicholls R., Cuthill I. C., Vinther J. Countershading and Stripes in the Theropod Dinosaur Sinosauropteryx Reveal Heterogeneous Habitats in the Early Cretaceous Jehol Biota (англ.) // Current Biology. — 2017. — Vol. 27, no. 21. — P. 3337—3343.e2. — doi:10.1016/j.cub.2017.09.032.
- Gishlick A. D., Gauthier J. A. On the manual morphology of Compsognathus longipes and its bearing on the diagnosis of Compsognathidae (англ.) // Zoological Journal of the Linnean Society. — 2007. — Vol. 149, no. 4. — P. 569—581. — doi:10.1111/j.1096-3642.2007.00269.x.
- Senter P., Kirkland J. I., Deblieux D. D., Madsen S., Toth N. New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail (англ.) // PLoS ONE. — 2012. — Vol. 7, no. 5. — P. e36790. — doi:10.1371/journal.pone.0036790.
- Hurum J. H., Luo Z.-X., Kielan-Jaworowska Z. Were mammals originally venomous? (англ.) // Acta Palaeontologica Polonica. — 2006. — Vol. 51, no. 1. — P. 1—11.
- Wang Y., Zheng S., Yang X., Zhang W., and Ni Q. The biodiversity and palaeoclimate of conifer floras from the Early Cretaceous deposits in western Liaoning, northeast China (англ.) // International Symposium on Cretaceous Major Geological Events and Earth System. — 2006. — P. 56A.
- Amiot R., Wang X., Zhou Z., et al. Oxygen isotopes of East Asian dinosaurs reveal exceptionally cold Early Cretaceous climates (англ.) // Proceedings of the National Academy of Sciences. — 2011. — Vol. 108, no. 13. — P. 5179—5183. — doi:10.1073/pnas.1011369108.
Ссылки
- Новые динозавры из Китая и происхождение птиц. Природа Земли. Дата обращения: 15 ноября 2018.