Тетануры
Тетануры[2] (лат. Tetanurae) — клада динозавров-теропод в составе группы аверостр, включающая птиц и все таксоны, более родственные им, чем цератозаврам[3][4]. Все тетануры, за исключением базальных родов, входят в состав клады Orionides[5]. В зависимости от того, какая топология принимается за верную, клада Orionides может подразделяться на карнозавров (Carnosauria) и целурозавров (Coelurosauria)[6], либо на мегалозавроид (Megalosauroidea) и аветеропод[7] (Avetheropoda), к последним из которых, в свою очередь, относят целурозавров и аллозавроид[8] (Allosauroidea)[9][10].
| Тетануры | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: аллозавр, спинозавр, торвозавр; | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Клада: Тетануры |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Tetanurae Gauthier, 1986 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
по The Theropod Database[1]:
|
|||||||||||||||||||||||||||||||||||||||||
| Дочерние таксоны | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 190 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Первые тетануры появляются в палеонтологической летописи в отложениях начала средней юры, возрастом около 190 млн лет, где представлены только скудными остатками[5]. Сама группа могла возникнуть несколько раньше — в конце ранней юры[6]. В средней юре тетануры распространились повсеместно. В ходе эволюции среди тетанур неоднократно независимо возникали крупные таксоны (закон Копа), в том числе самые большие тероподы. Линия, которая привела к птицам, в данном случае является исключением, поскольку её представители постоянно уменьшались в размерах[5][11].
Характеристика
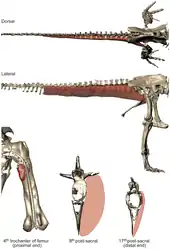
Клада Tetanurae была формально выделена Жаком Готье в 1986 году. Название таксона буквально переводится как «жёсткие хвосты» и относится к строению точки перехода, то есть области хвоста, где заканчивались боковые отростки и, вместе с ними, как предполагается, хвостово-бедренная мышца[4]. Исходя из расположения отростков, у нептичьих тетанур имелась самая длинная среди теропод хвостово-бедренная мышца (лат. musculus caudofemoralis), которая поддерживала хвост и ограничивала его гибкость. Увеличение хвостово-бедренной мышцы стало результатом тенденции к сокращению важности хвоста как части локомоторного модуля теропод, что имело значительные последствия для их локомоции и, в конечном итоге, эволюции птичьего полёта[12].
Готье (1986) выделяет следующие 17 синапоморфий, имевшихся у первых тетанур и унаследованных многими из их потомков[4]:
- отсутствие клыковидных зубов в нижней челюсти (не определимо у беззубых таксонов);
- верхнечелюстное отверстие большое, расположено кзади (у птицехвостых отсутствует);
- анторбитальный зубной ряд (нет у беззубых таксонов; конвергенция с другими хищным архозавроморфами);
- расширенный на конце остистый отросток эпистрофея;
- точка перехода начинается ближе к проксимальной половине хвоста (нет у птиц, лишённых пигостиля);
- лопатка узкая, но расширенная на концах (обычно симметрично, но у тираннозавра расширение есть только с дистальной стороны);
- коракоид удлинённый и заострённый кзади;
- кисть достигает не менее 2/3 длины плеча вместе с предплечьем;
- базальная половина пястной кости I прижата к пястной кости II;
- основание пястной кости III на ладонной поверхности кисти расположено вентрально по отношению к остальной части кости;
- четвёртый палец кисти отсутствует после прохождения эмбриональной стадии (сообщалось возможное наличие его зачатков у эмбрионов птиц);
- на седалищной кости имеется обтураторный отросток (возможный гомолог нижнего отростка седалищной кости птиц);
- на лонном сочленении имеется дистальный «башмак»;
- бедренная кость с крыловидным малым трохантером;
- восходящий отросток таранной кости высокий и широкий, поверхностно расположенный;
- плюсневые кости II и IV принимают значительное участие в формировании голеностопного сустава, а плюсневая кость III зажата между ними в разной степени;
- плюсневая кость I относительно короткая.
Позже также были отмечены пневматизация черепа, относительная редукция малоберцовой кости в сравнении с большеберцовой и наличие горизонтального желобка на мыщелке таранной кости[13][12].
Систематика


По первоначальному филогенетическому определению Готье (1986), к тетанурам относятся современные птицы (Aves) и все таксоны, более родственные им, чем цератозаврам (Ceratosauria)[4]. Теперь обычно используется более формальная версия этого определения, по которой тетануры — клада, включающая все таксоны, ближе родственные домовому воробью (Passer domesticus), чем цератозавру Ceratosaurus nasicornis[3][1][14]. Таким образом, тетануры и цератозавры являются сестринскими таксонами и вместе взятые образуют кладу аверостр (Averostra sensu Ezcurra & Cuny, 2007[15])[9], или Neotheropoda sensu Bakker, 1986[16][17].
Все тетануры, кроме базальных родов, относятся к кладе Orionides Carrano et al., 2012. М. Т. Каррано и коллеги (2012), определили её следующим образом: наименьшая клада, включающая мегалозавра Megalosaurus bucklandii (мегалозавроид), аллозавра Allosaurus fragilis (аллозавроид) и домового воробья (целурозавр)[5]. К числу базальных тетанур, не входящих в состав Orionides, могут относиться Chuandongocoelurus, Pandoravenator и Monolophosaurus[1].
Готье рассматривал в составе тетанур две основные подгруппы: карнозавры (Carnosauria) и целурозавры (Coelurosauria)[4]. Карнозавры в традиционном понимании являются полифилетической группой, в которую включали любых крупных теропод[5]. Схожей точки зрения придерживался Готье, включая в их состав тираннозавроид[4], которые теперь считаются целурозаврами, независимо достигшими гигантских размеров[9].
В настоящее время сторонники монофилии карнозавров помимо отдельных родов включают в их состав только спинозаврид (Spinosauridae), мегалозаврид[8] (Megalosauridae) и аллозавроид (первые два таксона могут объединяться в группу мегалозавроид, по Rauhut & Pol, 2019 являющуюся парафилетической), а также неустоявшегося положения Piatnitzkysauridae (мегалозавроиды[9][10], либо аллозавроиды[6]). Многие авторы, однако, полагают, что аллозавроиды более родственны целурозаврам, чем мегалозавроидам; в таком случае выделение карнозавров оказывается не имеющим практического смысла[9]. Аветероподы (Avetheropoda Paul, 1988[18]), или Neotetanurae Sereno et al., 1994[19] (первое название имеет приоритет), определяются как наименьшая клада, включающая аллозавра Allosaurus fragilis (аллозавроид) и домового воробья (целурозавр). Выделение аветеропод оправдано только если аллозавроиды более близки к целурозаврам, чем к мегалозавроидам[20].
Филогения
Филогенетическое положение тетанур может быть отражено следующей упрощённой кладограммой[9][6][21]:
| Averostra |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||


Спорное положение в пределах группы тетанур занимает клада Megaraptora, в разных исследованиях относимая к аллозавроидам семейства Neovenatoridae[22][5][10], к тираннозавроидам[23][24] или к базальным целурозаврам[25][26].
| Zanno & Makovicky, 2013[10] | Rauhut & Pol, 2019[6] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Примечания
- Mortimer M. Tetanurae (англ.). The Theropod Database. Дата обращения: 24 мая 2021.
- Татаринов, 2009, с. 71.
- Weishampel, Dodson & Osmólska, 2004, chpt. 4: "Basal Tetanurae" by T. R. Holtz Jr., R. E. Molnar and P. J. Currie, pp. 71—110.
- In Padian K. (ed.), 1986, "Saurischian monophyly and the origin of birds" by J. A. Gauthier, pp. 1—55.
- Carrano M. T., Benson R. B. J., Sampson S. D. The phylogeny of Tetanurae (Dinosauria: Theropoda) (англ.) // Journal of Systematic Palaeontology : journal. — 2012. — Vol. 10, iss. 2. — P. 211—300. — ISSN 1477-2019. — doi:10.1080/14772019.2011.630927.
- Rauhut O. W. M., Pol D. Probable basal allosauroid from the early Middle Jurassic Canadon Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs (англ.) // Scientific Reports : journal. — 2019. — Vol. 9. — ISSN 2045-2322. — doi:10.1038/s41598-019-53672-7.
- Татаринов, 2009, с. 73.
- Нэйш и Барретт, 2019, «Мегалозавриды, спинозавры и аллозавриды», с. 47—50.
- Hendrickx C., Hartman S. A., Mateus O. An Overview of Non-Avian Theropod Discoveries and Classification (англ.) // PalArch’s Journal of Vertebrate Palaeontology : journal. — 2015. — Vol. 12, iss. 1. — P. 1—73.
- Zanno L. E., Makovicky P. J. Neovenatorid theropods are apex predators in the Late Cretaceous of North America (англ.) // Nature Communications : journal. — 2013. — Vol. 4, iss. 1. — P. 2827. — ISSN 2041-1723. — doi:10.1038/ncomms3827.
- Hone D. W. E., Keesey T. M., Pisani D., Purvis A. Macroevolutionary trends in the Dinosauria: Cope's rule (англ.) // Journal of Evolutionary Biology : journal. — 2005. — Vol. 18, iss. 3. — P. 587—595. — ISSN 1420-9101. — doi:10.1111/j.1420-9101.2004.00870.x.
- Currie & Padian, 1997, "Tetanurae" by J. R. Hutchinson, pp. 727—728.
- Татаринов, 2009, с. 72.
- Sereno P. C. Tetanurae (англ.). TaxonSearch. Дата обращения: 24 мая 2021.
- Ezcurra M. D., Cuny G. The coelophysoid Lophostropheus airelensis, gen. nov.: A review of the systematics of "Liliensternus" airelensis from the Triassic-Jurassic outcrops of Normandy (France) (англ.) // Journal of Vertebrate Paleontology : journal. — 200. — Vol. 27, iss. 1. — P. 73—86. — ISSN 0272-4634. — doi:10.1671/0272-4634(2007)27[73:TCLAGN]2.0.CO;2.
- Bakker, 1986, pp. 460—461.
- Mortimer M. Neotheropoda (англ.). The Theropod Database. Дата обращения: 24 мая 2021.
- Paul, 1988, "Birdlike Avetheropods", p. 296.
- Sereno P. C., Wilson J. A., Larsson H. C. E., Dutheil D. B., Sues H.-D. Early Cretaceous Dinosaurs from the Sahara (англ.) // Science : journal. — 1994. — Vol. 266, iss. 5183. — P. 267—271. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.266.5183.267.
- Mortimer M. Avetheropoda (англ.). The Theropod Database. Дата обращения: 24 мая 2021.
- Hartman S., Mortimer M., Wahl W. R., Lomax D. R., Lippincott J. Supplemental Information: A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight (англ.) // PeerJ : journal. — 2019. — Vol. 7. — P. e7247. — ISSN 2167-8359. — doi:10.7717/peerj.7247.
- Benson R. B. J., Carrano M. T., Brusatte S. L. A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic (англ.) // Naturwissenschaften : journal. — 2010. — Vol. 97, iss. 1. — P. 71—78. — ISSN 1432-1904. — doi:10.1007/s00114-009-0614-x. — . — PMID 19826771.
- Novas F. E., Agnolín F. L., Ezcurra M. D., Canale J. I., Porfiri J. D. Megaraptorans as members of an unexpected evolutionary radiation of tyrant-reptiles in Gondwana (англ.) // Ameghiniana : journal. — 2012. — Vol. 49 (Suppl), no. 4. — P. R33.
- Porfiri J. D., Novas F. E., Calvo J. O., Agnolín F. L., Ezcurra M. D., Cerda I. A. Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation (англ.) // Cretaceous Research : journal. — 2014. — Vol. 51. — P. 35—55. — ISSN 0195-6671. — doi:10.1016/j.cretres.2014.04.007.
- Porfiri J. D., Juárez Valieri R. D., Santos D. D. D., Lamanna M. C. A new megaraptoran theropod dinosaur from the Upper Cretaceous Bajo de la Carpa Formation of northwestern Patagonia (англ.) // Cretaceous Research : journal. — 2018. — Vol. 89. — P. 302—319. — ISSN 0195-6671. — doi:10.1016/j.cretres.2018.03.014.
- Delcourt R., Grillo O. N. Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology : journal. — 2018. — Vol. 511. — P. 379—387. — ISSN 0031-0182. — doi:10.1016/j.palaeo.2018.09.003. — .
Литература
На русском языке
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 71—114. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Нэйш Д., Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. А. О. Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2019. — 223 с. — ISBN 978-5-91671-940-6.
На английском языке
- Encyclopedia of Dinosaurs (англ.) / ed. by Currie P. J., Padian K.. — Academic Press, 1997. — 869 p. — ISBN 978-0-12-116810-6.
- The Origin of Birds and the Evolution of Flight (англ.) / ed. by K. Padian. — San Francisco, CA: California Academy of Sciences, 1986. — Vol. 8. — 98 p. — (Memoirs of the California Academy of Sciences).
- The Dinosauria (англ.) / D. B. Weishampel, P. Dodson, H. Osmólska, eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p. — ISBN 978-0-520-25408-4.
- Paul G. S. Predatory Dinosaurs of the World: A Complete Illustrated Guide (англ.). — New York: Simon & Schuster, 1988. — 464 p. — ISBN 978-0-671-61946-6.
- Bakker R. T. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction (англ.). — New York: William Morrow and Company, 1986. — 481 p. — ISBN 978-0-688-04287-5.
| Таксономия |
|---|