Сахаромицеты
Сахаромице́ты[1] (лат. Saccharomycetes) — класс сумчатых грибов (аскомицетов). Это — единственный класс подотдела сахаромико́товые[2] (лат. Saccharomycotina). Класс содержит единственный порядок сахаромице́товые[1] (лат. Saccharomycetales); порядок включает 13 семейств и 99 родов[3].
| Сахаромицеты | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
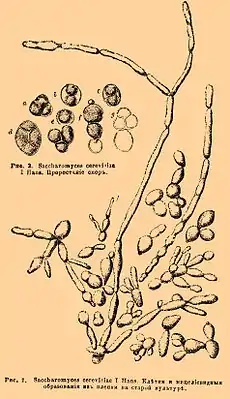 Saccharomyces cerevisiae. Иллюстрация из Энциклопедического словаря Брокгауза и Ефрона | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Отдел: Подотдел: Saccharomycotina O.E.Erikss. & Winka, 1997 Класс: Сахаромицеты |
||||||||||||
| Международное научное название | ||||||||||||
| Saccharomycetes O.E. Erikss. & Winka, 1997 | ||||||||||||
| Синонимы | ||||||||||||
| Дочерние таксоны | ||||||||||||
|
||||||||||||
| ||||||||||||
Иногда русское название «сахаромицеты» употребляют одновременно как для всего подотдела Saccharomycotina, так и для класса Saccharomycetes[1].
Систематика
Вплоть до конца XX века относимые ныне к сахаромицетам грибы включали в состав подкласса Hemiascomycetidae (гемиаскомицеты, или голосумчатые) класса Ascomycetes, объединяя в данном подклассе входившие к группе голосумчатых относили аскомицеты примитивного строения, у которых не образуются плодовые тела, а сумки развиваются либо непосредственно на мицелии, либо (у дрожжевых форм) как одиночные свободные клетки. Данный подкласс разделяли на 4 порядка[4]:
- Эндомицетовые (Endomycetales);
- Тафриновые (Taphrinales);
- Протомицетовые (Protomycetales);
- Аскосферовые (Ascosphaerales).
Впоследствии ранг гемиаскомицетов был поднят до класса, причём из него был выделен самостоятельный класс Archiascomycetes (археаскомицеты; в настоящее время — подотдел Taphrinomycotina[5]), к которому отнесли тафриновых и протомицетовых, а также семейство схизосахаромицетовые (Schizosaccharomycetaceae), включающее делящиеся дрожжи[6]. Аскосферовые перенесли в класс эуроциомицеты (Eurotiomycetes)[7]. Название «гемиаскомицеты» теперь стали применять к подотделу Hemiascomycotina, но вскоре последнее заменили типифицированным названием Saccharomycotina[1]. К 2009 году в составе подотдела имелся единственный класс Saccharomycetes с единственным порядком Saccharomycetales, включавшим 12 семейств (Ascoideaceae, Cephaloascaceae, Dipodascaceae, Endomycetaceae, Eremotheciaceae, Lipomycetaceae, Metschnikowiaceae, Pichiaceae, Saccharomycetaceae, Saccharomycodaceae, Saccharomycopsidaceae, Trichomonascaceae), а также 9 неклассифицированных родов[7].
На конец 2017 года в порядке Saccharomycetales выделяли 13 семейств и 12 неклассифицированных родов[3]:
- Lipomycetaceae: Dipodascopsis, Kockiozyma, Limtongia, Lipomyces, Myxozyma;
- Trigonopsidaceae: Botryozyma, Tortispora, Trigonopsis;
- Dipodascaceae: Blastobotrys, Dipodascus, Galactomyces, Geotrichum, Magnusiomyces, Saprochaete;
- Trichomonascaceae: Deakozyma, Diddensiella, Groenewaldozyma, Middelhovenomyces, Nadsonia, Spencermartinsiella, Starmerella, Sugiyamaella, Wickerhamiella, Yarrowia, Zygoascus;
- Pichiaceae: Brettanomyces, Dekkera, Hyphopichia, Komagataella, Kregervanrija, Kuraishia, Martiniozyma, Nakazawaea, Ogataea, Pachysolen, Peterozyma, Pichia, Saturnispora;
- Cephaloascaceae: Cephaloascus;
- Metschnikowiaceae: Clavispora, Danielozyma, Kodamaea, Metschnikowia, Wickerhamia;
- Debaryomycetaceae: Babjeviella, Debaryomyces, Hemisphaericaspora, Kurtzmaniella, Lodderomyces, Meyerozyma, Millerozyma, Priceomyces, Scheffersomyces, Schwanniomyces, Spathaspora, Suhomyces, Teunomyces, Yamadazyma;
- Ascoideaceae: Ascoidea;
- Saccharomycopsidaceae: Ambrosiozyma, Saccharomycopsis;
- Phaffomycetaceae: Barnettozyma, Cyberlindnera, Phaffomyces, Starmera, Wickerhamomyces;
- Saccharomycodaceae: Hanseniaspora, Kloeckera, Saccharomycodes;
- Saccharomycetaceae: Citeromyces, Cyniclomyces, Eremothecium, Hagleromyces, Kazachstania, Kluyveromyces, Lachancea, Metahyphopichia, Nakaseomyces, Naumovozyma, Saccharomyces, Tetrapisispora, Torulaspora, Vanderwaltozyma, Yueomyces, Zygosaccharomyces, Zygotorulaspora;
- incertae sedis: Aciculoconidium, Candida, Coccidiascus, Conidiascus, Diutina, Endomyces, Macrorhabdus, Oosporidium, Oscarbrefeldia, Phialoascus, Selenozyma, Sporopachydermia.
Семейства Endomycetaceae и Eremotheciaceae более не выделяются. Входившие в первое из них роды Endomyces и Phialoascus отнесены к группе родов неясного систематического положения; к ней же отнесён и входивший в семейство Eremotheciaceae род Coccidiascus, а второй род данного семейства — Eremothecium — включён в семейство Saccharomycetaceae[3]. Среди родов incertae sedis особенно следует отметить род Candida — сборный таксон, многочисленные виды которого, согласно данным молекулярной филогенетики, относятся к самым различным семействам сахаромицетов[8][9].
Филогения
В соответствии с результатами молекулярно-филогенетических исследований, филогенетические связи между отдельными семействами сахаромицетов можно отобразить при помощи следующей кладограммы[8][10][11]:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lipomycetaceae | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Описание
Вегетативное тело сахаромицетов часто представляет собой отдельные почкующиеся клетки, то есть дрожжевую форму, в определённых условиях они могут образовывать псевдомицелий, есть и виды, имеющие настоящий мицелий. Некоторые виды диморфны — в зависимости от условий развиваются в мицелиальной или дрожжевой форме. К мицелиальным сахаромицетам относится, например, семейство Диподасковые (Dipodascaceae), виды которого встречаются в слизистых истечениях растений, на древесине, в почве, в ассоциациях с беспозвоночными животными (насекомые, кольчатые черви).
Размножение — вегетативным, бесполым и половым способом, но вегетативное размножение (почкованием или фрагментацией гиф) у сахаромицетов часто трудно чётко отграничить от бесполого (образования конидий).
Половой процесс — копуляция вегетативных клеток, при которой после слияния плазмы (стадии плазмогамии) сразу следует слияние ядер (стадия кариогамии), дикариотическая фаза отсутствует. Аскогенные гифы и плодовые тела не образуются, зигота развивается непосредственно в сумку или образует аскофор — вырост, дающий одну или несколько сумок. У мицелиальных представителей известна и гаметангиогамия — так, у Dipodascus albidus образуются пары многоядерных гаметангиев различных размеров, сливается, однако, только одна пара ядер, а остальные дегенерируют. У других видов (Eremascus fertilis, Dipodascus aggregatus) известны гаметангии в виде одноядерных клеток или отростков клеток, содержащих по одному ядру. У вида Endomycopsis vernalis, встречающегося на истечениях сока из деревьев, наблюдается апомиксис — сумки развиваются не из зиготы, а из вегетативных клеток.
Сумки тонкостенные, выход спор осуществляется в результате разрыва или лизиса оболочек сумок.
Смена ядерных фаз в жизненном цикле у разных представителей различна. Например, у Saccharomyces cerevisiae длительное время происходит почкование гаплоидных клеток, после копуляции продолжается почкование в диплоидной фазе, а в благоприятных условиях диплоидные клетки образуют сумки. У видов, являющихся гаплобионтами диплоидизация происходит непосредственно перед формированием сумок, у других же, напротив, гаплоидная фаза представлена только аскоспорами, или даже копуляция происходит между спорами в сумке (Saccharomycodes ludwigii).
Экология
Условия, в которых обитают сахаромицеты бывают очень различными.
Хорошо известны представители, живущие в сахаристых жидкостях — соках деревьев, плодов, некоторые способны развиваться даже в мёде (род Zygosaccharomyces). Такие виды могут вызывать спиртовое брожение сахаров (в анаэробных условиях, то есть при недостаточном доступе воздуха) или их окисление. Некоторые виды дрожжей спиртового брожения известны только в культуре. Все эти грибки сбраживают, в первую очередь, глюкозу, другие моно- и олигосахариды. Существуют так называемые «молочные дрожжи» (род Kluyveromyces), сбраживающие лактозу и участвующие в образовании молочнокислых продуктов. Для всех дрожжей характерен и аэробный (дыхательный) метаболизм, поэтому при доступе воздуха углеводы не сбраживаются до спирта, а окисляются.
Есть сапротрофы, обитающие на поверхности плодов и растительных остатков, в почве. Большинство таких видов являются слабобродящими или вовсе не способными к спиртовому брожению, они обладают преимущественно или только аэробным обменом веществ и окисляют сахара и другие питательные субстраты. Из луговых и болотных почв часто выделяются виды Williopsis saturnus, Lindnera suaveolens и Komagataea pratensis (два последних вида ранее тоже относили к роду Williopsis), в луговых и чернозёмных почвах обитают представители рода Schwanniomyces. Особое значение в почвенной микрофлоре имеют липомицеты, или жировые дрожжи — род Lipomyces. Все виды липомицетов не вызывают брожения, они развиваются в условиях низкого содержания азота в различных типах почв и в разнообразных климатических условиях. Наличие липомицетов приводит к изменению почвенной структуры, а выделяемые ими олигосахариды привлекают азотфиксирующих бактерий.
Есть виды, тесно связанные с насекомыми, так, Dipodascus aggregatus обитает в личиночных ходах жуков-короедов, запах этого гриба привлекает жуков, которые распространяют липкие аскоспоры. Грибки из рода Pichia также известны в ассоциациях с насекомыми-ксилофагами, некоторыми комарами, плодовыми мушками (дрозофилами). У дрозофил дрожжи постоянно живут в кишечнике и при откладке яиц попадают на поверхность плодов. Личинки питаются грибковой биомассой, которая разрастается к моменту выхода их из яиц. Schwanniomyces cantarellii и Schwanniomyces formicarius (оба вида раньше относили к роду Debaryomyces) поселяются в муравейниках рыжего лесного муравья (Formica rufa), в муравьиных гнёздах эти дрожжи могут обнаруживаться в концентрации более 10 миллионов клеток на грамм субстрата. Насекомым дрожжевые грибки необходимы для завершения своего жизненного цикла, некоторые в стерильных условиях, в отсутствие грибка, погибают.
Виды рода Candida встречаются как представители нормальной микрофлоры человеческой кожи и слизистых оболочек, при дисбактериозах могут становиться патогенными, вызывая заболевания различной степени тяжести — кандидозы.
Считается, что галофилов среди сахаромицетов нет, виды, способные размножаться при повышенных концентрациях соли относят к галотолерантным (солеустойчивым). К нии принадлежат дебариомицеты, или «дрожжи де Бари» (род Debaryomyces), некоторые из них могут жить в рассолах, близких по концентрации к насыщенным. Дебариомицеты образуют «плесень» на поверхности сыров, колбасных изделий, солонины, плёнки на поверхности рассолов. Они высеваются также при микозах кожи у человека и животных, однако патогенность их не выяснена.
К паразитам растений относят группу грибков, которую раньше выделяли в семейство спермофторовых (Spermophthoraceae), они развиваются преимущественно на плодах и вызывают повреждения, называемые «стигматомикозами» (представители родов Eremothecium, Nematospora).
Некоторые представители рода Мечниковия (Metschnikowia), или «дрожжей Мечникова», являются паразитами пресноводных и морских беспозвоночных. Наземные представители этого рода обнаруживаются в нектаре цветков и на поверхности различных органов растений, вероятно, часть их жизненного цикла проходит в теле насекомых-переносчиков.
Оптимальная температура жизнедеятельности различна для разных представителей сахаромицетов. В тропических регионах из природных источников выделяются дрожжи, активно бродящие при 35—40 °C, теплолюбивые представители есть и среди культурных рас, используемых в пищевой промышленности. В умеренном климате широко распространены виды, обитающие на весенних истечениях соков из деревьев — дуба, берёзы, клёна (Magnusiomyces magnusii [ syn. Endomyces magnusii], Endomyces vernalis, «дрожжи Надсона» — род Nadsonia). Эти виды прекращают бродить при температуре выше 25 °C. От температурных условий может зависеть и характер размножения грибков, это свойство используют для проверки чистоты культурных рас, применяемых в виноделии: образцы культуры инкубируют при 35 °C и при 15—20 °C, если наблюдается образование сумок при пониженной температуре, это свидетельствует о загрязнении образца дикими дрожжами.
Виды, постоянно обитающие в кишечнике теплокровных животных характеризуются необычными потребностями: высокое потребление витаминов и аминокислот, особые требования к кислотности среды, газовому составу. Так, Cyniclomyces guttulatus из кишечника кролика нормально растёт только в кислой среде при pH 2,0—5,6, в атмосфере с содержанием кислорода 2%, углекислого газа 10—15% и температуре 30—40 °C. По этой причине вид длительное время не поддавался искусственному культивированию и изучению.
Практическое значение
Пищевая промышленность
Дрожжи спиртового брожения широко используются в хлебопечении (хлебопекарные дрожжи), пивоварении (пивоваренные дрожжи) и виноделии, производстве кваса, сидра. Путём дистилляции сброженного сусла получают крепкие алкогольные напитки.
Чаще всего используют дикие формы или культурные расы Saccharomyces cerevisiae. Этот вид в литературе называют «пивные» или «пекарские» дрожжи, хотя в пивоварении используются и другие виды.
Винодельческие расы дрожжей получают названия в зависимости от местности из которой они происходят или от сорта винограда. Характерный вкус и букет вина зависит от множества факторов, в том числе и от расы используемых дрожжей.
Вредны для виноделия виды, образующие плёнки на поверхности сусла и относящиеся к слабо бродящим или вовсе не способным к спиртовому брожению — представители родов Пихия (Pichia), Кандида (Candida), особенно Pichia membranifaciens. Они вызывают повышение концентрации органических кислот в виноматериалах и ухудшают их качество. Другие вредные для виноделия дрожжи известны под названием «апикулятусы», они вызывают интенсивное брожение и подавляют развитие винных дрожжей. К сорнякам брожения виноделы относят и сахаромикод Людвига (Saccharomycodes ludwigii), который часто обнаруживается в составе чайного гриба.
Молочные дрожжи (Kluyveromyces lactis, Kluyveromyces marxianus) используются в производстве молочнокислых продуктов.
К видам, которые способны вредить в пищевой промышленности относится мицелиальный грибок ярровия липолитическая (Yarrowia lipolytica) (дрожжевая анаморфа ярровии описана в роде Candida). Этот вид продуцирует ферменты, расцепляющие жиры и белки. Он может развиваться в продуктах с высоким содержанием жира и вызывает прогоркание.
Фармацевтика
В фармацевтике препараты дрожжей применяются как витаминные и другие биологически активные добавки, главным образом благодаря высокому содержанию в них витаминов группы B. Препараты дрожжей используют обычно в виде сухих или жидких экстрактов. Суточная норма витаминов группы B содержится в 25 граммах сухих дрожжей.
Кормовые дрожжи
В бродильных производствах в качестве побочного продукта накапливаются большие количества дрожжевой биомассы, богатой белками, жирами и углеводами. Дрожжевая масса или дрожжевые экстракты могут использоваться как кормовые добавки. Специально в качестве кормовых дрожжей культивируют вид Candida utilis, который хорошо развивается на различных субстратах, содержащих как гексозы, так и пентозы. Сухие кормовые дрожжи производятся промышленностью ежегодно в количестве десятков миллионов тонн.
Для получения кормового белка из молочной сыворотки используют молочные дрожжи (Kluyveromyces).
Эремотециум Эшби (Eremothecium ashbyi) используют в животноводстве для получения концентратов рибофлавина. Для этого грибок выращивают на зерне или пищевых отходах, при этом субстрат сохраняет кормовые качества и обогащается витамином.
Производство этилового спирта
Для производства спирта путём брожения с последующей перегонкой и очисткой продукта используют картофель, зерно, патоку, отходы деревообрабатывающей и целлюлозно-бумажной промышленности. Так как большинство дрожжей не способны расщеплять целлюлозу, её предварительно осахаривают гидролизом, для этого обрабатывают кислотами или ферментами солода. Некоторые виды, такие, как пахисолен таннофильный (Pachysolen tannophilus) и пихия стебельковая (Pichia stipitis) (Scheffersomyces stipitis) активно сбраживают ксилозу и применяются для переработки в спирт отходов древесины.
При брожении выделяется большое количество углекислого газа, который утилизируют; ценными побочными продуктами спиртового брожения являются также фурфурол, метиловый спирт, некоторые высшие спирты (см. Сивушные масла).
Производство витаминов
Для промышленного получения витаминов используются виды и штаммы, известные как «сверхсинтетики». Например, сверхсинтетиком рибофлавина (витамин B2) является эремотециум Эшби (Eremothecium ashbyi), в природе встречающийся как паразит хлопчатника. Эремотециум легко поддаётся выращиванию в глубинной культуре на углеводных питательных средах, обогащённых органическим азотом и витаминами при температуре 28—30 °C. Выход рибофлавина составляет от 1—2 до 6 граммов на 1 литр среды.
Другие биохимикаты
Различные дрожжи могут применяться в промышленности и в лаборатории для проведения стереоселективных биохимических синтезов. Так, если в жидкую питательную среду с пекарскими дрожжами добавить ацетоуксусный эфир, то после брожения из смеси можно выделить чистый (+)(S)-энантиомер эфира β-оксимасляной кислоты[12], который затем используют для синтеза других оптически активных соединений[13].
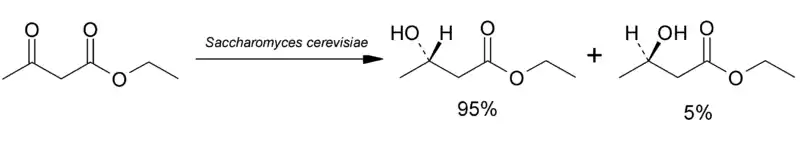
Другой способ состоит в том, что используют в качестве питательного субстрата синтетическую рацемическую смесь; при этом дрожжи потребляют только один энантиомер, а второй остаётся не изменённым. Оба метода имеют тот недостаток, что в результате синтеза (или потребления) оказывается доступным только один из двух энантиомеров.
Липомицеты (род Lipomyces) перспективны как продуценты липидов и амилаз.
Ярровия липолитическая (Yarrowia lipolytica) может применяться для различных биотехнологических синтезов на основе нефтепродуктов, так как этот вид хорошо развивается на нормальных алканах, выделяемых из нефти.
Аналитические тест-объекты
Виды и штаммы дрожжей различаются как по своим потребностям в витаминах, так и по способности самостоятельно синтезировать их. Эти особенности лежат в основе использования штаммов дрожжей с абсолютной потребностью в одном или двух витаминах в качестве биоиндикаторов. Существуют методики качественного и количественного определения при помощи дрожжевых тест-объектов витаминов группы B, являющихся факторами роста дрожжей — биотина, тиамина, инозита, пиридоксина, пантотеновой кислоты и никотиновой кислоты.
Патогены человека
Candida albicans — грибок, поселяющийся на коже и слизистых оболочках человека, обычно не вызывает патологических явлений, но при чрезмерном размножении может вызывать заболевания — кандидозы. Кандидоз может развиваться при заражении штаммом, обладающим повышенной патогенностью, нарушениях состава микрофлоры (дисбактериозах) или при снижении иммунитета, в частности, на фоне ВИЧ-инфекции.
Известны случаи продуцирования сахаромицетами токсичных для человека метаболитов.[14]
Патогены сельскохозяйственных растений
Мицелиальные или диморфные грибки родов Eremothecium и Nematospora паразитируют на растениях, преимущественно на плодах, и имеют значение как возбудители болезней культурных растений. Большинство видов распространены в субтропическом климате. Распространение инфекции и заражение растений происходит посредством насекомых.
Эремотециум Эшби (Eremothecium ashbyi) и Eremothecium gossypii (синонимы спермофтора хлопчатниковая, Spermophthora gossypii; нематоспора хлопчатниковая, Nematospora gossypii) поражают коробочки хлопчатника.
Нематоспора лещиновая (Nematospora coryli) вызывает гниль лесных орехов, а в Северной Африке и в Азии может наносить значительный ущерб урожаю фисташки. Внешне орехи выглядят неповреждёнными, однако семя темнеет а затем сгнивает и покрывается белым налётом аскоспор и мицелия гриба. Этот же вид встречается на томатах, ранее разновидность, поражающую томаты считали отдельным видом — нематоспора томатовая (N. lycopersici). На фасоли и других растениях паразитирует нематоспора фасолевая (Nematospora phaseoli).
Примечания
- Белякова, Дьяков, Тарасов, 2006, с. 186.
- Собченко и др., 2009, с. 43—44.
- Wijayawardene N. N., Hyde K. D., Lumbsch H. T., Liu Jian-Kui, Maharachchikumbura S. S. N., Ekanayaka A. H., Tian Qing, Phookamsak R. Outline of Ascomycota: 2017 // Fungal Diversity. — 2018. — Vol. 88, no. 1. — P. 167—263. — doi:10.1007/s13225-018-0394-8.
- Курс низших растений, 1981, с. 339—340.
- Собченко и др., 2009, с. 43.
- Гарибова, Лекомцева, 2005, с. 71—74.
- Lumbsch H. T., Huhndorf S. M. Myconet Volume 14. Part One. Outline of Ascomycota — 2009. Part Two. Notes on Ascomycete Systematics. Nos. 4751–5113 // Fieldiana Life and Earth Sciences. — 2010. — Vol. 1. — P. 1—64. — doi:10.3158/1557.1.
- Kurtzman C. P., Robnett C. J. Relationships among genera of the Saccharomycotina (Ascomycota) from multigene phylogenetic analysis of type species // FEMS Yeast Research. — 2013. — Vol. 13, no. 1. — P. 23—33. — doi:10.1111/1567-1364.12006. — PMID 22978764.
- Mühlhausen S., Kollmar M. Molecular phylogeny of sequenced Saccharomycetes reveals polyphyly of the alternative yeast codon usage // Genome Biology and Evolution. — 2014. — Vol. 6, no. 12. — P. 3222—3237. — doi:10.1093/gbe/evu152. — PMID 25646540.
- Shen Xing-Xing, Zhou Xiaofan, Kominek J., Kurtzman C. P., Hittinger C. T., Rokas A. Reconstructing the Backbone of the Saccharomycotina Yeast Phylogeny Using Genome-Scale Data // G3: Genes, Genomes, Genetics. — 2016. — Vol. 6, no. 12. — P. 3927—3939. — doi:10.1534/g3.116.034744. — PMID 27672114.
- Krassowski T., Coughlan A. Y., Shen Xing-Xing, Zhou Xiaofan, Kominek J., Opulente D. A., Riley R., Grigoriev I. V., Maheshwari N., Shields D. C., Kurtzman C. P., Hittinger C. T., Rokas A., Wolfe K. H. Evolutionary instability of CUG-Leu in the genetic code of budding yeasts // Nature Communications. — 2018. — Vol. 9, no. 1. — P. 1887 (8 p.). — doi:10.1038/s41467-018-04374-7. — PMID 29760453.
- С относительно высоким выходом (64 %) выделяется продукт с 95 %-ной энантиомерной чистотой, дальнейшая его очистка может проводиться химическими методами.
- Титце Л., Айхер Т. . Препаративная органическая химия: реакции и синтезы в практикуме органической химии и научно-исследовательской лаборатории / Пер. с нем. под ред. Ю. Е. Алексеева. — М.: Мир, 1999. — 704 с. — ISBN 5-03-002940-0. — С. 485—486.
- По «Ботанике» Беляковой; в более старых источниках пишут, что сахаромицеты не продуцируют токсины.
Литература
- Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. . Ботаника. Т. 1. Водоросли и грибы. — М.: Издательский центр «Академия», 2006. — 320 с. — ISBN 5-7695-2731-5.
- Гарибова Л. В., Лекомцева С. Н. . Основы микологии: Морфология и систематика грибов и грибоподобных организмов. — М.: Товарищество научных изданий КМК, 2005. — 220 с. — ISBN 5-87317-265-X.
- Жизнь растений. Энциклопедия в 6 тт. Т. 2: Грибы / Под ред. М. В. Горленко. — М.: Просвещение, 1976. — 479 с.
- Курс низших растений / Под ред. М. В. Горленко. — М.: Высшая школа, 1981. — 504 с.
- Собченко В. А., Храмченкова О. М., Бачура Ю. М., Цуриков А. Г. . Альгология и микология: Грибы и грибоподобные организмы. — Гомель: ГГУ им. Ф. Скорины, 2009. — 100 с. — ISBN 978-985-439-399-5.
Ссылки
- Saccharomycetales Kudrjanzev (англ.) на сайте MycoBank.
- Hemiascomycetes Engl. (англ.) на сайте MycoBank.
| Таксономия |
|---|