Кольчатые черви
Ко́льчатые че́рви, кольчецы́, аннели́ды (лат. Annelida, от anulus — кольцо) — тип беспозвоночных из группы первичноротых (Protostomia). Включают многощетинковых и малощетинковых червей, пиявок и мизостомид. Тип насчитывает около 18 тыс. видов, обитающих в морских и пресных водах и в толще почвы[1]. Некоторые виды пиявок перешли к наземному образу жизни в тропическом лесу[1]. Одни из наиболее известных представителей — дождевые черви. Размеры аннелид от 0,25 мм (у представителей рода Neotenotrocha) до 3 м (род Eunice)[2].
| Кольчатые черви | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Дождевой червь рода Lumbricus | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Тип: Кольчатые черви |
||||||||||||
| Международное научное название | ||||||||||||
| Annelida Lamarck, 1809 | ||||||||||||
| Классы | ||||||||||||
|
||||||||||||
| ||||||||||||
Строение и физиология
Сегментация
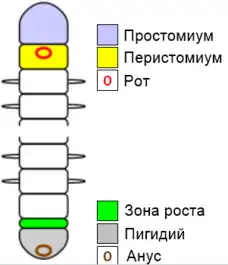
Тело кольчатых червей состоит из трех отделов: простомиума, туловища и пигидия. Удлиненное туловище состоит из сегментов — морфологически сходных единиц, располагающихся последовательно друг за другом от простомиума до пигидия. Снаружи сегменты отделены друг от друга неглубокими перетяжками. Создается впечатление, что тело кольчатых червей составлено из колец, откуда и произошло название этой группы. На переднем конце тела находится содержащая мозг головная лопасть — простомиум. На нем расположены органы чувств. Задняя, анальная, лопасть, или пигидий, несет анус. Первый сегмент — перистомиум — располагается непосредственно за простомиумом. Его вентральная поверхность охватывает рот.[3]
Простомиум и пигидий, хотя внешне и похожи на сегменты, на самом деле ими не являются, поскольку развиваются не из зоны роста. Зона роста находится непосредственно перед пигидием. В ней располагаются парные эктодермальные и мезодермальные телобласты. Эти клетки делятся, дифференцируются и дают начало новым сегментам. Рост червя осуществляется за счет последовательного встраивания все новых и новых сегментов на заднем конце тела. Такой тип роста называется телобластическим. В каждый данный момент времени самый молодой сегмент тела находится непосредственно перед пигидием, а самый старый (перистомиум) — позади простомиума.[3]
Все внутренние структуры кольчатых червей — придатки (когда они имеются), целомические мешки, нефридии и гонады — метамерны, т.е. повторяются в каждом сегменте. Однако некоторые органы и ткани проходят через все сегменты. Благодаря этому простой набор одинаковых элементов превращается в целостный, индивидуализированный организм. Основные интегрирующие структуры — это нервная и кровеносная системы, мускулатура и пищеварительная система.[3]
Кожно-мускульный мешок
Стенка тела кольчатого червя состоит из волокнистой коллагеновой кутикулы, железистого однослойного эпидермиса, соединительнотканного дермиса (его толщина может варьироваться) и мускулатуры, развивающейся из выстилки целома. Волокна кутикулы обычно расположены перекрестно-спирально. Они укрепляют стенку тела, препятствуют образованию аневризмов и часто придают телу червя переливчатый блеск. Некоторые полихеты, обитающие в трубках, лишены кутикулы, однако сами секретированные ими трубки напоминают кутикулу по структуре и составу. В состав эпидермиса обычно входит много железистых клеток, выделяющих слизь. Она укрепляет стенки норки червя и защищает поверхность его тела.[3]
Тонкие, эластичные щетинки выдаются наружу из эпидермиса. Они обеспечивают сцепление с субстратом при движении и выполняют другие функции. Каждый сегмент несет симметрично расположенные пучки простых щетинок: два дорсолатеральных и два вентролатеральных пучка. Встречаются и другие типы щетинок, и другие варианты их расположения на теле червя. Каждая щетинка формируется в небольшом мешочке — эпидермальном фолликуле, стенка которого состоит из фолликулярных клеток, а в основании находится особая клетка — хетобласт. Материал щетинки откладывается на поверхности длинных микроворсинок, которые затем деградируют, так что внутри сформированной щетинки остаются параллельные полые каналы. Основная часть щетинки выдается над поверхностью эпидермиса, а ее основание находится в фолликуле. Оно прикреплено к фолликулярным клеткам десмосомами, связанными с субэпидермальными мышцами. Их сокращение заставляет щетинку двигаться. Щетинки состоят из β-хитина — особой разновидности хитина, в которой полимерные, имеющие вид цепочек молекулы располагаются параллельно друг другу и имеют одинаковую полярность. Обычно β-хитин упруг и эластичен, но у некоторых аннелид щетинки укреплены склеротизированным белком и неорганическим материалом, например карбонатом кальция.[3]
Щетинки кольчатых червей принципиально отличаются от щетинок членистоногих. У членистоногих «щетинками» называют волосовидные или больше похожие на шипы сенсорные образования, подвижно сочлененные с наружным скелетом. Отклонение такой щетинки стимулирует сенсорный нейрон, прикрепленный к ее основанию. Щетинки членистоногих образуются в эпидермальном фолликуле, состоящем из одной или нескольких трихогенных клеток, которые секретируют щетинки, и одной или нескольких тормогенных клеток, формирующих подвижное сочленение. Фолликул включает также сенсорную клетку и окружающую ее текогенную клетку. Щетинки членистоногих бывают как полыми, так и сплошными, но в них никогда нет параллельных каналов, характерных для полихет. Они состоят из α-хитина — разновидности хитина, в котором соседние параллельные молекулы имеют противоположную полярность. В отличие от β-хитина α-хитин жесткий и неэластичный. Но эти его свойства могут быть дополнительно усилены благодаря отложению склеротизованного белка (у насекомых) или карбоната кальция (у многих ракообразных).[3]
Косоисчерченные мышцы кольчатых червей представлены наружным кольцевым и внутренним продольным слоями. Продольные мышцы обычно образуют четыре четко выраженных мышечных тяжа. У одних аннелид выстилка целома представлена мезотелием, образованным исключительно эпителиально-мускульными клетками мускулатуры стенки тела, а у других мускулатура отделена от целомической полости перитонеумом.[3]
Целом и кровеносная система

В каждом сегменте кольчатых червей симметрично залегают два целомических мешка, изолированных от целомов соседних сегментов поперечными септами. Левый и правый целомы в каждом сегменте отделены друг от друга продольными мезентериями, тянущимися вдоль мидсагиттальной плоскости тела; один из них расположен дорсально над кишкой, а второй вентрально, под ней. Положение септ соответствует положению перетяжек на поверхности тела. Каждая септа состоит из двух слоев мезотелия (один из них принадлежит предшествующему, а второй последующему сегментам) и залегающего между ними тонкого слоя соединительной ткани. Мезентерий организован так же, как и септы, но возникает в результате контакта стенок левого и правого целомов одного сегмента.[3]
Кольчатые черви, локомоция которых основана на использовании подвижных придатков или перистальтических сокращений всего тела, обычно имеют хорошо развитые септы, представляющие собой более или менее полные «переборки» между сегментами. Поскольку септы изолируют гидроскелет каждого отдельного сегмента, воздействие силы, возникающей при сокращении сегментарных мышц, ограничено только именно этим самым сегментом. С другой стороны, септы могут быть неполными или вообще подвергаются редукции. Подобное имеет место у кольчатых червей, ведущих сидячий образ жизни, а также у форм, которые используют иные механизмы локомоции, нежели упомянутые выше: например, ресничное движение (мелкие кольчатые черви), резкое изгибание тела или подтягивание за счет глотки, способной попеременно сильно выворачиваться и втягиваться (некоторые роющие кольчецы).[3]
Парные целомические полости каждого сегмента выстланы ресничным мезотелием, что следует рассматривать как примитивный признак. Участок мезотелиальной стенки целома, обращенный наружу, состоит из видоизмененных эпителиально-мускульных клеток, которые образуют и мышцы стенки тела. Из сократимых мезотелиальных клеток могут также формироваться радиальные и кольцевые мышцы септ, кольцевые мышцы кровеносных сосудов и мышцы стенки кишки (преимущественно кольцевые). На отдельных участках мезотелий представлен специализированными хлорагогенными клетками, которые желтым или коричневым слоем покрывают часть кишки и некоторые кровеносные сосуды. Хлорагогенная ткань — это главный центр, в котором синтезируются и запасаются гликоген и жир. Кроме того, в ее клетках накапливаются и обезвреживаются токсины, синтезируется гемоглобин, протекает катаболизм белков и образуется аммиак, осуществляется синтез мочевины.[3]
Целомическая жидкость, циркуляция которой обеспечивается работой ресничек и сокращением мышц стенки тела, содержит целомоциты. Эти клетки участвуют в реализации защитных реакций, а иногда и в газообмене (гемоглобинсодержащие целомоциты). Целом функционирует как гидроскелет. Усилия, развиваемые сокращающимися мышцами для изменения формы тела, должны передаваться гидроскелету. Сокращение продольных мышц заставляет тело расширяться, а сокращение кольцевых мышц приводит к его удлинению. В результате кольчатые черви могут проникать в почву и донные отложения, проделывая там ходы.[3]
В целомических полостях каждого сегмента осуществляется локальная циркуляция, однако транспорт в масштабах всего тела осуществляется хорошо развитой кровеносной системой, состоящей из кровеносных сосудов и сердец. У кольчатых червей с рудиментарными септами или совсем лишенных септ кровеносные сосуды сильно редуцированы или полностью исчезают. В подобных случаях транспорт по всему телу обеспечивает целомическая жидкость. Кровеносные сосуды и синусы представляют собой заполненные жидкостью каналы в компартменте соединительной ткани. В отличие от сосудов позвоночных они не выстланы эндотелием. Главные сосуды — спинной кровеносный сосуд в спинном мезентерии и брюшной кровеносный сосуд в брюшном мезентерии. По спинному сосуду кровь течет вперед, а по брюшному — назад. В каждом сегменте кровь возвращается из брюшного сосуда в спинной по капиллярной сети (плексусу), залегающему в стенке тела. Из спинного сосуда в брюшной кровь попадает по капиллярному плексусу, сосудам или синусам, охватывающим кишку. Главные кровеносные сосуды, особенно спинной, сократимы и прогоняют кровь за счет перистальтики. У некоторых кольчатых червей расширенный передний отдел спинного сосуда преобразован в мускулистое сердце, а у многих дождевых червей сердцами становятся несколько специализированных дорсовентральных сосудов, огибающих передний отдел кишечника.[3]
Гемоглобин, обычный дыхательный пигмент кольчатых червей, может находиться в целомической жидкости, крови, мышцах и в нервах. В целоме гемоглобин сконцентрирован в целомоцитах, а в кровеносной системе растворен в плазме крови. Газообмен осуществляется через стенку тела, придатки и жабры.[3]
Пищеварительная система
Пищеварительная система кольчатых червей представляет собой более или менее прямую трубку, открывающуюся ротовым отверстием на перистомиуме и заканчивающуюся расположенным на пигидии анусом. Кишка пронизывает септы и поддерживается мезентериями сверху и снизу. Пищеварительный канал состоит из эктодермальной передней кишки, энтодермальной средней кишки (желудок, кишечник) и эктодермальной задней, или прямой, кишки. Часто передняя кишка специализирована и подразделяется на два отдела: мускулистую глотку, способную выдвигаться или выворачиваться, и выстланный ресничным эпителием пищевод, связывающий глотку со средней кишкой.[3]
Нервная система и органы чувств
Центральная нервная система кольчатых червей представлена передним дорсальным мозгом, расположенным в простомиуме, и двумя продольными нервными стволами, проходящими вентрально. В простейшем случае мозг состоит из двух дорсальных надглоточных ганглиев. Пара окологлоточных коннектив соединяет мозг с двумя вентральными подглоточными ганглиями, приуроченными к перистомиуму. От последних отходят нервные стволы, тянущиеся вдоль всего тела. В каждом сегменте на нервных стволах располагается по одному ганглию, которые соединены между собой поперечной нервной комиссурой. Поскольку парные нервные стволы и их комиссуры в сегментах похожи на боковины и перекладины лестницы, подобный тип нервной системы называется брюшной нервной лестницей, или нервной системой лестничного типа. Каждая пара ганглиев по существу является «мозгом» сегмента, от которого симметрично в противоположные стороны отходит по одному сегментарному нерву. Последние, достигнув стенки тела, проникают в нее, отклоняются в дорсальном направлении и полностью или почти полностью опоясывают сегмент. Сегментарные нервы иннервируют мускулатуру стенки тела и сенсорные структуры данного сегмента.[3]
Сенсорные структуры кольчатых червей в основном представлены одноклеточными фото-, хемо- и механорецепторами. Они располагаются на голове и придатках, а также разбросаны по всему телу. Помимо этого полихеты (многощетинковые черви) обладают разнообразными органами чувств: глазками, глазами, статоцистами, специализированными органами химического чувства — нухальными органами. Глаза имеются и у некоторых пиявок. В большинстве случаев органы чувств концентрируются на простомиуме, перистомиуме и первых нескольких сегментах, но у некоторых полихет они располагаются также и на туловищных сегментах и даже на пигидии.[3]
Выделительная система
Экскреторные органы кольчатых червей представлены сегментарными нефридиями: метанефридиями или протонефридиями. Их свободные концы (воронки или терминальные клетки соответственно) часто располагаются на передней поверхности каждой септы. Они выступают в полость целома и омываются целомической жидкостью. Пронизывая септу, нефридиальный проток проходит в полость следующего сегмента и затем открывается во внешнюю среду. Как правило, метанефридии встречаются у аннелид, обладающих кровеносной системой, часть сосудов которой имеет покрытые подоцитами зоны фильтрации, протонефридии же свойственны кольчатым червям, лишенным кровеносной системы.[3]
Размножение и развитие
Кольчатые черви исходно раздельнополы. Продуцируемые ими гаметы выводятся во внешнюю среду через метанефридии. Их гонады располагаются в сегментах тела и представляют собой прикрепленные к мезотелию группы зародышевых клеток. Последние, освобождаясь, поступают в целомические полости и накапливаются в них. У зрелых особей целомические мешки полностью заполнены гаметами. Оказавшись в морской воде, гаметы копулируют. Образовавшиеся зиготы претерпевают спиральное дробление и дают начало личинкам — трохофорам. В процессе метаморфоза личиночная эписфера (верхнее полушарие трохофоры, ограниченное прототрохом) становится простомиумом, а область личинки, расположенная позади телотроха, становится пигидием. Сегменты, формирующие туловище, возникают в зоне роста, формирующейся перед телотрохом.[3]
Восстановительная регенерация хорошо развита среди аннелид. После удаления нескольких сегментов у многих видов сразу же образуется зона роста, в которой формируются новые сегменты (иногда столько же, сколько было утеряно).[4]
Тело полихеты Clymenella torquata, у которой постоянное число сегментов равно 22, может полностью восстанавливаться из 13 сегментов. Во время опыта были вырезаны участки из различных отделов тела. Утраченные сегменты из передней и задней частей тела были восстановлены во всех случаях в соответствии со своей исходной длиной. У Chaetopterus variopedatus один-единственный сегмент может наращивать недостающие участки тела как спереди, так и сзади, так что животное восстанавливается полностью. Напротив, Sabella никогда не восстанавливает более трёх сегментов; если же утеряно более трёх передних сегментов, то сегменты из задней части модифицируются в сегменты передней торакальной части. Особенно быстрой регенерацией обладает крона щупалец, которая крайне необходима при сборе пищи. Именно эта часть тела зачастую становится добычей рыб.[4]
У дождевых червей регенерацию, вероятно, контролирует нейросекреция передних ганглиев. Таким образом, передний конец тела может быть восстановлен, только в том случае, если один из этих ганглиев сохранился. Задняя часть тела восстанавливается полностью, однако чаще всего число новых сегментов не совпадает с первоначальным. Восстанавливающиеся животные становятся неподвижными; кроты пользуются таким поведением: они откусывают передние сегменты и складывают на хранение неподвижных дождевых червей в свои хранилища, расположенные в норах.[4]
Высокая способность тела аннелид к репаративной регенерации используется во многих случаях при бесполом размножении. Каудальная автотомия и последующая регенерация являются одним из этапов жизненного цикла дождевых червей. Деление на более или менее крупные части тела (фрагментация) с последующей регенерацией (архитомия) считается исключительным способом размножения и встречается, например, у олигохеты Enchytraeus fragmentosus. Способ деления, когда один сегмент может отделяться и из него образуется новая особь, встречается среди полихет, например у Dodecaceria caulleryi. Также распространена паратомия: при этом, как правило, в задней части тела образуются различной длины цепочки, состоящие из животных (столоны, зооиды), из которых далее формируются полностью дифференцированные особи. Этот способ размножения можно видеть, например, у Ctenodrilidae, Aeolosomatidae, Naididae. Такого рода образовавшиеся животные становятся носителями половых клеток, например, Syllidae; и поэтому особи, размножающиеся бесполым и половым путями у этих видов, обязательно связаны между собой.[4]
Иммунная система
Целомоциты олигохет делят на пять основных типов: базофилы и нейтрофилы (гиалиновые амебоциты), гранулоциты и ацидофилы (гранулярные амебоциты) и хлорагогенные клетки (элеоциты). Данные электронной микроскопии позволяют отнести базофилы к лимфоцитоподобным клеткам двух типов: с большим или меньшим количеством псевдоподий. Эти клетки демонстрируют структурное сходство с незрелыми лимфоцитами позвоночных. Нейтрофилы напоминают макрофаги позвоночных животных. В их вакуолях заметны различного рода включения, в том числе и бактериальные клетки. Гранулоциты и ацидофилы — клетки с большим количеством включений и вакуолей. Элеоциты — клетки самой разнообразной величины. У них отсутствуют псевдоподии. Цитоплазма переполнена гранулами. Данный класс клеток выполняет функцию обеспечения организма питательными веществами. У кольчатых червей процесс фагоцитоза, инкапсуляции, распознавания чужеродности, трансплантационного отторжения и адаптивного переноса обеспечивается в основном амебоцитами гиалинового типа: лимфоцитоподобными амебоцитами первого и второго типов (базофилами), а также нейтрофилами, характеризующимися способностью к активному фагоцитозу. Гранулярные клетки принимают незначительное участие в процессе фагоцитоза. Элеоциты вообще не фагоцитируют. Образование капсулы как реакции организма на крупные инородные частицы, каковыми являются, например, многоклеточные паразиты, осуществляется нейтрофилами, ацидофильными клетками и элеоцитами. Участие последних может быть объяснено их трофической функцией, но не иммунологической активностью. При отторжении алло- и ксенотрансплантатов в реакцию вступают в основном лимфоцитоподобные амебоциты и нейтрофилы-макрофаги.[5]
Изучение клеточного состава гемолимфы кольчатых червей привело к важному заключению. У данных животных впервые наблюдается дивергенция клеток с иммунологическими потенциями на два самостоятельных типа: макрофаги (нейтрофилы), обеспечивающие неспецифическую защиту, и лимфоцитоподобные амебоциты — участники специфических форм реагирования.[5]
Экология
Обитают в солёных и пресных водоемах и на суше. Встречаются виды — эктопаразиты и мутуалисты. Некоторые аннелиды — кровососущие, есть активные и пассивные хищники, падальщики и фильтраторы. Однако наибольшее экологическое значение имеют аннелиды, перерабатывающие почву; к ним относятся многие малощетинковые черви и даже пиявки. Почва, проходя через кишечник малощетинковых червей, становится более рыхлой и содержит больше влаги, необходимой для растений, а органика быстрее преобразуется в доступную для них форму. В почве может обитать от 50 до 500 червей на м².
Особенно многообразны морские формы, которые встречаются на разных глубинах вплоть до предельных (до 10—11 км) и во всех широтах Мирового океана. Они играют существенную роль в морских биоценозах и обладают высокой плотностью поселений: до 500—600 тыс. на 1 м² поверхности дна. Занимают важное положение в трофических цепях морских экосистем.[6][7]
Систематическое положение и классификация
Традиционно кольчатых червей сближают с членистоногими. Общими для них особенностями можно считать наличие постларвальной сегментации тела, связанные с этим особенности строения нервной системы (наличие надглоточного ганглия — головного мозга, окологлоточного нервного кольца и брюшной нервной цепочки). Из всех кольчатых червей по некоторым признакам наиболее сходны с членистоногими многощетинковые. Как и у членистоногих, у них развиты боковые придатки тела, параподии, которые считаются предшественниками ног членистоногих, а в циркуляции крови главную роль играет мускульная обкладка спинного сосуда. Наличие кутикулы нельзя считать чертой, уникально присущей кольчатым червям и членистоногим, поскольку кутикулярные эпителии довольно широко распространены в разных группах беспозвоночных. Более того, кутикула аннелид по большей части не содержит хитина, характерного для членистоногих. У аннелид, как и у некоторых других групп червей, хитинсодержащая кутикула имеется только на небольших участках тела: на «челюстях» в глотке и на щетинках.
Вместе с тем, по типу дробления, развитию и строению личинок — трохофор кольчатые черви близки к прочим трохофорным животным, например, моллюскам и сипункулидам.
Ряд исследователей полагает, что сходство с членистоногими по таким признакам, как сегментация, строение нервной системы, кутикула — результат конвергентной (параллельной) эволюции.
В соответствии с современными данными о происхождении двустороннесимметричных (Bilateria) представляется весьма вероятным, что метамерия и наличие целома (который у членистоногих формируется в эмбриогенезе) — исходные признаки для этой группы животных, утраченные позднее представителями многих типов[8].
В соответствии с новейшими молекулярными данными, ближайшие сестринские группы кольчатых червей — немертины, плеченогие и форониды, более далёкая родственная группа — моллюски, ещё более далекая — плоские черви, а наименее родственная среди всех первичноротых — клада, объединяющая головохоботных, круглых червей и членистоногих (в широком смысле)[9][10]. Таким образом, эти данные подтвердили концепцию Lophotrochozoa и Ecdysozoa.

К аннелидам причисляют от 7000 до 16 500 видов, разделяемых в разных классификациях на разное число классов. Традиционная классификация предполагает деление на 3 класса: многощетинковые черви, малощетинковые черви и пиявки. В качестве классов аннелид также рассматриваются эхиуры, мизостомиды и погонофоры (Pogonophora). Так, В. Н. Беклемишев (1964) предлагал делить аннелид на надклассы беспоясковых (классы многощетинковые и эхиуры) и поясковых (классы малощетинковые и пиявки). Помимо этих классов к аннелидам иногда относят представителей типа сипункулид.
Разными авторами предлагалось также другие системы типа, например, деление на классы:
- Мизостомиды (Myzostomida)
- Многощетинковые черви (Polychaeta) — наиболее многочисленный класс, преимущественно морские формы, например, морская мышь
- Поясковые черви (Clitellata), у которых на теле имеется характерный поясок, делятся на подклассы:
- Малощетинковые черви (Oligochaeta), к которым относятся дождевые черви
- Пиявки (Hirudinea)
- Acanthobdellida
- Branchiobdellida
В других системах предлагалось также выделять в качестве отдельных классов «аберрантных олигохет» — например, Aelosomatidae[11] и Potamodrilidae, лобатоцеребрид и др. В то же время архианнелиды (по крайней мере большинство из них), видимо, являются вторично упрощёнными в связи с мелкими размерами полихетами, Branchiobdellida относят к олигохетам, а Acanthobdellida — к пиявкам (см., например,[12]).
По данным молекулярных исследований последнего десятилетия, ни одна из этих систем не является удовлетворительной ([9][13] и др.). В соответствии с этими данными все вышеупомянутые «классы» (включая погонофор, эхиур и сипункулид), скорее всего, являются ближайшими родственниками отдельных групп полихет. Таким образом, «Polychaeta» — парафилетический таксон, а система типа нуждается в коренном пересмотре.
Альтернативная классификация
По данным World Register of Marine Species, на март 2020 года к типу относят следующие современные таксоны до ранга отряда включительно[14]:
- Класс Многощетинковые черви (Polychaeta)
- Подкласс Эхиуры (Echiura)
- Отряд Echiuroidea
- Подкласс Errantia (syn. Aciculata)
- Отряд Amphinomida
- Отряд Eunicida
- Отряд Phyllodocida
- Подкласс Sedentaria
- Инфракласс Canalipalpata
- Отряд Sabellida
- Отряд Spionida
- Отряд Terebellida
- Инфракласс Scolecida
- Инфракласс Canalipalpata
- Подкласс Эхиуры (Echiura)
- Класс Поясковые черви (Clitellata)
- Подкласс Пиявки (Hirudinea)
- Инфракласс Acanthobdellidea
- Инфракласс Настоящие пиявки[15] (Euhirudinea)
- Отряд Челюстные пиявки или бесхоботные пиявки (Arhynchobdellida)
- Отряд Хоботные пиявки (Rhynchobdellida)
- Подкласс Малощетинковые черви (Oligochaeta)
- Отряд Capilloventrida
- Отряд Crassiclitellata
- Отряд Enchytraeida
- Отряд Haplotaxida
- Отряд Lumbriculida
- Подкласс Пиявки (Hirudinea)
- Отряды incertae sedis
- Отряд Мизостомиды (Myzostomida)
По молекулярно-генетическим данным немецкого учёного Torsten Struck (2011) полихеты — это полифилетическая группа и она нуждается в распределении между Errantia и Sedentaria, причём в последнюю группу входят Clitellata. Мизостомиды, сипункулиды и ряд групп «полихет» считаются базально отходящими[16].
Эволюция
О происхождении кольчатых червей существует несколько теорий; как правило, их выводят от низших плоских червей. Ряд черт в организации кольчецов свидетельствует об их родстве с низшими червями. Так, у личинки кольчецов — трохофоры — имеются первичная полость тела, протонефридии, ортогональная нервная система и на ранних стадиях — слепой кишечник. Однако, как уже упоминалось, в соответствии с современными данными о происхождении двустороннесимметричных (Bilateria) представляется весьма вероятным, что метамерия и наличие целома — исходные признаки для этой группы животных, утраченные позднее представителями многих типов[8]. В случае правильности данной теории общие предки аннелид и плоских червей должны были напоминать скорее первых, чем последних.
Центральной группой кольчатых червей в свете молекулярных данных считаются многощетинковые, от которых, в связи с переходом к пресноводному и наземному образу жизни, произошли малощетинковые черви, давшие начало пиявкам.
Ископаемых остатков аннелид немного, но несколько форм известно со среднего кембрия. К аннелидам, ранее относили вендское ископаемое Сприггина, но сходство оказалось исключительно внешним. К кембрийским ископаемым относится Canadia, найденная в Берджес-Шейл. У Canadia и других кембрийских червей не было челюстей, но позднее у некоторых многощетинковых развился хитиновый челюстной аппарат, чьи остатки в ископаемом виде известны как сколекодонты. Они имеют вид чёрных зазубренных искривленных пластинок размером от 0,3 до 3 мм. Чаще всего сколекодонты встречаются в отложениях ордовика и девона, хотя известны даже из послепермских отложений. Фоссилизированных остатков самих червей мало. Великолепным исключением, например, служит недавно найденный отпечаток представителя палеозойских Machaeridia, позволивший всю эту группу расположить среди отрядов современных полихет[17].
Поскольку аннелиды являются бесскелетными животными, их ископаемые остатки крайне редки[18]. Многощетинковые черви встречаются чаще всего в виде челюстей, которыми обладали некоторые виды и минерализованных трубок, выполнявших роль скелета[19]. Некоторые представители эдиакарской биоты, такие как Dickinsonia иногда классифицируются как многощетинковые черви, хотя сходство не слишком выражено, чтобы утверждать это однозначно[20]. Cloudina, возрастом ок. 549—542 млн лет, принадлежащая к эдиакарской скелетной фауне также классифицируется некоторыми исследователями как аннелида, тогда как другие относят её к кишечнополостным[21][22]. До 2008 года самые ранние находки, которые можно надёжно отнести к полихетам, Canadia и Burgessochaeta, обе из канадских сланцев Бёрджес, имели возраст 505 млн лет и относилий к раннему кембрию[23]. Myoscolex, найденный в австралии, немного старше сланцев Бёрджес, также предположительно является аннелидой. Однако у него отсутствуют некоторые типичные черты аннелидов, и имеются некоторые черты, характерные для других типов животных[23]. В 2008 году С. Моррис и Дж. Пил (John Peel) сообщили об остатках Phragmochaeta в Сириус Пассет (север Гренландии) возрастом около 518 млн лет и сочли их наиболее древними остатками аннелид[20]. Активно обсуждался вопрос о том, является ли Wiwaxia из сланцев Бёрджес моллюском или аннелидом[23]. Разделение полихет на более мелкие таксономические группы произошло в раннем ордовике, около 488—474 млн лет. Только в ордовикских отложениях были обнаружены наиболее ранние челюсти аннелид, таким образом, наиболее поздний общий предок для всех существующих ныне аннелид не мог появится ранее этого срока[24]. К концу карбона, около 300 млн лет назад, уже существовали все существующие ныне группы свободно передвигающихся полихет[23]. Найдены минерализованные трубки, похожие на трубки современных sessile полихет[25] , однако первые трубки, надёжно относящиеся к полихетам, датируются юрским периодом, то есть имеют возраст не более 200 млн лет[23].
Наиболее ранние ископаемые остатки, определённо принадлежащие малощетинковым червям, относятся к третичному периоду (не более 65 млн лет). Предполагается, что их эволюция связана с появлением цветковых растений в раннем меловом периоде (130-90 млн лет)[26]. Находки в виде нор, частично заполненных каловыми гранулами могут служить свидетельством, что дождевые черви уже существовали в раннем триасе (251—245 млн лет)[26][27]. Окаменевшие остатки организмов, относящиеся к ордовику (472—461 млн лет), предварительно классифицируются как олигохеты, но справедливость этой классификации оспаривается некоторыми исследователями[26][28].
Примечания
- Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423.
- Struck, T. H., Halanych, K. M., Purschke, G. (2005). Dinophilidae (Annelida) is most likely not a progenetic Eunicida: evidence from 18S and 28S rDNA. Molecular Phylogenetics and Evolution 37: 619—623. Текст (англ.) (Дата обращения: 5 октября 2011)
- Э. Э. Рупперт, Р. С. Фокс, Р. Д. Барнс. Зоология беспозвоночных.
- Под редакцией В. Вестхайде и Р. Ригера. Зоология беспозвоночных.
- В. Г. Галактионов. Эволюционная иммунология. — 2005.
- Tulkki P. Effect of pollution on the benthos of Göthenberg // Helgoländer wiss. Meeresunters. — 1968. — 17, 1 — 4. — P. 209—215.
- López Gappa J. J., Tablado A., Magaldi N. H. Influence of sewage pollution on a rocky intertidal community dominated by the mytilid Brachydontes rodriguezi // Mar. Ecol. Prog. Ser. — 1990. — 63. — P. 163—175.
- http://soil.ss.msu.ru/~invert/main_rus/science/library/bilateria.pdf (недоступная+ссылка)
- Casey W. Dunn et al. Broad phylogenomic sampling improves resolution of the animal tree of life // Nature. 2008. V. 452, р.745-750.
- Элементы — новости науки: Новые данные позволили уточнить родословную животного царства
- Барнс Р. и др. Беспозвоночные: Новый обобщенный подход. — М.: Мир, 1992.
- Зоология беспозвоночных в двух томах. Том 1: от простейших до моллюсков и артропод. Под ред. В.Вестхайзе и Р.Ригера. М.: Т-во научных изданий КМК, 2008.
- BioMed Central | Full text | Annelid phylogeny and the status of Sipuncula and Echiura
- Тип Annelida (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 26 апреля 2020).
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М. : Сов. энциклопедия, 1986. — С. 473. — 831 с. — 100 000 экз.
- Phylogenomics. annelida.de. Дата обращения: 25 февраля 2018.
- Vinther, J., P. Van Roy, & D.E.G. Briggs. Machaeridians are Palaeozoic armoured annelids // Nature 451, 185—188.
- Rouse, G. The Annelida and their close relatives // Invertebrate Zoology (неопр.) / Anderson, D.T.. — Oxford University Press, 1998. — С. 202. — ISBN 0-19-551368-1.
- Briggs, D.E.G.; Kear, A.J. Decay and preservation of polychaetes; taphonomic thresholds in soft-bodied organisms (англ.) // Paleobiology : journal. — Paleontological Society, 1993. — Vol. 19, no. 1. — P. 107—135.
- Conway Morris, S.; Pjeel, J.S. The earliest annelids: Lower Cambrian polychaetes from the Sirius Passet Lagerstätte, Peary Land, North Greenland (англ.) // Acta Palaeontologica Polonica : journal. — Polish Academy of Sciences, 2008. — Vol. 53, no. 1. — P. 137—148. — doi:10.4202/app.2008.0110.
- Miller, A.J. A Revised Morphology of Cloudina with Ecological and Phylogenetic Implications (2004). Дата обращения: 12 апреля 2009. Архивировано 27 марта 2009 года.
- Vinn, O.; Zatoń, M. Inconsistencies in proposed annelid affinities of early biomineralized organism Cloudina (Ediacaran): structural and ontogenetic evidences (англ.) // Carnets de Géologie : journal. — 2012. — No. CG2012_A03. — P. 39—47. — doi:10.4267/2042/46095.
- Dzik, J. Anatomy and relationships of the Early Cambrian worm Myoscolex (англ.) // Zoologica Scripta : journal. — 2004. — Vol. 33, no. 1. — P. 57—69. — doi:10.1111/j.1463-6409.2004.00136.x.
- Budd, G. E.; Jensen, S. A critical reappraisal of the fossil record of the bilaterian phyla (англ.) // Biological Reviews of the Cambridge Philosophical Society : journal. — 2000. — May (vol. 75, no. 2). — P. 253—295. — doi:10.1111/j.1469-185X.1999.tb00046.x. — PMID 10881389.
- Vinn, O.; Mutvei, H. Calcareous tubeworms of the Phanerozoic (неопр.) // Estonian Journal of Earth Sciences. — 2009. — Т. 58, № 4. — С. 286—296. — doi:10.3176/earth.2009.4.07.
- Humphreys, G.S. Evolution of terrestrial burrowing invertebrates // Advances in Regolith (неопр.) / Roach, I.C.. — CRC LEME, 2003. — С. 211—215. — ISBN 0-7315-5221-0.
- Retallack, G.J. Palaeosols in the upper Narrabeen Group of New South Wales as evidence of Early Triassic palaeoenvironments without exact modern analogues (англ.) // Australian Journal of Earth Sciences : journal. — 1997. — Vol. 44, no. 2. — P. 185—201. — doi:10.1080/08120099708728303. (недоступная ссылка)
- Conway Morris, S.; Pickerill, R.K.; Harland, T.L. A possible annelid from the Trenton Limestone (Ordovician) of Quebec, with a review of fossil oligochaetes and other annulate worms (англ.) // Canadian Journal of Earth Sciences : journal. — 1982. — Vol. 19, no. 11. — P. 2150—2157. — doi:10.1139/e82-189.
Литература
- Аннелиды // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
Ссылки
| Таксономия | |
|---|---|
| В библиографических каталогах |
