Киноринхи
Кинори́нхи[2] (лат. Kinorhyncha, от др.-греч. κινέω — двигаю и ῥύγχος — рыло) — класс беспозвоночных животных из группы Scalidophora (некоторые исследователи рассматривают его как самостоятельный тип). Киноринхи обитают в поверхностном слое илистого грунта или между песчинками в морях по всему миру. Отличительной анатомической чертой киноринх является метамерия в строении кутикулы, мускулатуры, эпидермальных желёз и нервной системы. Тело разделено на 13 сегментов, первым из которых является хобот (интроверт). Они не имеют конечностей и двигаются благодаря втягиванию хобота. В составе хобота выделяют два подотдела: ротовой конус и центральную часть. У киноринх ротовой конус выдвигается, а не выворачивается, как у приапулид. Киноринхи раздельнополы, хорошо выражен половой диморфизм. Детали эмбрионального развития киноринх неизвестны. Вылупившиеся черви похожи на взрослых особей и сразу же приступают к питанию.
| Киноринхи | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Внешний вид Echinoderes komatsui | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Клада: Класс: Киноринхи |
||||||||||||
| Международное научное название | ||||||||||||
| Kinorhyncha Reinhard, 1881[1] | ||||||||||||
| Отряды | ||||||||||||
|
||||||||||||
| ||||||||||||
Первые представители группы были описаны в 1841 году. В настоящее время класс киноринх подразделяется на два отряда: Cyclorhagida и Homalorhagida. По состоянию на 2015 год известно около 220 видов киноринх, и число описанных видов постоянно растёт[3]. В 2016 году был секвенирован митохондриальный геном двух видов киноринх — Echinoderes svetlanae и Pycnophyes kielensis.
Анатомия и физиология
Общий план
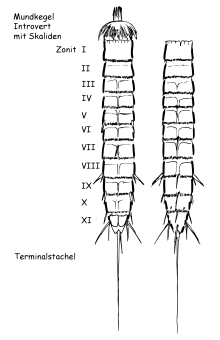
Киноринхи — червеобразные двусторонне-симметричные животные длиной от 100 мкм до 1,03 мм[4]. Самый маленький представитель, Cephalorhyncha liticola, достигает от 127 до 210 мкм в длину, а самый крупный, Pycnophyes greenlandicus, может достигать 1,035 мм[5]. Тело подразделяется на два отдела: хобот (интроверт) и туловище. Туловище в поперечном сечении может быть округлым, овальным, треугольным[4]. При помощи особых мышц хобот может втягиваться в туловище. Выворачивание хобота происходит за счёт давления, создаваемого при сжатии туловища в спинно-брюшном направлении. Тело в общей сложности состоит из 13 сегментов. Первым сегментом считают хобот, вторым — шейный отдел, который представляет собой серию пластинок, прикрывающих хобот, втягивающийся в тело. Форма и число пластинок шейного отдела служат важными признаками, использующимися в классификации киноринх. Собственно туловище метамерно разделено на 11 сегментов (зонитов), каждый из которых образован одной или несколькими кутикулярными пластинами[6].
В составе хобота выделяют два подотдела: ротовой конус и центральную часть. Ротовой конус представляет собой усечённый конус с ротовым отверстием на вершине, окружённым 9 стилетами, которые задействованы в сборе пищи. У киноринх ротовой конус выдвигается, а не выворачивается, как у приапулид[7]. Тонкая и гибкая кутикула в области конуса образует продольные складки. Поверхность центральной части несёт концентрические круги скалид — шипообразных кутикулярных образований. В состав каждого круга может входить разное число придатков. Скалиды, как правило, состоят из двух члеников[6].
Каждый сегмент несёт свои кутикулярные пластинки (плациды). Хотя внешне сегменты кажутся одинаковыми, во внутреннем строении киноринх метамерии не прослеживается. Количество и форма пластин сегментов активно используются в качестве систематических признаков. Как правило, один туловищный сегмент несёт три пластины: одну спинную (тергит) и две брюшные (стерниты). Кутикулярные пластинки на втором или третьем сегменте образуют замыкательный аппарат при полностью ввёрнутом интроверте[7]. На туловище располагаются шипы и другие придатки. Они представляют собой кутикулярные образования, в которые могут заходить отростки клеток гиподермы и нейроны. Шипы задействованы в локомоции животных и функционируют в качестве органов чувств[6].
Движение киноринх осуществляется за счёт выворачивания интроверта, заякоривания в субстрате при помощи скалид и последующего подтягивания тела[6].
Покровы
Тело киноринх покрыто плотным кутикулярным панцирем, который подстилается однослойной гиподермой. Кутикула киноринх состоит из хитина[7]. К кутикуле прикрепляется локомоторная мускулатура, более того, она определяет расположение всех внутренних органов. Кроме того, кутикула выполняет защитную роль. Как упоминалось выше, в типичном случае каждый сегмент покрыт одной спинной и двумя брюшными пластинами. Спинные и брюшные пластины каждого сегмента соединены гибкими волокнистыми связками, которые известны как плевральные пластины. Брюшные пластины соединяются при помощи волокнистого шва с хорошо развитым гребнем. Как правило, самые мощные кутикулярные покровы имеет первый тергит. У некоторых примитивных форм, таких как Zelinkaderes floridensis, толщина кутикулы одинакова по всей поверхности тела[8]. На поверхности кутикулы располагаются микроскопические шипики и волоски, направленные назад. На кутикуле также имеются тонкие папиллы, собранные в так называемые флоскули и «чувствительные пятна». Кроме того, на поверхности кутикулы находятся чувствительные щетинки и поры, которыми открываются наружу гиподермальные железы[9]. На четвёртом зоните (нередко лишь у самцов) могут присутствовать прикрепительные трубочки[7].
Поверх кутикулы находится тонкая трёхслойная мембрана 7—10 нм толщиной. Под ней расположен гомогенный слой толщиной от 1 до 2,5 мкм. На расстоянии 10—20 нм от гомогенного слоя залегает гиподерма. В некоторых случаях промежуток между гомогенным слоем и гиподермой достигает 0,5 мкм и заполнен рыхлым тонковолокнистым материалом, который представляет собой самый внутренний, третий слой кутикулы. В местах прикрепления мышц гомогенный слой и гиподерма контактируют непосредственно. Кутикулярные пластины пронизаны многочисленными ветвящимися каналами, которые особенно многочисленны в местах прикрепления мышц к кутикуле. Каналы заполнены тонковолокнистым материалом, который также образует третий слой кутикулы[9].
В области плевральных пластин кутикула состоит из наружной осмофильной мембраны, плотного гомогенного слоя толщиной 0,5 мкм, 5—6 слоёв толстых фибрилл и тонковолокнистого внутреннего слоя толщиной 0,2—0,3 мкм. На её поверхности заметны мельчайшие продольные гребни. Фибриллы соседних слоёв залегают перпендикулярно друг другу[9].
Передние края всех пластин, а также обращённые к брюшному шву края стернитов имеют характерные утолщения, представляющие собой кутикулярные выросты, направленные внутрь тела, — пахицикли. К ним крепятся хорошо развитые продольные мышцы[9].
Кутикула, покрывающая хобот, заметно тоньше и гибче туловищной кутикулы. Под наружной мембраной находится плотный слой толщиной 0,4—0,6 мкм, под которым залегает слой тонковолокнистого материала толщиной 1—3 мкм[9].
Рост тела киноринх сопровождается сбрасыванием старой кутикулы в ходе линьки. При линьке в клетках наружного слоя гиподермы накапливаются пузырьки, происходящие от эндоплазматического ретикулума. Когда плотный гомогенный слой кутикулы отходит от гиподермы, содержимое пузырьков, представляющее собой материал новой кутикулы, изливается наружу. Как и при линьке насекомых, происходит частичный лизис старой кутикулы, которая образует экзувий. Киноринха вспарывает его при помощи скалид и особых спинных зубцов и выходит наружу. После этого новая кутикула расправляется и становится жёсткой[10].
У многих киноринх кутикула чрезвычайно гидрофобна. Эта особенность используется для извлечения киноринх из субстрата, при котором пробы грунта сильно встряхивают, и киноринхи прилипают к пузырькам воздуха, поднимаясь вместе с ними на поверхность воды[11].
Гиподерма представляет собой однослойный погружённый эпителий. Клетки гиподермы связаны с кутикулой посредством полудесмосом. Апикальные участки соседних гиподермальных клеток связаны десмосомами, а в других местах их мембраны удалены на расстояние 15—20 нм; промежуток между ними заполнен особым тёмным материалом. Выросты одной клетки могут входить в выемки другой, формируя некое подобие замка. В гиподерме также выявляются септированные контакты. В гиподерме находятся метамерно расположенные слизистые железы, которые открываются на кутикуле специальными отверстиями и выделяют на её поверхности слой слизи[7]. Со стороны полости тела гиподерма подстилается мелкогранулярным внеклеточным матриксом толщиной 20 нм, который, по сути, является базальной мембраной[12].
Клетки гиподермы очень физиологически активны. Они имеют крупные овальные ядра, в их цитоплазме хорошо развиты гранулярный эндоплазматический ретикулум и аппарат Гольджи. В них также можно обнаружить хорошо выраженные ламеллярные структуры, часто свёрнутые в глобулы, липоидные включения, секреторные гранулы и везикулы разных размеров с гомогенным содержимым. В промежутках между гиподермальными валиками клетки гиподермы бедны органеллами. В тех местах, где мышцы крепятся к кутикуле, гиподерма пересечена мощными пучками тонофиламентов[12].
Мускулатура
Киноринхи имеют своеобразные поперечнополосатые мышцы. Саркомеры разделяются рядами так называемых Z-тел, которые представляют собой веретеновидные электронноплотные структуры. К ним крепятся актиновые филаменты диаметром 7 нм. Толстые (25 нм) миозиновые волокна могут вдвигаться в пространство между Z-телами и даже заходить в соседний саркомер. Благодаря этой особенности мускулатура киноринх способна к сверхсокращению (то есть сокращению более чем на 50 %). Z-тела могут располагаться хаотически или зигзагом, и границы саркомеров нередко оказываются сильно изогнутыми. В миоцитах киноринх чётко выражены сократимая и электронносветлая саркоплазматическая часть, бедная органеллами[13].
Скелетные мышцы крепятся к кутикуле через тонкие прослойки гиподермы. Таким образом, в месте прикрепления есть два ряда десмосомных пластинок: между мышечной и гиподермальной клетками и между гиподермальной клеткой и кутикулой. Как и для кутикулы, для мускулатуры киноринх характерна метамерия[13].
Скелетная мускулатура киноринх представлена продольными, дорсовентральными и диагональными мышцами. Продольные мышцы представлены двумя парами продольных мышечных лент: вентро- и дорсолатеральных. У некоторых представителей, таких как Zelinkaderes floridensis, продольные мышцы распространены равномерно по периметру туловища, у других, таких как Echinoderes, Pycnophyes и Kinorhynchus, они собраны в тяжи[4]. Спинные тяжи развиты значительно слабее брюшных, потому что животные чаще и сильнее изгибаются на брюшную сторону. Каждая лента состоит из отдельных продольных мышц, крепящихся к пахициклям. В последний сегмент мышцы практически не заходят, поскольку крепятся к утолщениям кутикулы вблизи гонопора на предпоследнем сегменте. При укорочении тела из-за синхронного сокращения продольных мышц отдельные сегменты могут заходить друг на друга. В каждом сегменте есть две дорсовентральные мышцы, соединяющие тергит и стерниты. Киноринхи имеют большое количество мышц-протракторов и ретракторов, которые обеспечивают выворачивание и вворачивание интроверта. Кольцевая мускулатура в стенке тела развита плохо. Отдельные кольцевые мышцы есть в интроверте. По набору мышечных элементов к киноринхам наиболее близки приапулиды, лорициферы и волосатики[13].
Нервная система
Нервная система киноринх прилегает к гиподерме, поэтому базальная мембрана, подстилающая гиподерму, одновременно отделяет нервную систему от полости тела. Центральную нервную систему составляют окологлоточное нервное кольцо (кольцевой мозг) и двойной брюшной нервный ствол. Мозг широкий, имеет вид воротничка и состоит из трёх колец[14]. Передний край мозга состоит из 10 скоплений биполярных чувствительных нейронов, которые иннервируют органы чувств переднего конца тела. Задняя часть мозга также состоит из тел нейронов, которые могут быть как собраны, так и не собраны в ганглии. Из заднего мозга выходят 8 продольных нервов, которые иннервируют шею и туловище[14]. У Zelinkaderes floridensis от заднего мозга отходят 12 продольных нервов, а у Pycnophyes dentatus — 7[4]. Промежуток между передней и задней частями мозга занят нейропилем. В его средней части залегают немногочисленные нейроны. Нервная ткань занимает весомую долю объёма интроверта, и мышцы-ретракторы проходят прямо сквозь нейропиль. Нервный ствол имеет метамерное строение, соответствующее наружной метамерии[15].
В состав периферической нервной системы киноринх входят латеральные, дорсо- и вентролатеральные нервы, связанные кольцевыми комиссурами, а также чувствительные клетки, иннервирующие соматические органы чувств. Совокупность элементов периферической нервной системы, которая повторяется в каждом сегменте, называется нейрозонитом[15].
Мускулатура у киноринх иннервируется брюшным нервным стволом, а также дорсо- и вентролатеральными нервами. Примечательно, что в нервную ткань заходят непосредственно отростки мышечных клеток. Ротовой конус иннервируется 10 нервами, выходящими из задней части мозга. Ключевым элементом нервной системы, связанным с пищеварительной системой, является стоматогастрическое нервное кольцо. Строго говоря, чаще всего оно незамкнуто и имеет, скорее, подковообразную форму[16]. Оно принимает нисходящие нервы ротового конуса и даёт начало глоточным нервам. В глоточном эпителии имеются чувствительные клетки, а в выстилке пищевода многочисленны нервные клетки[15]. Нейронов, иннервирующих кольцевые и продольные мышцы средней кишки, не обнаружено, поэтому, возможно, они сокращаются автономно. Возможно также, что их сокращения стимулируются чувствительно-двигательными клетками кишечного эпителия[4].
В целом план строения нервной системы киноринх схож с таковым у приапулид, лорицифер и волосатиков[17].
Органы чувств
В покровах киноринх нет свободных нервных окончаний из-за плотной кутикулы. Дендритные отростки сенсорных нейронов связаны с особыми органами чувств: флоскулями, чувствительными пятнами, щетинками, железистыми трубочками, шипами, скалидами и другими придатками. Они функционируют как хемо- и механорецепторы[4]. Сенсорные нейроны киноринх представлены биполярными нейронами, тела которых лежат в гиподерме вблизи соответствующих органов чувств. Флоскули представляют собой кутикулярные выросты, в центре которых находится вогнутый диск с по́рой, окружённый папиллами. Расположение чувствительных пятен на туловище киноринх видоспецифично[4]. В основании щетинок могут находиться железы, и на вершине щетинки нередко имеется отверстие, через которое дендритные отростки сообщаются с внешней средой. Главным комплексным органом чувств является интроверт в вывернутом состоянии. В глотке имеются особые глоточные стилеты, в которые входят дендритные отростки; по-видимому, они функционируют в качестве хемо- и механорецепторов. На заднем конце тела самцов киноринх есть копулятивные шипы, которые не только выполняют вспомогательные функции при спаривании, но и являются органами чувств. Каждый копулятивный шип заключает в себе дендритный отросток и вырост железистой клетки. Многие киноринхи имеют парные глазки на интроверте. Они выглядят как оранжевые чашечки вокруг линзообразных утолщений кутикулы. Нередко киноринхи, лишённые глазков, всё-таки обладают светочувствительностью[18].
Пищеварительная система
В состав пищеварительной системы киноринх входит выстланная кутикулой передняя кишка, средняя кишка, образованная клетками с апикальной щёточной каёмкой, и кутикуляризованная задняя кишка. На всём своём протяжении пищеварительная трубка сохраняет клеточное строение[4]. Слюнные и пищеварительные железы не обнаружены[4]. Ротовое отверстие находится на конце тела и окружено кутикулярными губами. Оно открывается в выстланную кутикулой буккальную трубку, имеющую многочисленные кутикулярные складки, которые, как предполагают, играют роль фильтра. В ней также находятся 3 или 4 круга стилетов. За буккальной трубкой следует сосущая глотка, передний конец которой может нести кутикулярное кольцо (глоточную корону). Для усиления сосущего эффекта глотка может частично выталкиваться наружу специальными протракторами, а голова при этом смещается ближе к туловищу[4]. Вокруг переднего конца глотки находятся 10 железистых клеток с одной ресничкой, их тела залегают между глоткой и стоматогастрическим нервным кольцом, а апикальные концы открываются по периметру глоточной короны. Глотка имеет внутренний эпителиальный и наружный мышечный слой, поверх которого находится слой внеклеточного матрикса (базальная пластинка). Мышечный и эпителиальный слои глотки разделены такой же пластинкой. В эпителии глотки, помимо эпителиальных клеток, есть железистые, чувствительные и нервные клетки. Мускулатура глотки представлена радиальными и кольцевыми мышечными клетками. Глотка (как, вероятно, и все органы киноринх) имеет постоянный клеточный состав. После глотки следует пищевод, короткий, выстланный кутикулой, в состав которого входят эпителиальные и нервные клетки. Эпителий средней кишки представлен высокопризматическими клетками с хорошо выраженной щёточной каёмкой. Кроме того, в нём встречаются отдельные железистые клетки с крупными зимогеновыми гранулами и мощно развитым шероховатым эндоплазматическим ретикулумом, а также моноцилиарные чувствительные клетки. Задняя кишка образована уплощённым эпителием и выстлана кутикулой, которая постепенно утолщается кзади, достигая в задней части толщины наружной кутикулы. В клетках задней кишки очень много митохондрий, что позволяет предполагать наличие у них функции регуляции ионного баланса[19]. Задняя кишка открывается наружу анальным отверстием на конце 13-го зонита[14].
Полость тела
Полость тела киноринх (гемоцель) редуцирована, представлена щелевидными пространствами между органами и заполнена жидкостью. Она лишена специализированной клеточной выстилки. Полостная жидкость содержит хлопья аморфного электронноплотного вещества, которое, возможно, является дыхательным пигментом. В ней также встречаются многочисленные амёбоциты, содержащие электронноплотные гранулы и вакуоли неправильной формы с тёмным рыхлым содержимым[20]. Амёбоциты могут даже занимать большую часть объёма гемоцеля. Их функция неизвестна[21].
Выделительная система
Выделительная система киноринх представлена парой протонефридиев, которые залегают вдоль кишки рядом с дорсовентральной мышцей десятого сегмента. Каждый протонефридий состоит из трёх двужгутиковых терминальных клеток, которые лежат в гемоцеле, длинной канальцевой клетки и нефридиопоровой клетки. У Pycnophyes kielensis протонефридий включает 22 двужгутиковые терминальные клетки, две двужгутиковые канальцевые клетки и нефридиопоровую клетку. У P. greenlandicus имеется 11 терминальных клеток и 2 канальцевые[4]. Протоки протонефридиев открываются нефридиопорами на боковой поверхности 11-го сегмента. Количество выводных каналов и число пор могут варьировать от 6 до нескольких десятков[22]. В отличие от приапулид и лорицифер, у киноринх выделительная и половая системы независимы друг от друга[14].
Половая система
Бесполое размножение у киноринх никогда не наблюдалось[4]. Киноринхи раздельнополы, хорошо выражен половой диморфизм. У самок гонады располагаются сбоку от кишки в полости тела и тянутся от 5-го до начала 13-го зонита. В самом центре гонады (яичника) находится крупный зрелый ооцит с многочисленными желтковыми гранулами, а другие, мелкие, ооциты выполняют роль питающих клеток. В предпоследнем сегменте от яичника отходит короткий яйцевод, расширенная передняя часть которого образует подобие воронки. Рядом с яйцеводом находятся крупные гиподермальные железы, а непосредственно над ним в каждой гонаде находится мешковидный семяприёмник. Рядом с гонопором находятся различные железисто-чувствительные органы. Киноринхи не отличаются большой плодовитостью: в каждом яичнике зрелости достигают лишь 4 или 5 ооцитов. Низкая плодовитость свойственна для обитателей мейобентоса, к которым относятся киноринхи. Оплодотворение, по-видимому, происходит около протока семяприёмника. Связанные с яйцеводом железы формируют плотную оболочку яйца перед откладыванием его в грунт[23].
Мешковидные семенники располагаются сбоку от кишечника, начинаясь от границы 5-го и 6-го сегментов, и тянутся до гонопоров на границе двух последних сегментов. В передней части семенника залегают сперматогонии, далее следуют сперматоциты и сперматиды задних стадий, а в задней половине видны плотно упакованные сперматозоиды. Сперматозоиды киноринх имеют несколько модифицированное строение. Они представляют собой веретеновидные клетки, длина которых составляет более 10 % от длины взрослого животного. Центральную часть сперматозоида занимает очень длинное и тонкое ядро. Типичной акросомы нет. Сперматозоиды киноринх способны к червеобразным движениям за счёт изгибания клетки. Задняя широкая часть семенника в 12-м сегменте резко сужается в тонкий семяпровод, который открывается гонопором на конце длинной папиллы, образованной гибкой кутикулярной связкой между стернитами 12-го и 13-го сегментов. Вокруг гонопора обычно располагается густой венчик полых кутикулярных щетинок. Рядом с гонопором находятся копулятивные шипы. Они обладают чувствительностью и служат для плотного фиксирования сперматофора на задних сегментах и поддержания его во время копуляции. При копуляции сперматофор прикрепляется около гонопора самки[23]. У некоторых видов при копуляции две особи сцепляются задними концами тела[24].
Развитие
Детали эмбрионального развития киноринх неизвестны. Самки откладывают оплодотворённые яйца по одному в тот же субстрат, где живут взрослые животные. Самка прикрепляет их к частицам грунта за счёт клейкой оболочки. Яйца богаты желтком. В яйце формируется червеобразная личинка, которая сворачивается по окружности яйца. По строению она очень близка к личинке волосатиков. По мере созревания личинки оболочка яйца истончается. При вылуплении личинка распрямляется и выходит из яйца, используя шипы интроверта. Вылупившиеся черви похожи на взрослых особей и сразу же приступают к питанию. Ювенильные формы, как и взрослые особи, состоят из 13 сегментов, но отличаются от взрослых более крупными интровертом и глоткой, кроме того, все их придатки ещё очень мягкие и несклеротизированные[25]. В процессе роста молодые животные периодически линяют, но по достижении взрослого состояния линьки прекращаются[14] (однако у Z. floridensis и Antygomonas oreas наблюдались линьки во взрослом состоянии[4]). Всего имеется 6 ювенильных стадий, и с каждой линькой увеличивается число туловищных сенсилл и головных скалид[24].
Распространение и экология
Киноринхи — исключительно свободноживущие[4] морские мейобентосные животные, распространены по всему миру. Их можно найти в составе мейофауны песчаных пляжей, заиленных песков, мягких илов, богатых органикой. Обычно они населяют первые 3 см грунта. Ключевым фактором, ограничивающим глубину, на которой могут жить киноринхи, является доступность кислорода. Киноринх находили от литоральной зоны до абиссали — до глубины 5300 м[4][7]. Плотность популяций киноринх может составлять от 45 особей на 10 см2, как в стоячих антарктических водах, до 1—10 особей на 10 см2 на глубоководьях[4]. Иногда киноринхи обнаруживаются на поверхности водорослей и других беспозвоночных, таких как многощетинковые черви, мшанки, губки и моллюски[26]. Пресноводные и наземные формы среди киноринх отсутствуют[8]. Они очень чувствительности к загрязнениям воды[27].
Киноринхи часто встречаются в биотопах вместе с веслоногими рачками, нематодами, тихоходками, турбелляриями, инфузориями. Как правило, в популяциях поровну самок и самцов. Меньше всего ювенильных особей можно встретить зимой, а наибольшая доля половозрелых особей в умеренных широтах отмечается в конце зимы — начале весны. В тропиках размножение киноринх происходит круглый год[26].
Покровы киноринх выделяют слизь, которая склеивает частички субстрата, и нередко животные сами облепляют себя песчинками. Питаются киноринхи диатомовыми водорослями, мелкими частицами органического детрита или и тем, и другим. Некоторые виды фильтруют бактерии с помощью фильтра в буккальной трубке[4]. Хищные формы неизвестны. Питание осуществляется только при вывернутом интроверте[8]. Питается ли кто-то киноринхами, доподлинно неизвестно[26][14].
У киноринх впервые эндосимбионты были обнаружены в кишечном эпителии. Бактериальные клетки локализованы в цитоплазме только определённых клеток кишечника (бактериоцитов). Бактериоциты располагаются в центральной части средней кишки среди обычных высокопризматических эпителиальных клеток. Каждый бактериоцит может содержать более сотни бактериальных клеток. Некоторые учёные утверждают, что бактерии в кишечнике киноринх являются хемолитоавтотрофными окислителями серы, однако эта гипотеза пока не доказана. Возможно, что эти бактерии в действительности являются не эндосимбионтами, а поглощаются кишечником при питании[4]. В кишечных клетках киноринх могут паразитировать микроспоридии[28]. В кишечнике Kinorhynchus yushini и Z. floridensis были обнаружены паразитические апикомплексы[4].
Классификация и филогения

В настоящее время киноринх рассматривают в ранге класса в составе группы Scalidophora. Класс киноринх подразделяется на два отряда[29]:
- Отряд Cyclorhagida (Zelinka, 1896) Higgins, 1964. Наиболее широко распространённый и разнообразный класс киноринх. У представителей отряда втягиваться может только первый зонит. Кутикула второго зонита (шейного отдела) подразделяется на 14—16 пластинок, которыми покрыт передний конец туловища при втягивании хобота. Туловище снабжено шипами, его сегменты округлые в поперечном сечении. Часто встречаются в морском иле и песке в приливно-отливной зоне[24][30]. В отряде выделяют следующие семейства[31]:
- Antygomonidae Adrianov & Malakhov, 1994;
- Cateriidae Gerlach, 1956;
- Dracoderidae Higgins & Shirayama, 1990;
- Echinoderidae Zelinka, 1894;
- Semnoderidae Remane, 1929;
- Zelinkaderidae Higgins, 1990;
- Cyclorhagida incertae sedis.
- Отряд Homalorhagida (Zelinka, 1896) Higgins, 1964. Относительно крупные киноринхи (до 1 мм). Могут втягиваться два первых зонита. Кутикула шейного отдела подразделяется на не более чем 8 пластинок. В роли замыкателя при втягивании интроверта выступает третий сегмент. Туловище несёт несколько шипов. Туловищные зониты треугольные в поперечном сечении. Обитатели морских илов[32][30]. В отряде выделяют следующие семейства[33]:
- Neocentrophyidae Higgins, 1969;
- Pycnophyidae Zelinka, 1896.

В настоящее время киноринх включают в состав клады линяющие (Ecdysozoa), что подтверждается как морфологическими, так и молекулярными данными. Однако стероидных гормонов линьки у них выявлено не было[4]. Наиболее близкие родственники киноринх — приапулиды, которые являются их сестринской группой. Раньше киноринх включали в состав группы головохоботные (Cephalorhyncha), которая также содержит приапулид, волосатиков и лорицифер. Однако, когда выяснилось, что волосатики более близки к нематодам, то приапулид, лорицифер и киноринх стали объединять в группу Scalidophora. Впрочем, её монофилия находится под вопросом[34].
Недавний молекулярный анализ в целом подтвердил традиционную классификацию киноринх, основанную на морфологических признаках. Однако он показал, что Cyclorhagida — полифилетическая группа, так как входящий в неё род Dracoderes, по-видимому, является сестринской группой Homalorhagida или даже входит в состав этого отряда. Монофилия Homalorhagida подтвердилась молекулярными данными[35].
В 2016 году был секвенирован митохондриальный геном двух видов киноринх — Echinoderes svetlanae и Pycnophyes kielensis. Порядок генов в митохондриальных геномах у киноринх уникален для Metazoa и царства животных вообще. У обоих исследованных видов дуплицирован ген метиониновой тРНК[36].
История изучения
Впервые киноринхи были описаны французским натуралистом Феликсом Дюжарденом. В 1841 году он собирал морских беспозвоночных на северном побережье в Бретани во Франции. В 1863 году найденное им животное получило название Echinoderes dujardinii Claparède, 1863[35]. В 1881 году оно было выделено в самостоятельную группу Kinorhyncha. После открытия первых киноринх несколько исследователей занялись описанием представителей этой группы, однако в силу новизны группы в их описаниях было много неточностей. В конце XIX — начале XX века К. Зелинка опубликовал ряд работ, в которых упорядочивал данные других исследователей и описал несколько новых видов киноринх[5]. Важный вклад в развитие систематики группы внёс американский учёный Роберт Хиггинс, описавший многие виды киноринх и, в частности, предложивший выделение семейства Zelinkaderidae[37]. В разное время киноринх относили к таким таксонам, как Aschelminthes, Nemathelminthes, Pseudocoelomata и Cycloneuralia. В 1994 году было предложено выделение группы линяющие, к которой был отнесён и этот класс[4]. Активное описание новых видов и родов киноринх продолжается и по сей день[38][39][40].
Примечания
- Reinhard W. Über Echinoderes und Desmoscolex der Umgegend von Odessa // Zoologischer Anzeiger. — 1881. — Bd. 2, № 1. — S. 403—448.
- Киноринхи / Чесунов А. В. // Канцелярия конфискации — Киргизы. — М. : Большая российская энциклопедия, 2009. — С. 720. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 13). — ISBN 978-5-85270-344-6.
- Altenburger Andreas, Rho Hyun Soo, Chang Cheon Young, Sørensen Martin Vinther. Zelinkaderes yong sp. nov. from Korea - the first recording of Zelinkaderes (Kinorhyncha: Cyclorhagida) in Asia (англ.) // Zoological Studies. — 2015. — 5 February (vol. 54, no. 1). — ISSN 1810-522X. — doi:10.1186/s40555-014-0103-6.
- Neuhaus B. Ultrastructure, Biology, and Phylogenetic Relationships of Kinorhyncha (англ.) // Integrative and Comparative Biology. — 2002. — 1 July (vol. 42, no. 3). — P. 619—632. — ISSN 1540-7063. — doi:10.1093/icb/42.3.619.
- Handbook of Zoology, 2013, p. 181.
- Адрианов, Малахов, 1994, с. 5—10.
- Рупперт, Фокс, Барнс, 2008, с. 53.
- Вестхайде, Ригер, 2008, с. 753.
- Адрианов, Малахов, 1994, с. 10—31.
- Адрианов, Малахов, 1994, с. 32.
- Вестхайде, Ригер, 2008, с. 754.
- Адрианов, Малахов, 1994, с. 33—35.
- Адрианов, Малахов, 1994, с. 36—51.
- Рупперт, Фокс, Барнс, 2008, с. 55.
- Адрианов, Малахов, 1994, с. 51—71.
- Herranz M., Pardos F., Boyle M. J. Comparative morphology of serotonergic-like immunoreactive elements in the central nervous system of kinorhynchs (Kinorhyncha, Cyclorhagida). (англ.) // Journal Of Morphology. — 2013. — March (vol. 274, no. 3). — P. 258—274. — doi:10.1002/jmor.20089. — PMID 23109054.
- Адрианов, Малахов, 1994, с. 72.
- Адрианов, Малахов, 1994, с. 72—101.
- Адрианов, Малахов, 1994, с. 107—120.
- Адрианов, Малахов, 1994, с. 123—125.
- Рупперт, Фокс, Барнс, 2008, с. 54.
- Адрианов, Малахов, 1994, с. 125—128.
- Адрианов, Малахов, 1994, с. 128—149.
- Вестхайде, Ригер, 2008, с. 755.
- Адрианов, Малахов, 1994, с. 150—157.
- Адрианов, Малахов, 1994, с. 161—164.
- Chariton A. A., Ho K. T., Proestou D., Bik H., Simpson S. L., Portis L. M., Cantwell M. G., Baguley J. G., Burgess R. M., Pelletier M. M., Perron M., Gunsch C., Matthews R. A. A molecular-based approach for examining responses of eukaryotes in microcosms to contaminant-spiked estuarine sediments. (англ.) // Environmental Toxicology And Chemistry. — 2014. — February (vol. 33, no. 2). — P. 359—369. — doi:10.1002/etc.2450. — PMID 24399368.
- Адрианов, Малахов, 1994, с. 157—160.
- Класс Kinorhyncha (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 6 июля 2017)
- Рупперт, Фокс, Барнс, 2008, с. 56.
- Отряд Cyclorhagida (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 8 декабря 2018)
- Вестхайде, Ригер, 2008, с. 756.
- Отряд Homalorhagida (англ.) в Мировом реестре морских видов (World Register of Marine Species). (Дата обращения: 8 декабря 2018)
- Yamasaki H., Fujimoto S., Miyazaki K. Phylogenetic position of Loricifera inferred from nearly complete 18S and 28S rRNA gene sequences. (англ.) // Zoological Letters. — 2015. — Vol. 1. — P. 18—18. — doi:10.1186/s40851-015-0017-0. — PMID 26605063.
- Sørensen M. V., Dal Zotto M., Rho H. S., Herranz M., Sánchez N., Pardos F., Yamasaki H. Phylogeny of Kinorhyncha Based on Morphology and Two Molecular Loci. (англ.) // PloS One. — 2015. — Vol. 10, no. 7. — P. e0133440—0133440. — doi:10.1371/journal.pone.0133440. — PMID 26200115.
- Popova O. V., Mikhailov K. V., Nikitin M. A., Logacheva M. D., Penin A. A., Muntyan M. S., Kedrova O. S., Petrov N. B., Panchin Y. V., Aleoshin V. V. Mitochondrial Genomes of Kinorhyncha: trnM Duplication and New Gene Orders within Animals. (англ.) // PloS One. — 2016. — Vol. 11, no. 10. — P. e0165072—0165072. — doi:10.1371/journal.pone.0165072. — PMID 27755612.
- Robert Higgins. Zelinkaderidae, a New Family of Cyclorhagid Kinorhyncha.
- Yamasaki H., Fujimoto S. Two new species in the Echinoderes coulli group (Echinoderidae, Cyclorhagida, Kinorhyncha) from the Ryukyu Islands, Japan. (англ.) // ZooKeys. — 2014. — No. 382. — P. 27—52. — doi:10.3897/zookeys.382.6761. — PMID 24624018.
- Thomsen V. G., Rho H. S., Kim D., Sørensen M. V. A new species of Dracoderes (Kinorhyncha: Dracoderidae) from Korea provides further support for a dracoderid-homalorhagid relationship. (англ.) // Zootaxa. — 2013. — Vol. 3682. — P. 133—142. — PMID 25243279.
- Sørensen M. V., Rho H. S., Kim D. A new species of Condyloderes (Cyclorhagida, Kinorhyncha) from Korea. (англ.) // Zoological Science. — 2010. — March (vol. 27, no. 3). — P. 234—242. — doi:10.2108/zsj.27.234. — PMID 20192691.
Литература
- Адрианов А. В., Малахов В. В. Киноринхи. — М.: Наука, 1994. — 260 с. — ISBN 5-02-005737-1.
- Зоология беспозвоночных : в 2 т. / под ред. В. Вестхайде и Р. Ригера. — М. : Товарищество научных изданий КМК, 2008. — Т. 2: от артропод до иглокожих и хордовых. — iv + 513—935 + iii с. — ISBN 978-5-87317-495-9.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Зоология беспозвоночных: Функциональные и эволюционные аспекты : в 4 т. / под ред. А. А. Добровольского и А. И. Грановича. — М. : Издательский центр «Академия», 2008. — Т. 4. — 352 с. — ISBN 978-5-7695-3497-3.
- Iben Heiner Bang-Berthelsen, Reinhardt Kristensen, Andreas Maas, Birger Neuhaus. Handbook of Zoology. Nematomorpha, Priapulida, Kinorhyncha, Loricifera / Andreas Schmidt-Rhaesa. — Walter de Gruyter, 2013. — 391 p. — ISBN 978-3-11027253-6.
| Таксономия | |
|---|---|
| В библиографических каталогах |
