Мышечное сокращение
Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Это жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами.
Все виды произвольных движений — ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счёт скелетных мышц. Непроизвольные движения (кроме сокращения сердца) — перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря — обусловлены сокращением гладкой мускулатуры. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Типы

Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечают миофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1-2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновыми филаментами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновых филаментов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела[1].
Изометрическое сокращение
- Основная статья: Изометрическое сокращение
Изометрическое сокращение мышцы создаёт напряжение без изменения длины. Пример можно найти, когда мышцы руки и предплечья захватывают объект; суставы руки не двигаются, но мышцы генерируют достаточную силу, чтобы предотвратить падение объекта.
Изотоническое сокращение
- Основная статья: Изотоническое сокращение
При изотоническом сокращении напряжение в мышцах остаётся постоянным, несмотря на изменение длины мышц. Это происходит, когда сила сокращения мышц соответствует общей нагрузке на мышцы.
Концентрическое сокращение
При концентрическом сокращении мышечное напряжение является достаточным для преодоления нагрузки, а мышца укорачивается при сокращении. Это происходит, когда сила, создаваемая мышцей, превышает нагрузку, противодействующую её сокращению.
Эксцентрическое сокращение
- См. также: Эксцентрическая тренировка
При эксцентрическом сокращении напряжение недостаточно для преодоления внешней нагрузки на мышцы и мышечные волокна, удлиняются при их сокращении. Вместо того, чтобы тянуть сустав в направлении сокращения мышц, мышца действует так, чтобы замедлить сустав в конце движения или иным образом контролировать перемещение груза. Это может происходить невольно (например, при попытке переместить вес, слишком тяжёлый для подъёма мышцы) или добровольно (например, когда мышца «сглаживает» движение или сопротивляется гравитации, как, например, во время ходьбы вниз). В краткосрочной перспективе силовые тренировки с участием как эксцентрических, так и концентрических сокращений, по-видимому, увеличивают мышечную силу больше, чем тренировки только с концентрическими сокращениями. Однако вызванное физическими упражнениями повреждение мышц также больше при удлинении сокращений.
Эксцентрические сокращения в движении
Эксцентрические сокращения обычно возникают как тормозящее усилие в противоположность концентрическому сжатию, чтобы защитить суставы от повреждения. Во время практически любого обычного движения эксцентрические сокращения помогают сохранять плавность движений, но также могут замедлять быстрые движения, такие как удар или бросок. Часть тренировки для быстрых движений, таких как качки во время бейсбола, включает в себя уменьшение эксцентрического торможения, позволяющего развивать большую мощность во время движения.
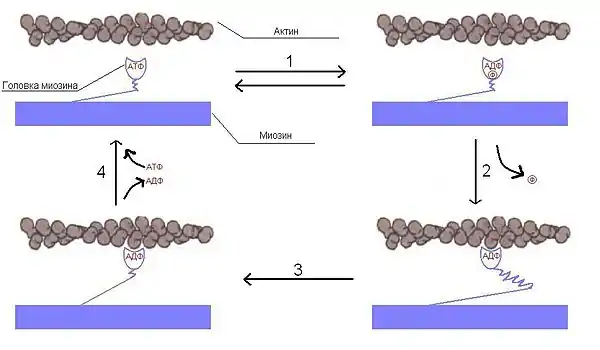
Скольжение миозина относительно актина
Головки миозина расщепляют АТФ и за счет высвобождающейся энергии меняют конформацию, скользя по актиновым филаментам. Цикл можно разделить на 4 стадии:
- Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата и остаётся связанной с ними. (Обратимый процесс — энергия, выделившаяся в результате гидролиза, запасается в изменённой конформации миозина).
- Головки слабо связываются со следующей субъединицей актина, фосфат отделяется, и это приводит к прочному связыванию головки миозина с актиновым филаментом. Эта реакция уже необратима.
- Головка претерпевает конформационное изменение, производящее подтягивание толстого филамента к Z-диску (или, что эквивалентно, свободных концов тонких филаментов друг к другу).
- Отделяется АДФ, за счёт этого головка отделяется от актинового филамента. Присоединяется новая молекула АТФ.
Далее цикл повторяется до уменьшения концентрации ионов Ca2+ или исчерпании запаса АТФ (в результате смерти клетки). Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновом филаменте много (около 500) молекул миозина и, следовательно, при сокращении цикл повторяется сотнями головок сразу, что и приводит к быстрому и сильному сокращению. Следует заметить, что миозин ведёт себя как фермент — актин-зависимая АТФаза. Так как каждое повторение цикла связано с гидролизом АТФ, а следовательно, с положительным изменением свободной энергии, то процесс однонаправленный. Миозин движется по актину только в сторону плюс-конца.
Источник энергии для сокращения
Для сокращения мышцы используется энергия гидролиза АТФ, но мышечная клетка имеет крайне эффективную систему регенерации запаса АТФ, так что в расслабленной и работающей мышце содержание АТФ примерно равно. Фермент фосфокреатинкиназа катализирует реакцию между АДФ и креатинфосфатом, продукты которой — АТФ и креатин. Креатинфосфат содержит больше запасённой энергии, чем АТФ. Благодаря этому механизму при вспышке активности в мышечной клетке падает содержание именно креатинфосфата, а количество универсального источника энергии — АТФ — не изменяется. Механизмы регенерации запаса АТФ могут различаться в зависимости от парциального давления кислорода в окружающих тканях (см. Анаэробные организмы).
Механизм регуляции
В основном в регуляции мышечной активности участвуют нейроны, но есть случаи, когда сокращением гладкой мускулатуры управляют и гормоны (например, адреналин и окситоцин). Сигнал о сокращении можно разделить на несколько этапов:
От клеточной мембраны до саркоплазматического ретикулума
Воздействие медиатора, выделившегося из мотонейрона, вызывает потенциал действия на клеточной мембране мышечной клетки, который передаётся далее с помощью специальных впячиваний мембраны, называемых Т-трубочками, которые отходят от мембраны внутрь клетки. От Т-трубочек сигнал передаётся саркоплазматическому ретикулуму — особому компартменту из уплощенных мембранных пузырьков (эндоплазматической сети мышечной клетки), окружающих каждую миофибриллу. Этот сигнал вызывает открытие Ca2+-каналов в мембране ретикулума. Обратно ионы Ca2+ попадают в ретикулум с помощью мембранных кальциевых насосов — Ca2+-АТФазы.
От выделения ионов Ca2+ до сокращения миофибрилл
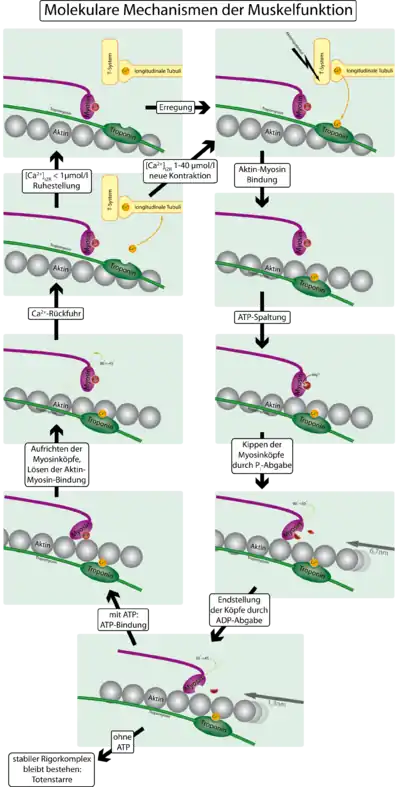
Для того, чтобы контролировать сокращение, к актиновому филаменту прикрепляется белок тропомиозин и комплекс из трёх белков — тропонин (субъединицы этого комплекса называются тропонинами T,I и C). Тропонин C — близкий гомолог другого белка, кальмодулина. Через каждые семь субъединиц актина расположен только один тропониновый комплекс. Связь актина с тропонином I перемещает тропомиозин в положение, мешающее связи миозина с актином. Тропонин C связывается с четырьмя ионами Ca2+ и ослабляет действие тропонина I на актин, и тропомиозин занимает положение, не препятствующее связи актина с миозином. Источником энергии для сокращения мышечных волокон служит АТФ. При связывании тропонина с ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. За счет ферментативной активности головок миозина гидролизуется АТФ, расположенный на головке миозина, что обеспечивает энергией изменение конформации головок и скольжение нитей. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. К миозиновой головке присоединяется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снизится до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
Основные белки миофибрилл
| Белок | Доля белка % | Его мол. масса, kDa | Его функция |
|---|---|---|---|
| Миозин | Главный компонент толстых филаментов. Образует связи с актином. Движется по актину за счёт гидролиза АТФ. | ||
| Актин | Главный компонент тонких филаментов. Во время сокращения мышцы по нему движется миозин. | ||
| Титин | Большой гибкий белок, образующий цепь для связывания миозина с Z-диском. | ||
| Тропонин | Комплекс из трёх белков, регулирующий сокращение при связывании с ионами Ca2+. | ||
| Тропомиозин | Связанный с актиновыми филаментами стержневидный белок, блокирующий движение миозина. | ||
| Небулин | Длинный нерастяжимый белок, связанный с Z-диском и идущий параллельно актиновым филаментам. |
Примечания
Литература
- Б. Альбертс, Д. Брей, Дж. Льюис, М. Рефф, К. Робертс, Дж. Уотсон, Молекулярная биология клетки — В 3-х т. — Пер. с англ. — Т.2. — М.: Мир, 1994. — 540 с.
- М. Б. Беркинблит, С. М. Глаголев, В. А. Фуралев, Общая биология — В 2-х ч. — Ч.1. — М.:МИРОС, 1999. — 224 с.: ил.
- Феррара Ж. Мышцы мышц // Наука и жизнь : журнал. — 1987. — № 1. — С. 95-96, V. — ISSN 0028-1263.
