Саркоплазматический ретикулум
Саркоплазмати́ческий рети́кулум (СР) — мембранная органелла мышечных клеток, схожая с эндоплазматическим ретикулумом (ЭПР) других клеток. Главная функция СР — запасание ионов кальция (Ca2+). Уровень кальция в клетке поддерживается относительно постоянным, причём концентрация кальция внутри клеток поддерживается в 100000 раз меньше, чем вне клеток. Поэтому небольшое повышение концентрации кальция в клетке может быть легко обнаружено и может сообщать о важных изменениях внутри клетки (кальций относят к числу так называемых вторичных посредников). Слишком высокий уровень кальция приводит к кальцификации некоторых внутриклеточных структур (например, митохондрий)[1], что приводит к гибели клетки. Поэтому в живой клетке уровень кальция жёстко контролируется, может при необходимости быть выделен в клетку и удалён из неё.
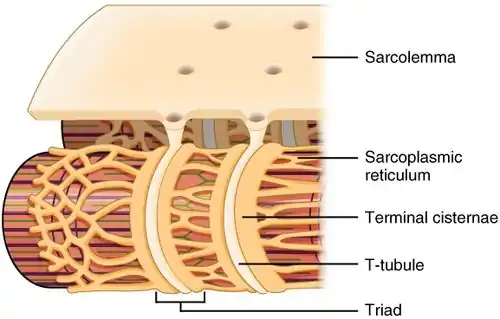
Структура
СР представляет собой сеть трубочек, тянущуюся по всем мышечным клеткам, обвиваясь, но не контактируя непосредственно, вокруг миофибрилл (сократительные единицы клеток). Мышечные клетки сердечной и скелетной мускулатуры содержат структуры, известные как T-трубочки, которые представляют собой впячивания клеточной мембраны, тянущиеся к центру клетки. T-трубочки тесно связаны с особыми элементами СР, известными как терминальные цистерны в случае сердечной мышцы и соединительный СР в случае скелетной мускулатуры (англ. junctional SR). Они разделены расстоянием около 12 нм. Это первичный сайт высвобождения кальция[2]. Продольные элементы СР представлены тонкими участками, которые соединяют между собой терминальные цистерны (соединительный СР). Именно в продольных участках кальциевые каналы, необходимые для его абсорбции, наиболее многочисленны[3].
Абсорбция кальция
Мембрана СР содержит ионные каналы (насосы), которые закачивают внутрь него Ca2+. Поскольку концентрация кальция в СР выше, чем в других частях клетки, ионы кальция не могут свободно поступать внутрь него: для этого нужны особые насосы, которые накачивают кальций внутрь с расходом энергии в виде АТФ. Такие насосы называются саркоплазматические Ca2+-АТФазы (англ. Sarcoplasmic reticulum Ca2+ ATPase, SERCA). Существует несколько разновидностей SERCA, причём SERCA 2a встречаются в основном в сердечной и скелетной мускулатуре[4].
SERCA состоит из 13 субъединиц, обозначаемых M1—M10, N, P и A. Кальций связывается с субъединицами M1—M10, которые находятся в мембране, в то время как АТФ связывается субъединицами N, P и А. Когда 2 иона кальция, а также одна молекула АТФ связываются с цитоплазматической стороной канала (то есть стороной, обращённой к цитоплазме), канал открывается, при этом АТФ превращается в АДФ, выделяя энергию. Высвободившаяся при этом фосфатная группа связывается с каналом, побуждая его изменить свою форму. Из-за этого изменения формы цитоплазматическая сторона канала открывается, и два иона кальция поступают в канал. Далее цитозольная сторона насоса закрывается, внутренняя открывается, высвобождая ионы кальция внутрь СР[5].
В сердечной мышце содержится белок, известный как фосфоламбан (PLB), который блокирует работу SERCA. Связываясь с каналом, PLB снижает его сродство к ионам кальция, мешая поступлению кальция внутрь СР. Если кальций не удаляется в СР из цитозоля, то мышца не может расслабиться, а значит, и снова сократиться. Однако адреналин и норадреналин могут мешать связыванию PLB с SERCA. Когда они связываются с β1-адренорецептором, расположенным в клеточной мембране, они запускают серию реакций, в конце концов приводящих к активации протеинкиназы А (PKA). PKA может фосфорилировать PLB, предотвращая его связывание с SERCA и запуская расслабление мышцы[6].
Запасание кальция
Внутри СР находится белок, известный как кальсеквестрин. Этот белок связывает около 50 ионов кальция, что снижает количество свободного кальция внутри СР. Благодаря этому в СР может запасаться больше Ca2+[7]. В основном кальсеквестрин находится внутри соединительного СР/терминальных цистерн, где он находится в близкой связи с кальциевыми каналами[8].
Высвобождение кальция
Высвобождение кальция из СР происходит в соединительном СР/терминальных цистернах через рианодиновые рецепторы (RyR) и также известно как кальциевая вспышка[9]. Существует три типа рианодиновых рецепторов: RyR1 (в скелетных мышцах), RyR2 (в сердечной мышце) и RyR3 (в мозге)[10]. В разных мышцах высвобождение кальция через рианодиновые рецепторы запускается по-разному. В сердце и в гладких мышцах электрический импульс (потенциал действия) запускает выход кальция в клетку через кальциевые каналы L-типа, расположенные в клеточной мембране (гладкие мышцы) или мембране T-трубочек (сердечная мышца). Эти ионы кальция связываются с рианодиновыми рецепторами и активируют их, в результате чего уровень кальция в клетке быстро повышается[11]. Кофеин, содержащийся в кофе, может связываться с рианодиновыми рецепторами и стимулировать их активность. Кофеин делает рианодиновые рецепторы более чувствительными к потенциалу действия (скелетная мускулатура) или кальцию (сердце и гладкие мышцы), в результате чего кальциевые вспышки происходят чаще[12].
Триадин и джунктин (англ. Triadin and Junctin) — это белки, находящиеся в мембране СР и связаны с RyR. Главная роль этих белков заключается в заякоривании кальсеквестрина к рианодиновым рецепторам. При нормальных (физиологических) уровнях кальция кальсеквестрин связывается с RyR, триадином и джунктином, что предотвращает открытие RyR[13]. Если концентрация кальция в СР становится слишком низкой, с кальсеквестрином связывается меньшее количество ионов кальция, и в этих условиях кальсеквестрин прочно связывается с триадином, джунктином и RyR. Если же кальция в СР слишком много, то он связывается с кальсеквестрином, и последний связан с триадином, джунктином и RyR менее прочно. Поэтому RyR могут открыться и высвободить кальций в клетку[14].
Кроме вышеописанного действия на фосфоламбан, которое приводит к расслаблению сердечной мышцы, РКА (а также другой фермент, известный как кальмодулин-киназа II) может фосфорилировать рианодиновые рецепторы. В фосфорилированном виде они более чувствительны к кальцию, поэтому открываются чаще и на большее время. Это приводит к выходу кальция из СР, усиливая темпы сокращения[15].
Механизм прекращения выхода кальция через RyR до конца не понятен. Некоторые учёные полагают, что это происходит при случайном закрытии RyR, или же рианодиновые рецепторы становятся неактивными после кальциевой вспышки. Другие учёные утверждают, что снижение уровня кальция в СР заставляет рецепторы закрыться[16].
Примечания
- Trump B. F., Berezesky I. K., Laiho K. U., Osornio A. R., Mergner W. J., Smith M. W. The role of calcium in cell injury. A review. (англ.) // Scanning electron microscopy. — 1980. — No. Pt 2. — P. 437—462. — PMID 6999604.
- Sommer J. R. The anatomy of the sarcoplasmic reticulum in vertebrate skeletal muscle: its implications for excitation contraction coupling. (англ.) // Zeitschrift fur Naturforschung. Section C, Biosciences. — 1982. — Vol. 37, no. 7-8. — P. 665—678. — PMID 7136180.
- Arai M., Matsui H., Periasamy M. Sarcoplasmic reticulum gene expression in cardiac hypertrophy and heart failure. (англ.) // Circulation research. — 1994. — Vol. 74, no. 4. — P. 555—564. — PMID 8137493.
- Periasamy M., Kalyanasundaram A. SERCA pump isoforms: their role in calcium transport and disease. (англ.) // Muscle & nerve. — 2007. — Vol. 35, no. 4. — P. 430—442. — doi:10.1002/mus.20745. — PMID 17286271.
- Kekenes-Huskey P. M., Metzger V. T., Grant B. J., Andrew Mc Cammon J. Calcium binding and allosteric signaling mechanisms for the sarcoplasmic reticulum Ca²+ ATPase. (англ.) // Protein science : a publication of the Protein Society. — 2012. — Vol. 21, no. 10. — P. 1429—1443. — doi:10.1002/pro.2129. — PMID 22821874.
- Akin B. L., Hurley T. D., Chen Z., Jones L. R. The structural basis for phospholamban inhibition of the calcium pump in sarcoplasmic reticulum. (англ.) // The Journal of biological chemistry. — 2013. — Vol. 288, no. 42. — P. 30181—30191. — doi:10.1074/jbc.M113.501585. — PMID 23996003.
- Beard N. A., Laver D. R., Dulhunty A. F. Calsequestrin and the calcium release channel of skeletal and cardiac muscle. (англ.) // Progress in biophysics and molecular biology. — 2004. — Vol. 85, no. 1. — P. 33—69. — doi:10.1016/j.pbiomolbio.2003.07.001. — PMID 15050380.
- MacLennan D. H., Wong P. T. S. Isolation of a Calcium-Sequestering Protein from Sarcoplasmic Reticulum // Proceedings of the National Academy of Sciences. — 1971. — 1 июня (т. 68, № 6). — С. 1231—1235. — ISSN 0027-8424. — doi:10.1073/pnas.68.6.1231.
- Cheng H., Lederer W. J., Cannell M. B. Calcium sparks: elementary events underlying excitation-contraction coupling in heart muscle. (англ.) // Science (New York, N.Y.). — 1993. — Vol. 262, no. 5134. — P. 740—744. — PMID 8235594.
- Lanner J. T., Georgiou D. K., Joshi A. D., Hamilton S. L. Ryanodine receptors: structure, expression, molecular details, and function in calcium release. (англ.) // Cold Spring Harbor perspectives in biology. — 2010. — Vol. 2, no. 11. — P. 003996. — doi:10.1101/cshperspect.a003996. — PMID 20961976.
- Cheng H., Lederer W. J. Calcium sparks. (англ.) // Physiological reviews. — 2008. — Vol. 88, no. 4. — P. 1491—1545. — doi:10.1152/physrev.00030.2007. — PMID 18923188.
- Sitsapesan R., Williams A. J. Mechanisms of caffeine activation of single calcium-release channels of sheep cardiac sarcoplasmic reticulum. (англ.) // The Journal of physiology. — 1990. — Vol. 423. — P. 425—439. — PMID 2167363.
- Zhang Lin, Kelley Jeff, Schmeisser Glen, Kobayashi Yvonne M., Jones Larry R. Complex Formation between Junctin, Triadin, Calsequestrin, and the Ryanodine Receptor // Journal of Biological Chemistry. — 1997. — 12 сентября (т. 272, № 37). — С. 23389—23397. — ISSN 0021-9258. — doi:10.1074/jbc.272.37.23389.
- Györke I., Hester N., Jones L. R., Györke S. The role of calsequestrin, triadin, and junctin in conferring cardiac ryanodine receptor responsiveness to luminal calcium. (англ.) // Biophysical journal. — 2004. — Vol. 86, no. 4. — P. 2121—2128. — doi:10.1016/S0006-3495(04)74271-X. — PMID 15041652.
- Bers D. M. Cardiac ryanodine receptor phosphorylation: target sites and functional consequences. (англ.) // The Biochemical journal. — 2006. — Vol. 396, no. 1. — P. e1—3. — doi:10.1042/BJ20060377. — PMID 16626281.
- Sham J. S. K., Song L.-S., Chen Y., Deng L.-H., Stern M. D., Lakatta E. G., Cheng H. Termination of Ca2+ release by a local inactivation of ryanodine receptors in cardiac myocytes // Proceedings of the National Academy of Sciences. — 1998. — 8 декабря (т. 95, № 25). — С. 15096—15101. — ISSN 0027-8424. — doi:10.1073/pnas.95.25.15096.