Конфуциусорнисы
Конфуциусо́рнисы[1] (лат. Confuciusornis, от лат. Confucius — латинизированная форма имени Конфуция и греч. ὄρνις — птица) — род примитивных ископаемых авиал (птиц в широком смысле) из нижнего мела Китая, живших 125—120 млн лет назад. Как и у современных птиц, клюв Confuciusornis был беззубым, однако, более близкие родственники современных птиц, — гесперорнисы и ихтиорнисы имели зубы, что говорит о независимой утрате зубов у конфуциусорнисов. Так или иначе, это самая ранняя из известных птиц, имевших клюв[2]. Назван в честь китайского философа Конфуция (551—479 до н. э.). Конфуциусорнис — одно из наиболее многочисленных позвоночных животных, обнаруженных в геологической формации Исянь (Yixian); было найдено несколько сотен полностью сочленённых экземпляров[3].
| † Конфуциусорнисы | ||||||
|---|---|---|---|---|---|---|
| ||||||
| Научная классификация | ||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Клада: Отряд: † Confuciusornithiformes Hou et al., 1995 Семейство: Род: † Конфуциусорнисы |
||||||
| Международное научное название | ||||||
| Confuciusornis Hou et al., 1995 | ||||||
| Синонимы | ||||||
|
||||||
| Виды | ||||||
| ||||||

.jpg.webp)
История изучения
_1.jpg.webp)
В ноябре 1993 года китайские палеонтологи Hou Lianhai и Hu Yoaming из института палеонтологии позвоночных и палеоантропологии (аббревиатируется как IVPP) посетили коллекционера окаменелостей Zhang He в его доме в Цзиньчжоу, где он показал палеонтологам образец ископаемой птицы, который он купил на местной барахолке. В декабре того же года L. Hou узнал о втором экземпляре, который был обнаружен фермером Yang Yushan. Оба экземпляра ископаемой птицы происходят из одной и той же местности в посёлке Шанъюань (Бэйпяо)[4][5]. В 1995 году L. Hou и его коллеги официально описали эти два экземпляра, а также третий, как типовых экземпляров нового рода (Confuciusornis) и вида ископаемой птицы — Confuciusornis sanctus. Первый полученный учёными экземпляр конфуциусорниса был обозначен как голотип и каталогизирован под номером IVPP V10918; данный типовой экземпляр включает в себя фрагментарный скелет с черепом и фрагментарной передней конечностью. Второй полученный экземпляр, обозначенный как паратип (IVPP V10895), включает целый таз и заднюю конечность. Другой обнаруженный паратип (IVPP V10919—10925) включает в себя фрагментарную заднюю конечность и отпечатки перьев по обеим сторонам большеберцовой кости[6]. Вскоре было отмечено, что два паратипа включают в себя только кóсти, отсутствующие у голотипа, что ставит принадлежность паратипов к виду C. sanctus под сомнение[7]. Лишь обнаружение множества хорошо сохранившихся экземпляров вскоре после этого подтвердило, что вышеупомянутые паратипы и голотип действительно представляют один вид[8]:16.
Виды и синонимы
С тех пор, как был описан C. sanctus, были официально выделены и описаны ещё пять видов. Как и в случае со многими другими ископаемыми родáми, отдельные виды трудно определить, поскольку различия между видами часто трудно отличить от внутривидовых изменений[9]:50. В случае с Confuciusornis, единственным общепринятым видом считается типовой вид C. sanctus.
- Confuciusornis chuonzhous был выделен L. Hou в 1997 году на основе морфологии образца IVVP V10919, первоначально являвшегося паратипом C. sanctus. Видовое название дано в честь Chuanzhou — устаревшего названия Бэйпяо[10]. В настоящее время C. chuonzhous признан синонимом C. sanctus[11].
- Counfuciusornis siniae, выделенный L. Hou в 1997 году в той же публикации, что и C. chuonzhous, был описан на основе морфологии образца IVPP V11308. Видовое название дано в честь жены Shikuan Liang — мадам Sun, отдавшей окаменелости в институт палеонтологии позвоночных и палеоантропологии[10]. В настоящее время большинство учёных признают C. siniae синонимом C. sanctus[11].
- Confuciusornis dui был выделен L. Hou и его коллегами в 1999 году. Видовое название дано в честь Du Wengya, отдавшего окаменелости в институт палеонтологии позвоночных и палеоантропологии. Голотип вида (IVPP V11553) представляет собой почти полный скелет взрослой особи, сохранивший в ископаемом состоянии пару удлинённых рулевых перьев и клюв с рамфотекой. Второй типовой экземпляр вида — паратип IVPP 11521, является фрагментарным: включает в себя некоторые позвонки, рёбра, хвост, грудную кость, таз и бедренную кость. Со слов Hou и его коллег, C. dui был меньше и грациознее других видов конфуциусорниса; образцы C. dui на 15 % меньше голотипа C. sanctus и на 30 % меньше крупных образцов C. sanctus. Кроме того, краниальные (передние) концы челюстей C. dui были острее, чем у C. sanctus; на нижней челюсти C. dui отсутствовал нижний киль (англ. underside keel); на первых пальцах передних конечностей C. dui когти не были увеличены, в отличие от C. sanctus; грудная кость C. dui была более удлинённой, чем у C. sanctus, и отличалась морфологическими особенностями, а цевка была короче пигостиля[12]. Статистический анализ, проведённый Jesús Marugán-Lobón и его коллегами в 2011 году, не выявил существенных различий между маленькими образцами конфуциусорниса, на основании чего можно предположить, что образцы C. dui являются особями C. sanctus, претерпевшими изменчивость. Тем не менее, авторы этого анализа не смогли повторно определить местонахождение голотипа C. dui, который, возможно, был утерян, в связи с чем учёным пришлось изучать слепок голотипа. Для подтверждения валидности вида требуется повторное изучение образцов C. dui[13].
- Confuciusornis feducciai был назван в 2009 году Zhang Fucheng и его коллегами в честь выдающегося палеоорнитолога Alan Feduccia. Голотип вида (D2454) был найден в местности Sihetun и в данный момент хранится в Дальянском музее природы (англ. Dalian Natural Museum). Со слов Zhang и его коллег, C. feducciai отличался от прочих видов конфуциусорниса более крупными размерами, иными пропорциями костей и рядом других уникальных морфологических особенностей:
- передние конечности C. feducciai были на 15 % длиннее задних, тогда как у C. sanctus они были одинаковой длины;
- в дорсальной (верхней) части плечевой кости C. feducciai отсутствовало большое отверстие, характерное для других видов конфуциусорниса;
- первые фаланги пальцев были стройными.
Прочие отличительные особенности вида заключаются в V-образной вилочке, широкой грудной кости и седалищной кости, которая долго сравнивалась с лобковой костью[14]. В 2011 году Jesús Marugán-Lobón и его коллеги заявили, что установление C. feducciai как отдельного вида спорно; несмотря на то, что в левой плечевой кости голотипа C. feducciai отсутствовало характерное для рода Confuciusornis отверстие, оно явно присутствовало в правой плечевой кости данного типового экземпляра. Кроме того, статистический анализ показал, что образцы C. feducciai могут являться особями C. sanctus, претерпевшими изменчивость. В связи с этим авторы исследования предположили, что C. feducciai является младшим синонимом C. sanctus[13].
- Confuciusornis jianchangensis был выделен в 2010 году Li Li и его коллегами на основе морфологии образца PMOL-AB00114, найденного в Toudaoyingzi. В отличие от других видов конфуциусорниса, C. jianchangensis обитал в геологической формации Цзюфотан (Jiufotang)[15].
В 2002 году L. Hou описал род Jinzhouornis, однако, в 2018 году Luis M. Chiappe, Wang и их коллеги пришли к выводу, что на основании морфометрии и осмотра образцов конфуциусорниса, этот род должен быть признан младшим синонимом рода Confuciusornis[16][17].
Систематика
Результаты кладистического анализа показывают, что конфуциусорнис — одна из наиболее примитивных пигостилевых птиц. Его череп был более примитивен, чем у археоптерикса, однако, это — древнейшая из известных птиц, у которых утрачен длинный рептильный хвост, вместо которого хвостовые позвонки срастаются, образуя короткий пигостиль[18]. В одном из спорных исследований утверждается, что конфуциусорнис мог являться более родственным микрорапторам и прочим дромеозаврид, чем археоптериксу, однако, научные методы этого исследования были подвергнуты критике на методологических основаниях[19].
| Paraves |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма по данным Xu и др., 2011[20]:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Настоящую стандартную интерпретацию филогенетической позиции конфуциусорниса может отражать приведённая ниже кладограмма:
| Aves |
| ||||||||||||||||||||||||||||||||||||
Близкий родственник конфуциусорниса, Changchengornis hengdaoziensis, также обитал в геологической формации Исянь (Yixian). Подобно конфуциусорнису, хвост Changchengornis нёс пару удлинённых рулевых перьев, похожих на таковых у некоторых, более продвинутых энанциорнисовых птиц. Правда, подвижные рулевые перья, расположенные «веером», появились лишь у представителей клады Euornithes и, возможно, независимо появились у энанциорнисовой птицы Shanweiniao[21][22].
Описание
Размеры
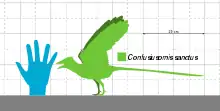
Конфуциусорнис был размером с современного голубя; его длина составляла 50 см[23] (из них примерно 30 см занимали рулевые перья), тогда как размах крыльев составлял 70 см. Вес конфуциусорниса, по оценкам разных авторов, составлял до 1,5 кг[24], или менее 0,2 кг[25]. C. feducciai являлся крупнейшим видом конфуциусорниса: экземпляры этого вида примерно на 30 % длиннее, чем средние экземпляры C. sanctus[26].
Отличительные особенности
Ископаемые остатки конфуциусорниса демонстрируют, что у него была необычайно большая плечевая кость. Грудина являлась относительно большой и имела невысокий киль, повышавшийся к каудальному (заднему) концу. Лопатка была неподвижно срощена с коракоидом и могла служить опорой для прикрепления летательных мышц. Плечевое сочленение было обращено вбок, а не повёрнуто слегка вверх, как у современных птиц; это указывает на то, что конфуциусорнис был неспособен высоко поднимать крылья над спиной. Подобно археоптериксу, конфуциусорнис был неспособен к машущему полёту (Phil Senter, 2006), однако, своеобразное строение плечевого пояса указывает на то, что он мог использовать и другую, не свойственную современным птицам технику полёта.
Конфуциусорнис обладал рядом более продвинутых черт, чем археоптерикс: например, скелет хвоста у него был укорочен и был развит пигостиль (косточка, сформировавшаяся из укороченных сросшихся хвостовых позвонков), но в то же время многие особенности строения этой птицы являлись более примитивными, чем у современных птиц: у него сохранились когти на пальцах крыльев, примитивное строение затылочной части черепа и относительно небольшая грудина[27].
Перья и мягкие ткани
Перья на крыльях конфуциусорниса являлись длинными, по строению и расположению аналогичными таковым у современных птиц. Маховые перья 1-го порядка экземпляра весом 500 г достигают 20,7 см в длину. Пять длиннейших маховых перьев 1-го порядка конфуциусорниса были в 3,5 раза длиннее кисти и относительно длиннее, чем у всех современных птиц, но в то же время маховые перья 2-го порядка на предплечье являлись относительно короткими[24]. Крайние маховые перья 1-го порядка были намного короче крайних маховых 2-го порядка, образуя относительно округлое, широкое крыло. Форма крыла конфуциусорниса была не похожа на какую-либо форму крыла среди современных птиц[28]. Маховые перья 1-го порядка являлись в разной степени асимметричными, особенно крайние маховые[8]. Неизвестно, были ли у конфуциусорниса маховые перья 3-го порядка. У некоторых экземпляров в ископаемом состоянии сохранились кроющие и контурные перья[8].
В отличие от некоторых более продвинутых птиц, у конфуциусорниса отсутствовало крылышко (лат. alula). У современных птиц крылышко сформировано перьями, прикреплёнными к первому пальцу крыла, но у конфуциусорниса этот палец не был покрыт перьями и имел коготь, а все три пальца крыла не срастались[8]. По мнению Dieter Stefan Peters, третий палец крыла конфуциусорниса мог формировать аналог крылышка на тыльной стороне крыла, функционировавший как предкрылок Крюгера[29]. Несмотря на относительно продвинутые и длинные маховые перья, на предплечьях образцов конфуциусорниса отсутствуют какие-либо признаки наличия papillae ulnares и костных точек прикрепления перьев[8].
Многие экземпляры в ископаемом состоянии сохранили пару узких лентовидных рулевых перьев с коротким опахалом на конце, которые были длиннее всего тела конфуциусорниса. У современных птиц к пигостилю «веером» прикрепляются основания рулевых перьев, в то время как у конфуциусорниса к пигостилю прикреплялись основания не аэродинамичных перьев, по строению не имеющих аналогов среди современных птиц[8]. Данные перья были устроены очень примитивным образом: в отличие от таковых у ныне живущих птиц, рулевые перья конфуциусорниса не состояли из стержня и бородок по большей части своей длины. Скорее, по большей части своей длины данные перья представляли из себя подобный ленте слой, приблизительно шесть миллиметров шириной. Только в последней четверти перьев, по направлению к закруглённому кончику, они становились дифференцированными в центральный стержень со сцепленными бородками. Однако у некоторых экземпляров конфуциусорниса подобных удлинённых рулевых перьев нет, возможно, из-за полового диморфизма (см. раздел «Палеобиология», а именно «Рулевые перья»).
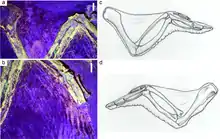
Лазерная флуоресценция двух образцов конфуциусорниса выявила прочие особенности строения мягких тканей животного. Подобно современным птицам, пропатагиум (англ. propatagium) конфуциусорниса был большим, вероятно, относительно толстым и располагался между плечом и запястьем; размер постпатагиума (англ. postpatagium) так же был схож с таковым у ныне живущих птиц. Сетчатые чешуйки (англ. reticulate scales) покрывали вентральную (нижнюю) часть стопы́, тогда как фаланги и плюсневые кости поддерживали большие мясистые подушечки, но в то же время межпальцевые подушечки являлись либо маленькими, либо вовсе отсутствовали[28].
Окрас оперения

В начале 2010 года, изучая окаменелости конфуциусорниса при помощи электронного микроскопа, группа учёных во главе с Zhang Fucheng обнаружили меланосомы в ископаемом образце IVPP V13171. Авторы исследования сообщили о наличии меланосом двух типов: эумеланосом (eumelanosomes) и феомеланосом (pheomelanosomes). Наличие данных меланосом указывает на то, что оперение конфуциусорниса могло совмещать в себе оттенки серого, красного/коричневого и чёрного цветов, т. е. по окрасу конфуциусорнис мог напоминать зяблика. Если результаты исследования были верны, то это был первый случай, когда в окаменелостях ранних птиц сохранились феомеланосомы[30]. Тем не менее, вторая группа учёных не смогла обнаружить зарегистрированные следы феомеланосом. Повторный осмотр образца в 2011 году выявил корреляцию между присутствием определённых металлов, таких как медь, и сохранившимися меланинами. Принимая во внимание сочетание окаменелых оттисков меланосом и присутствием металла в перьях, вторая группа учёных реконструировала конфуциусорниса с тёмными перьями туловища и тёмными перьями на верхней части крыла, но не обнаружила ни меланосом, ни металлов в большинстве перьев крыла. Учёные предположили, что перья на крыльях конфуциусорниса были белыми или, возможно, были окрашены каротиноидными пигментами (от жёлтого, до чёрно-коричневого цвета). Со слов авторов исследования, длинные рулевые перья конфуциусорниса так же были тёмного цвета по всей длине[31].
Исследование образца CUGB P1401, проведённое в 2018 году, показало наличие больших пятен на крыльях, шее и холке конфуциусорниса[32].
 Confuciusornis sanctus. Confuciusornis sanctus в музее Cosmocaixa, Барселона
Confuciusornis sanctus. Confuciusornis sanctus в музее Cosmocaixa, Барселона Confuciusornis sanctus. Confuciusornis sanctus на экспозиции Китайского музея палеозоологии
Confuciusornis sanctus. Confuciusornis sanctus на экспозиции Китайского музея палеозоологии Экземпляр Confuciusornis на экспозиции Китайского музея палеозоологии
Экземпляр Confuciusornis на экспозиции Китайского музея палеозоологии Confuciusornis sanctus. Confuciusornis sanctus на экспозиции в Гонконгском естественнонаучном музее
Confuciusornis sanctus. Confuciusornis sanctus на экспозиции в Гонконгском естественнонаучном музее
Палеобиология
Большие, мясистые подушечки на пальцах, маленькие межпальцевые подушечки, наличие лишь сетчатых чешуек (англ. reticulate scales) на вентральной (нижней) стороне стопы (что повышает гибкость) и изогнутые цепкие когти стопы конфуциусорниса — все перечисленные черты характерны для современных древесных птиц, на основании чего можно предположить, что конфуциусорнис вёл древесный образ жизни[28].
Сравнение между склеротикальным кольцом (лат. anulus ossicularis sclerae), позволяющим поддерживать постоянную форму глаза с таковым у современных птиц и рептилий показывает, что конфуциусорнис мог вести дневной образ жизни[33].
Полёт

Традиционно считается, что конфуциусорнис являлся отличным летуном, принимая во внимание его необычайно длинные крылья с крайне асимметричными перьями. Другие адаптации для хорошего полёта включают в себя: изгиб запястья, укороченный скелет хвоста, окостеневшую грудину с центральным килем, коракоид, большой дельтопекторальный гребень, сильное предплечье и увеличенную пястную кость[34]. Киль и большой дельтопекторальный гребень (способствующие более мощному подъёму в воздух) у современных птиц являются адаптациями к машущему полёту, что указывает на то, что, возможно, конфуциусорнис был способен осуществлять машущий полёт. Тем не менее, в связи с неспособностью вращать крыльями сзади тела, конфуциусорнис, возможно, имел и другую, не свойственную современным птицам технику полёта, а относительно меньший, чем у ныне живущих птиц киль указывает на то, что конфуциусорнис, вероятно, был неспособен к полёту в течение длительных промежутков времени[28].
Против этого было выдвинуто несколько противоположных утверждений о полёте конфуциусорниса. Первая рассматриваемая проблема — достижение крутой траектории полёта в связи с ограниченной амплитудой крыльев. В интерпретации Phil Senter (2006) положение плечевых суставов конфуциусорниса исключает возможность осуществлять полный взмах крыльями. Менее радикальным утверждением является то, что в связи с отсутствием килевой грудины (англ. keeled sternum) и наличием большого количества акрокоракоидов (англ. acrocoracoids), малая грудная мышца конфуциусорниса была неспособна приводить в движение Musculus supracoracoideus, поднимавшего плечевую кость при помощи сухожилия, проходившего через foramen triosseum. Это, наряду с ограниченной подвижностью крыльев вверх, обусловленной особым положением плечевых суставов, затруднило бы повышение высоты во время полёта, иными словами, конфуциусорнис был неспособен высоко летать. В связи с этим некоторые учёные предположили, что конфуциусорнис использовал свои когти больших пальцев для лазания по стволам деревьев. Ларри Дин Мартин высказал предположение, что конфуциусорнис был способен распрямить туловище на стволе дерева почти вертикально, подобно современным беличьим (лат. Sciuridae)[27]. Признавая, что лазание конфуциусорниса по деревьям было вероятным, Daniel Hembree отметил, что крестец, по-видимому, не был поднят более чем на 25° относительно бедра в вертикальном положении птицы, что было обусловлено расположением противовертела (англ. antitrochanter) в тазобедренном суставе[35]. Dieter S. Peters, со своей стороны, посчитал маловероятным, чтобы конфуциусорнис взбирался на стволы деревьев, поскольку поворот когтистых больших пальцев вытянул бы длинное крыло вперёд, — прямо на пути препятствующих веток. По мнению этого исследователя, конфуциусорнис мог осуществлять машущий полёт, однако, специализировался на парящем[29].

Вторая проблема — прочность перьев. В 2010 году Robert L. Nudds и Gareth J. Dyke опубликовали исследование, в котором они заявили, что и у конфуциусорниса, и у археоптерикса стержни маховых перьев являлись слишком тонкими и непрочными, чтобы оставаться жёсткими в процессе взмаха крыльями, необходимого для машущего полёта. Кроме того, Nudds и Dyke утверждали, что конфуциусорнис специализировался на планирующем полёте, что коррелирует с необычными адаптациями, наблюдаемыми в костях предплечья и, скорее всего, конфуциусорнис пользовался своими крыльями как «парашютом» — уменьшая скорость падения, в том случае, если бы конфуциусорнис упал с дерева[24]. Однако Грегори Скотт Пол не согласился с данным исследованием. Он утверждал, что Nudds и Dyke переоценили вес конфуциусорниса, и что более точные оценки веса предполагают совершение машущего полёта даже с тонкими стержнями маховых перьев. Nudds и Dyke оценили вес конфуциусорниса в 1,5 кг, в то время как Пол утверждал, что более разумная оценка массы тела составляет всего около 180 г. Пол также отметил, что образцы конфуциусорниса обычно встречаются в виде крупных скоплений в донных отложениях озера, тогда как ископаемые остатки животных, осуществлявших планирующий полёт, крайне редко встречаются в больших количествах на дне водоёмов. Скорее, скопления ископаемых остатков указывают на то, что конфуциусорнис в многочисленных стаях пересекал озеро — водную средý, характерную для животных, способных к машущему полёту[25]. Ряд других исследователей так же поставили под сомнение правильность измерений стержней маховых перьев, заявив, что исследованные Nudds и Dyke образцы в действительности имели диаметр стержней маховых перьев 2,1—2,3 мм (0,083—0,091 дюйма), а не 1,2 мм (0,047 дюйма), как сообщили Nudds и Dyke[36]. Со своей стороны, Nudds и Dyke возразили, что, помимо аспекта массы тела, такой большой диаметр стержней маховых перьев сам по себе мог бы обеспечить машущий полёт, однако, они допускали наличие двух видов конфуциусорниса в ископаемом материале Китая с различным диаметром стержней маховых перьев[37].
В 2016 году Amanda R. Falk и её коллеги привели доводы в пользу хороших лётных возможностей конфуциусорниса, принимая во внимание данные лазерной флуоресценции двух образцов, сохранивших мягкие ткани. Авторы исследования выявили, что вопреки утверждениям Nudds и Dyke, стержни маховых перьев конфуциусорниса являлись относительно крепкими, с максимальным диаметром более 1,5 мм (0,059 дюйма). Крылья исследованных образцов учёные сочли аналогичными либо крыльям современных птиц, живущих в густых лесах, либо крыльям современных птиц, специализирующихся на планирующем полёте; первый вывод коррелирует с тем, что среда обитания кофуциусорниса была плотно покрыта лесами[38], в связи с чем конфуциусорнису требовалась высокая манёвренность во время полёта. Кроме того, со слов Falk и её коллег, ярко выраженный пропатагиум (англ. propatagium) обеспечил бы крылья конфуциусорниса большой подъёмной силой, тогда как большой постпатагиум (англ. postpatagium) обеспечил бы большую площать прикрепления очина перьев, что позволило бы сохранить ровное положение крыльев во время полёта. Все вышеперечисленные доводы в совокупности убедительно свидетельствуют о том, что конфуциусорнис был способен к механическому (с использованием движений крыльев) полёту, хотя бы на короткие промежутки времени[28].
Рулевые перья

Многие экземпляры конфуциусорниса в ископаемом состоянии сохранили пару длинных рулевых перьев, по длине похожих на те, что присутствуют у некоторых современных райских птиц (лат. Paradisaeidae)[39]. Образцы, у которых отсутствуют рулевые перья, сохранили контурные перья, следовательно, причина отсутствия рулевых перьев не заключается в их плохой сохранности[8]. В 1998 году Ларри Мартин и его коллеги заявили, что рулевые перья присутствовали примерно у 5—10 % образцов, известных на тот период времени[27]. Статистический анализ, проведённый в 2011 году Jesús Marugán-Lobón и его коллегами, показал, что из 130 образцов 18 % имели рулевые перья, тогда как 28 % — нет, но в то же время у оставшихся 54 % сохранность была недостаточной для определения отсутствия или присутствия данных перьев[13]. Биологический смысл этого пáттерна расценивался неоднозначно. Ларри Мартин и его коллеги предположили, что паттерн может отражать половой диморфизм, т. е. длинные рулевые перья присутствовали только у самцов, и использовались в брачных демонстрациях[27]. Эта интерпретация поддерживалась большинством последующих исследований[40]. В 1999 году Luis Chiappe и его коллеги утверждали, что половой диморфизм — не единственное, но наиболее разумное объяснение наличия/отсутствия рулевых перьев, отмечая, что у современных птиц длина перьев часто варьируется в зависимости от пола[8].

Противоречие возникло из наблюдения, что известные образцы конфуциусорниса можно разделить на большую и небольшую группы, однако, данное мультимодальное распределение не связано с наличием рулевых перьев. В 2008 году Luis Chiappe и его коллеги утверждали, что подобное распределение образцов по размеру можно объяснить «динозавроподобным» способом роста (см. раздел «Онтогенез»), и что половой диморфизм является наиболее вероятным объяснением наличия/отсутствия рулевых перьев[16]. Со своей стороны, в 2009 году Winfried Peters и Dieter Peters возразили, что у обоих полов конфуциусорниса, вероятно, были длинные рулевые перья, как это имеет место у большинства современных птиц, обладающих подобными по длине рулевыми перьями. Эти исследователи заявили, что особи одного из полов могли иметь более крупные размеры тела, чем особи противоположного пола. Кроме того, авторы исследования заключили, что мультимодальное распределение экземпляров конфуциусорниса по размеру и наличию рулевых перьев делает конфуциусорниса похожим на водяного фазанчика (лат. Hydrophasianus chirurgus) — современную птицу, у которой самки крупнее самцов, а у взрослых особей обоих полов в период размножения вырастают длинные рулевые перья. Однако конфуциусорнис отличается от водяного фазанчика тем, что рулевые перья присутствуют у экземпляров самых различных размеров, даже у некоторых из самых маленьких. Со слов Peters и Peters, этот факт указывает на то, что конфуциусорнисы могли не использовать рулевые перья в брачных демонстрациях[41].
Различными авторами было предложено несколько альтернативных гипотез, объясняющих частое отсутствие рулевых перьев. В 1999 году Luis Chiappe и его коллеги обсуждали вероятность того, что экземпляры конфуциусорниса умерли во время линьки — на момент смерти они сбросили длинные рулевые перья. Хотя прямых доказательств линьки у мезозойских птиц не найдено, отсутствие истирания перьев у образцов конфуциусорниса указывает на то, что оперение у них периодически обновлялось. Подобно современным птицам, самцы и самки конфуциусорниса могли линять в разное время и, возможно, поэтому в одной группе образцов присутствовали как экземпляры с рулевыми перьями, так и без них[8]. Winfried Peters и Dieter Peters, со своей стороны, предположили, что конфуциусорнисы сбрасывали рулевые перья как средство защиты, подобно некоторым современным птицам. Такое выпадение рулевых перьев могло быть обусловлено стрессом, вызванным извержением вулканов, которое и погребло некоторых конфуциусорнисов под землёй[41]. Со своей стороны, в 2011 году Jesús Marugán-Lobón и его коллеги заявили, что необходимо учитывать даже наличие двух видов конфуциусорниса: одного с длинными рулевыми перьями, другого — без. Однако в настоящее время эта возможность будет необоснованной, поскольку другие морфологические различия между этими двумя возможными видами не выявлены[13].
Размножение
В 2010 году Gary W. Kaiser сообщил об экземпляре конфуциусорниса, под правой задней конечностью которого сохранилось первое возможное яйцо, относящееся к этому рóду. У самогó экземпляра отсутствуют длинные рулевые перья, что указывает на то, что при жизни он мог являться самкой. Яйцо могло выпасть из тела после смерти предполагаемой самки, хотя не исключено, что яйцо принадлежало другой самке, и оказалось под задней конечностью вышеупомянутого скелета случайно. Найденное яйцо округлой формы и имеет диаметр 17 мм, что немного меньше диаметра черепа образца; со слов Kaiser, яйцо точно бы прошло через тазовый канал[комм. 1] конфуциусорниса, что подтверждает принадлежность яйца к данному роду[42]:244—245[43]. У ящеротазовых динозавров, включая ранних мезозойских птиц, таз являлся закрытым, тогда как ширина тазового канала являлась относительно небольшой из-за соединения вентральных (нижних) частей лобковых костей (т. н. лобковый симфиз), в результате чего образовывалось V-образное отверстие, через которое откладывались яйца. У ныне живущих птиц таз уже открытый (лобковый симфиз отсутствует), в связи с чем современные птицы обрели способность откладывать относительно крупные яйца. В 2010 году Gareth Dyke и Gary Kaiser выявили, что яичная скорлупа у яйца конфуциусорниса пропускала воздух меньше, чем можно было бы ожидать от яйца современной птицы таких же размеров[43]. В своей книге 2016 года Luis Chiappe и Meng Qinjin утверждали, что V-образное отверстие лобковых костей образца конфуциусорниса крупных размеров (DNHM-D 2454) указывает на максимально возможный диаметр яйца (которое смогло бы пройти через тазовый канал птицы) 23 мм (0,91 дюйма). Среди современных птиц относительно более крупные яйца обычно встречаются у видов, чьи птенцы продолжительное время своего роста неспособны выжить без заботы родителей (птенцовые птицы), в то время как относительно менее крупные яйца обычно встречаются у видов, чьи птенцы развиваются быстрее и становятся независимыми раньше (выводковые птицы). Поскольку обнаруженное яйцо конфуциусорниса на 30 % меньше, чем можно было бы ожидать от современной птенцовой птицы, вполне вероятно, что конфуциусорнис являлся выводковым[9]. В 2018 году Charles Deeming и Gerald Mayr измеряли размер тазового канала различных ранних мезозойских птиц, включая конфуциусорниса. Исследователи пришли к выводу, что яйца были небольшими по отношению к массе тела для всех ранних мезозойских птиц в целом. Кроме того, эти исследователи заключили, что инкубация яиц, которую демонстрирует большинство современных птиц (высиживание яиц) была физически невозможна для нептичьих динозавров и ранних мезозойских птиц, включая конфуциусорниса, поскольку эти животные являлись слишком тяжёлыми по отношению к их относительно небольшим яйцам[44]. В 2010 году Gary Kaiser утверждал, что конфуциусорнис, вероятно, не вил гнёзда из веток, травы, листьев, перьев, пуха, грязи и подобных материалов, вместо чего мог гнездиться в дуплах деревьев, и что относительно небольшой размер единственного яйца указывает на большой размер кладки[42]:246. Со своей стороны, в 2016 году David J. Varricchio и Frankie D. Jackson возразили, что гнездование над землёй возникло в эволюции авиал гораздо позже и лишь у представителей веерохвостых птиц, и что мезозойские птицы закапывали свои яйца в земле — полностью или частично, как это наблюдается у других вымерших паравесов[45].
Онтогенез
Для развития любого существа в онтогенезе характерно увеличение массы тела, т. е. наличие роста — количественного признака, характеризующегося увеличением количества клеток и накоплением массы внутриклеточных образований, линейных размеров тела. Рост вымерших животных можно реконструировать, учитывая внутреннюю структуру кости́. Первое подобного рода исследование кости конфуциусорниса было проведено в 1998 году Fucheng Zhang и его коллегами, которые изучили поперечное сечение бедренной кости при помощи сканирующей электронной микроскопии. Со слов авторов исследования, бедренная кость была хорошо васкуляризирована (содержала множество кровеносных сосудов) и демонстрировала лишь одну линию задержки роста, в связи с чем учёные пришли к выводу, что рост конфуциусорниса являлся быстрым и непрерывным, как у современных птиц, и что конфуциусорнис являлся теплокровным[46]. Zhang и его коллеги подтвердили свои выводы в следующей статье, заявив, что внутренняя структура бедренной кости конфуциусорниса не похожа на таковую у современных аллигаторовых (лат. Alligatoridae), и похожа на таковую у оперённого нептичьего динозавра бэйпяозавра[47]. Однако эти исследователи предположили, что у конфуциусорниса и современных птиц истинная гомойотермия возникла независимо, то есть её не было у ближайшего общего предка всех авиал[46]. Данная гипотеза согласуется с более ранней работой Anusuya Chinsamy и её коллег, описавших отчётливые линии медленного роста и низкой васкуляризации кости у других мезозойских птиц. По мнению этих исследователей, медленный рост мезозойских птиц указывает на их относительно низкую (по сравнению с современными птицами) скорость метаболизма. Авторы исследования пришли к выводу, что истинная гомойотермия возникла поздно в эволюционной линии авиал, ведущей к современным птицам[48]. Однако данное утверждение было оспорено последующими исследованиями, в которых было указано, что медленный рост кости не указывает на априори низкую скорость метаболизма животного, а в случае мезозойских птиц, — их относительно медленные темпы роста, скорее, были обусловлены эволюционным уменьшением размеров тела, которое характеризует раннюю эволюцию авиал[49][50]. Со своей стороны, в 2003 году Armand de Ricqlès и его коллеги провели более подробное гистологическое исследование, основанное на восьмидесяти тонких срезах кости, взятых у экземпляра конфуциусорниса, на момент смерти достигшего половой зрелости. Данное исследование подтвердило высокие темпы роста конфуциусорниса, предложенные Fucheng Zhang и его коллегами. Со слов Armand de Ricqlès и его коллег, быстрорастущая фиброламеллярная костная ткань конфуциусорниса похожа на таковую у нептичьих динозавров, и выбранная для исследования особь, вероятно, достигла размеров взрослой особи гораздо раньше, чем за 20 недель. Авторы исследования заключили, что небольшой размер тела конфуциусорниса был достигнут не за счёт сокращения всей фазы роста, а за счёт сокращения стремительной фазы роста. Скорость роста, оценённая для конфуциусорниса, всё ещё ниже, чем чрезвычайно быстрый рост, характерный для современных птиц (6—8 недель), на основании чего можно предположить, что скорость роста веерохвостых птиц является их апоморфией[50].
В 2008 году Luis Chiappe и его коллеги провели статистический анализ на основе 106 образцов конфуциусорниса, с целью выявить, существует ли корреляция между размером тела образцов и наличием рулевых перьев. Данный анализ выявил бимодальное распределение образцов конфуциусорниса по двум размерным группам, однако, не было выявлено корреляции между размером тела и наличием рулевых перьев. В связи с этим авторы исследования пришли к выводу, что либо особи обоих полов конфуциусорниса были одинаковыми по размеру, либо у особей обоих полов были длинные рулевые перья. Первый вариант был признан учёными наиболее правдоподобным, в связи с чем причина бимодального распределения образцов по размеру тела оставалась неясной. Авторы исследования выдвинули гипотезу, что образцы конфуциусорниса отличаются по размеру в связи с тем, что менее крупные образцы представлены ювенильными (незрелыми) особями, в то время как более крупные образцы представлены половозрелыми особями, а редкость особей промежуточных (средних) размеров была обусловлена тем, что рост конфуциусорниса ускорялся непосредственно перед достижением половой зрелости; эта стремительная фаза роста являлась весьма быстрой, в связи с чем особей конфуциусорниса в вышеупомянутой фазе роста было довольно мало и, как следствие, после гибели бóльшая их часть не подверглась фоссилизации. Первоначально довольно медленный рост с последующим его ускорением незадолго до достижения половой зрелости указывает на «кривую роста»[комм. 2] S-образной формы, — подобную той, что предполагается большинством учёных для нептичьих динозавров. Подобный способ роста противоречит предыдущим гистологическим исследованиям, предполагавшим гораздо более «птичий» рост конфуциусорниса. Со слов Chiappe и его коллег, в качестве альтернативы наблюдаемое бимодальное распределение образцов конфуциусорниса по размеру можно объяснить тем, что образцы представляют разные виды, хотя не было выявлено никаких морфологических различий между образцами, указывающих на это. Другое альтернативное объяснение заключается в том, что у ювенильных и половозрелых особей была наиболее высокая смертность[16].
Со своей стороны, в том же году Winfried Peters и Dieter Peters подвергли критике «динозавроподобный» способ роста конфуциусорниса, предложенный Luis Chiappe и его коллегами, утверждая, что размер тела образцов меньшей размерной группы слишком велик, чтобы представлять ювенильных особей. Анализируя расширенный набор данных, Peters и Peters выявили третью размерную группу, предположительно состоявшую из ювенильных особей. Поскольку маловероятно, чтобы у конфуциусорниса было две стремительных фазы роста (этой особенности не было выявлено ни у одного известного амниота), авторы исследования пришли к выводу, что две большие размерные группы состоят из особей разных полов, а не из особей разного возраста, причём длинные рулевые перья были у особей обоих полов. Эта интерпретация согласуется с «птичьим» типом роста конфуциусорниса, который предполагали более ранние гистологические исследования. Более того, эта интерпретация также согласуется с современными птицами, у большинства из которых длинные рулевые перья присутствуют у особей обоих полов. Авторы исследования предположили, что отсутствие длинных рулевых перьев у многих экземпляров было обусловлено стрессом (вызвавшим выпадение перьев) незадолго до смерти животных[41].
Со своей стороны, в 2010 году в своём комментарии Luis Chiappe и его коллеги настояли на своих выводах, утверждая, что предполагаемый «птичий» рост конфуциусорниса маловероятен. Расчёты, проведённые Armand De Ricqlès в 2003 году, по которым рост конфуциусорниса был оценён длительностью менее 20 недель, были основаны на предположении, что диаметр исследованной кости конфуциусорниса увеличивался на 10 мкм в день, что является субъективным. Со слов Chiappe и его коллег, в действительности гистология выявляет наличие в кости различных типов ткани, растущих с разной скоростью, а также на фазы замедления роста, на что указывают линии задержки роста. Таким образом, согласно результатам двух исследований, рост конфуциусорниса длился гораздо дольше, чем у большинства современных птиц и, вероятно, длился несколько лет, как у современных киви (лат. Apteryx)[51][52]. Данный вывод говорит о том, что наблюдаемое бимодальное распределение образцов по размеру тела можно объяснить, допуская «динозавроподобный» способ роста конфуциусорниса[51][53]. Со своей стороны, в том же году Winfried Peters и Dieter Peters заявили, что Chiappe и его коллеги не прокомментировали их главный аргумент — разницу в размерах тела между меньшей размерной группой и предполагаемыми детёнышами, которая составляет десять раз[53]. В 2011 году Jesús Marugán-Lobón и его коллеги изучили взаимосвязь между наличием/отсутствием рулевых перьев и длиной костей конечностей, руководствуясь 130-ю образцами конфуциусорниса, подтверждая, что между размером тела и наличием/отсутствием рулевых перьев нет никакой корреляции; корреляция присутствует между рулевыми перьями и разными пропорциями передней и задней конечностей. Авторы исследования пришли к выводу, что значение наблюдаемого бимодального распределения как рулевых перьев, так и размеров тела остаётся спорным[13].
В 2008 году Chiappe и его коллеги заключили, что рост костей конечностей конфуциусорниса являлся почти изометрическим[комм. 3][16]. Однако данное утверждение было оспорено Peters и Peters в 2009 году, которые выявили, что кости крыльев, как правило, относительно длиннее у очень маленьких экземпляров конфуциусорниса (подобно современным курам), в связи с чем авторы исследования пришли к выводу, что рост костей конфуциусорниса являлся аллометрическим[41]. Со своей стороны, в своём комментарии в 2010 году Chiappe и его коллеги возразили, что вариации пропорций костей крыльев присутствуют у экземпляров самых различных размеров, и что аллометрический рост костей конфуциусорниса не был доказан исследованиями, проведёнными Peters и Peters[51].
Возможная медуллярная кость
В 2013 году Anusuya Chinsamy и её коллеги провели гистологическое исследование, в котором они сообщили, что обнаружили медуллярную кость в полостях трубчатых костей образца конфуциусорниса без рулевых перьев (DNHM-D1874), но не обнаружили медуллярную кость у трёх образцов с рулевыми перьями. В связи с этим авторы исследования предположили, что образцы конфуциусорниса без рулевых перьев при жизни были самками, тогда как образцы с рулевыми перьями — соответственно, самцами. Обнаруженная предполагаемая самка на момент смерти уже прошла стремительную фазу роста, хотя её размер всё ещё являлся значительно меньше максимального размера, достигнутого крупными экземплярами конфуциусорниса. Кроме того, со слов авторов исследования, можно выделить по крайней мере две линии задержки роста, указывающие на постепенный рост особи в течение нескольких лет; предполагаемая самка, скорее всего, умерла в возрасте трёх лет. Исследователи отметили, что длинные рулевые перья присутствуют у мелких экземпляров конфуциусорниса, размер тела которых составляет всего 23 % от размера тела крупных. В связи с этим фактом авторы исследования пришли к выводу, что половая зрелость предполагаемых самцов, скорее всего, наступила задолго до того, как они достигли своих окончательных размеров, что кардинально отличает их от современных птиц, у которых половая зрелость наступает тогда, когда они уже перестают расти[40]. Со своей стороны, в 2018 году Jingmai O’Connor и её коллеги поставили под сомнение идентификацию медуллярной кости, утверждая, что предполагаемая медуллярная кость была обнаружена только в костях передней конечности конфуциусорниса, в то время как у современных птиц она в основном откладывается в костях задних конечностей. Кроме того, рассматриваемая ткань сохранилась в виде маленьких фрагментов, что затрудняет её интерпретацию. Однако этим исследователям удалось выявить медуллярную кость в костях задней конечности энанциорнисовой птицы. Как и в случае с конфуциусорнисом, эта предполагаемая самка не достигла своих окончательных размеров, что подтверждает «динозавроподобный» способ роста мезозойских птиц, предполагавшийся в более ранних гистологических исследованиях[54].
Питание

В 1999 году китайский палеонтолог Lianhai Hou и его коллеги предположили, что конфуциусорнис являлся растительноядным, отмечая, что клюв конфуциусорниса был не похож на клюв современных хищных птиц, однако, содержимое желудка конфуциусорниса оставалось неизвестным на момент исследования, что указывает на то, что клюв птицы был изогнут вверх и был неприспособлен для умерщвления добычи[12]. В том же году палеонтологи Dieter S. Peters и Ji Qiang выдвинули гипотезу, что хотя не найдено никаких остатков перепонок между пальцами задних конечностей (указывающих на ихтиофагию), конфуциусорнис являлся ихтиофагом, и использовал свой довольно мягкий клюв для поиска добычи под поверхностью воды. Кроме того, авторы исследования подобрали несколько современных птиц в качестве наиболее близких аналогов конфуциусорниса. По мнению Dieter Peters, наиболее подходящим аналогом конфуциусорниса является белохвостый фаэтон (лат. Phaeton lepturus) — ихтиофаг, который, подобно конфуциусорнису, обладает длинным хвостом и узкими крыльями, а также часто гнездится рядом с вулканами[29].
Со своей стороны, в 2008 году польский палеонтолог Andrzej Elżanowski посчитал маловероятным, чтобы такая длиннокрылая и коротконогая птица как конфуциусорнис кормилась в кронах деревьев, вместо чего выдвинул свою версию о том, что конфуциусорнис охотился на лету, хватая добычу с поверхности земли / поверхности воды. Со слов Andrzej Elżanowski, на такую стратегию охоты указывают:
- длинные крылья образцов конфуциусорниса, по-видимому, адаптированные для парящего полёта;
- пропорции задних конечностей (длинные бедренные кости и короткие стопы), напоминающие таковые у современных фрегатов (лат. Fregata) и зимородковых (лат. Alcedinidae);
- большое затылочное отверстие (лат. foramen magnum), открывавшееся сзади;
- беззубый клюв, похожий на клюв современных кукабар (лат. Dacelo), только более короткий;
- отсутствие видимых адаптаций для плавания.
Кроме того, Andrzej Elżanowski допускал, что конфуциусорнис умел плавать[55].
В 2003 году китайские палеонтологи Zhonghe Zhou и Fucheng Zhang заявили, что хотя не найдено никаких прямых доказательств рациона конфуциусорниса, принимая во внимание его крепкие и беззубые челюсти, можно предположить, что он питался семенами. Кроме того, исследователи отметили, что внутри ископаемых остатков джехолорниса сохранились прямые доказательства подобного рациона (семена)[56].
В 2006 году Johan Dalsätt и его коллеги описали образец C. sanctus (IVPP V13313), найденный в пластах геологической формации Цзюфотан (Jiufotang), внутри ископаемых остатков которого сохранилось от семи до девяти позвонков и несколько рёбер, принадлежавших рыбе, вероятно, Jinanichthys. Данные кости рыбы были близко расположены, образуя скопление диаметром около 6 мм (0,24 дюйма), находившееся рядом с седьмым и восьмым шейными позвонками образца конфуциусорниса. Со слов авторов исследования, состояние вышеупомянутых костей рыбы указывает на то, что она либо хранилась в зобе конфуциусорниса, либо конфуциусорнис собирался отрыгнуть её в виде погадки (она поднялась по пищеводу незадолго до гибели птицы). В ходе раскопок не было обнаружено никаких других ископаемых остатков рыб в плите. Несмотря на то, что неизвестно, какую часть рациона конфуциусорниса составляла рыба, данное открытие не подтверждает фитофагию, которая изначально предполагалась для конфуциусорниса. Кроме того, исследователи отметили, что внутри ископаемых остатков конфуциусорниса ни разу не было найдено гастролитов — камней, обломков минералов и горных пород, которые целенаправленно заглатываются современными животными с относительно слабой (по отношению к употребляемой пище) зубной системой, и присутствующие в желудках многих (преимущественно зерноядных) современных птиц. Авторы исследования заключили, что конфуциусорнис, возможно, являлся всеядным, как, например, врановые[57].
Со своей стороны, в 2009 году Андрей Валерьевич Зиновьев предположил, что конфуциусорнис ловил рыбу с поверхности воды на лету[58].
Сочленение костей черепа конфуциусорниса являлось неподвижным, тогда как кинетизм черепа был выражен в гораздо меньшей степени, чем у современных птиц — верхняя челюсть животного являлась относительно неподвижной по отношению к черепной коробке. Данная неподвижность была обусловлена наличием Y-образной заглазничной кости (лат. os postorbitale), отделявшей глазницу от подвисочного отверстия (англ. infratemporal fenestra) (подобно более базальным тероподам), и наличием предчелюстной кости, полностью доходившей до лобной кости, прижимая носовые кости к боковым сторонам черепа птицы[8].
В 2020 году Case Vincent Miller и его коллеги предположили, что у конфуциусорниса было относительно слабое крепление клюва и рамфотеки, что даёт основания полагать, что клюв птицы мог выдерживать только низкие нагрузки. Тем не менее, функциональные последствия крепления клюва у современных птиц всё ещё недостаточно изучены, чтобы делать какие-либо категоричные выводы по этому поводу. В связи с слабым креплением клюва, конфуциусорнис, вероятно, был способен питаться лишь относительно мягкой пищей: у современных животных, питающихся мягкой пищей, челюсти, как правило, хуже приспособлены к нагрузкам, возникающим в процессе откусывания, измельчения и / или пережёвывания твёрдой пищи. Данный вывод подтверждает метод конечных элементов нижней челюсти C. sanctus, свидетельствующий о том, что нижняя челюсть птицы испытывала низкие нагрузки, сопоставимые с таковыми у современных насекомоядных и растительноядных птиц, но не сопоставимые с таковыми у современных хищных и зерноядных птиц. Авторы исследования заключили, что конфуциусорнис и прочие конфуциусорнитиды (лат. Confuciusornithidae) могли питаться твёрдой пищей (семенами) лишь в том случае, если бы они заглатывали гастролиты, однако, гастролиты и остатки проглоченных семян, известные у других мезозойских птиц, не найдены ни у одного известного конфуциусорнитида[59].
Примечания
Комментарии
- Проход, через который у живородящих и яйцеживородящих рождается детёныш, в то время как у яйцеродящих откладываются яйца.
- График, отображающий зависимость массы животного от его возраста.
- Изометрический рост — это равномерный рост частей тела, в процессе которого не меняются пропорции тела. Противопоставляется аллометрическому росту.
Источники
- Курочкин Е. Н. Базальная диверсификация пернатых. Эволюция биосферы и биоразнообразия. — К 70-летию А. Ю. Розанова. — М. : Т-во науч. изд. КМК, 2006. — С. 219—232.
- Ivanov M., Hrdlickova S. & Gregorova R. (2001) The Complete Encyclopedia of Fossils. Rebo Publishers, Netherlands. pp. 312.
- Xu Xing, Mark A. Norell. Non-avian dinosaur fossils from the Lower Cretaceous Jehol Group of western Liaoning, China (англ.) // Geological Journal. — 2006-09. — Vol. 41, iss. 3—4. — P. 419—437. — ISSN 1099-1034 0072-1050, 1099-1034. — doi:10.1002/gj.1044.
- Zhonghe, Zhou; Lianhai, Hou. “The discovery and study of Mesozoic birds in China” // Mesozoic birds: above the heads of dinosaurs (англ.) / Luis M. Chiappe; Lawrence M. Witmer. — Berkeley/Los Angeles/London: University of California Press, 2002. — P. 160—183. — ISBN 0-520-20094-2.
- Zhou, Z. The discovery of Early Cretaceous birds in China // Acta Palaeornithologica. — 1995. — Т. 181. — С. 9—22.
- Hou, L.; Zhou, Z.; Gu, Y.; Zhang, H. Confuciusornis sanctus, a new Late Jurassic sauriurine bird from China (англ.) // Chinese Science Bulletin : journal. — 1995. — Vol. 40, no. 18. — P. 1545—1551.
- Chiappe, Luis M. The first 85 million years of avian evolution (англ.) // Nature. — 1995. — Vol. 378, no. 6555. — P. 349—355. — doi:10.1038/378349a0. — .
- Chiappe, Luis M.; Shu-An, Ji; Qiang, Ji; Norell, Mark A. “Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China”. — Bulletin of the American Museum of Natural History, 1999. — Т. 242. — 89 с.
- Chiappe, Luis M.; Qinjin, Meng. Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs (англ.). — JHU Press, 2016. — ISBN 978-1421420240.
- Hou, L. (1997). Mesozoic Birds of China (PDF) (in Chinese). Translated by Downs, W. Phoenix Valley Provincial Aviary of Taiwan.
- Li, D.; Sulliven, C.; Zhou, Z.; Zhang, Z. Basal birds from China: a brief review (англ.) // Chinese Birds. — 2010. — Vol. 1, no. 2. — P. 83—96. — doi:10.5122/cbirds.2010.0002.
- Hou, L.; Martin, L. D.; Zhou, Z.; Feduccia, A.; Zhang, F. A diapsid skull in a new species of the primitive bird Confuciusornis (англ.) // Nature : journal. — 1999. — Vol. 399, no. 6737. — P. 679—682. — doi:10.1038/21411. — .
- Marugán-Lobón, Jesús; Chiappe, Luis M.; Ji, Shu'an; Zhou, Zhonghe; Chunling, Gao; Hu, Dongyu; Meng, Qinjing. Quantitative patterns of morphological variation in the appendicular skeleton of the Early Cretaceous bird Confuciusornis (англ.) // Journal of Systematic Palaeontology : journal. — Taylor & Francis, 2011. — Vol. 9, no. 1. — P. 91—101. — doi:10.1080/14772019.2010.517786.
- Zhang, Z.; Gao, C.; Meng, Q.; Liu, J.; Hou, L.; Zheng, G. Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China (англ.) // Journal of Ornithology : journal. — 2009. — Vol. 150, no. 4. — P. 783—790. — doi:10.1007/s10336-009-0399-x.
- Li, L. (2010). “[Chinese] [A new species of Confuciusornis from Lower Cretaceous of Jianchung, Liaoning, China]”. Global Geology. 29 (2): 183—187.
- Chiappe, L. M.; Marugán-Lobón, J.; Ji, S.; Zhou, Z. Life history of a basal bird: morphometrics of the Early Cretaceous Confuciusornis (англ.) // Biology Letters : journal. — Royal Society Publishing, 2008. — Vol. 4, no. 6. — P. 719—723. — doi:10.1098/rsbl.2008.0409. — PMID 18832054.
- Wang, M; O'Connor, J; Zhou, Z H. A taxonomical revision of the Confuciusornithiformes (Aves: Pygostylia) (англ.) // Vertebrata PalAsiatica : journal. — doi:10.19615/j.cnki.1000-3118.18053.
- Clarke, Julia A., Norell, Mark A. (2002) “The Morphology and Phylogenetic Position of Apsaravis ukhaana from the Late Cretaceous of Mongolia” // American Museum Novitates, No. 3387, American Museum of Natural History, New York, NY 10024.
- Mayr, G.; Pohl, B.; Peters, D. S. A well-preserved Archaeopteryx specimen with theropod features (англ.) // Science : Journal. — 2005. — Vol. 310, no. 5753. — P. 1483—6. — doi:10.1126/science.1120331. — . — PMID 16322455.
- Xing Xu, Hailu You, Kai Du and Fenglu Han. An Archaeopteryx-like theropod from China and the origin of Avialae (англ.) // Nature : journal. — 2011. — 28 July (vol. 475). — P. 465—470. — doi:10.1038/nature10288.
- Clarke, J. A.; Zhou, Z; Zhang, F. Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui (англ.) : Journal. — 2006. — Vol. 208, no. 3. — P. 287—308. — doi:10.1111/j.1469-7580.2006.00534.x. — PMID 16533313.
- O'Connor, Jingmai K.; Wang, Xuri; Chiappe, Luis M.; Gao, Chunling; Meng, Qingjin; Cheng, Xiaodong; Liu, Jinyuan. Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species (англ.) // Journal of Vertebrate Paleontology : journal. — 2009. — Vol. 29, no. 1. — P. 188—204. — doi:10.1080/02724634.2009.10010371. — .
- Thomas R. Holtz. Dinosaurs : the most complete, up-to-date encyclopedia for dinosaur lovers of all ages. — 1st ed. — New York: Random House, 2007. — 427 pages с. — ISBN 978-0-375-82419-7.
- R. L. Nudds, G. J. Dyke. Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability (англ.) // Science. — 2010-05-14. — Vol. 328, iss. 5980. — P. 887—889. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1188895.
- G. S. Paul. Comment on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability” (англ.) // Science. — 2010-10-15. — Vol. 330, iss. 6002. — P. 320—320. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1192963.
- Zihui Zhang, Chunling Gao, Qingjin Meng, Jinyuan Liu, Lianhai Hou. Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China (англ.) // Journal of Ornithology. — 2009-10. — Vol. 150, iss. 4. — P. 783—790. — ISSN 2193-7206 2193-7192, 2193-7206. — doi:10.1007/s10336-009-0399-x.
- L. D. Martin, Z. Zhou, L. Hou, A. Feduccia. Confuciusornis sanctus Compared to Archaeopteryx lithographica (англ.) // Naturwissenschaften. — 1998-06. — Vol. 85, iss. 6. — P. 286—289. — ISSN 1432-1904 0028-1042, 1432-1904. — doi:10.1007/s001140050501.
- Amanda R. Falk, Thomas G. Kaye, Zhonghe Zhou, David A. Burnham. Laser Fluorescence Illuminates the Soft Tissue and Life Habits of the Early Cretaceous Bird Confuciusornis (англ.) // PLOS One / Matthew Shawkey. — 2016-12-14. — Vol. 11, iss. 12. — P. e0167284. — ISSN 1932-6203. — doi:10.1371/journal.pone.0167284.
- Dieter Stefan Peters, Ji Qiang. Mußte Confuciusornis klettern? (нем.) // Journal of Ornithology. — 1999-01. — Bd. 140, H. 1. — S. 41—50. — ISSN 1439-0361 0021-8375, 1439-0361. — doi:10.1007/BF02462087.
- Fucheng Zhang, Stuart L. Kearns, Patrick J. Orr, Michael J. Benton, Zhonghe Zhou. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds (англ.) // Nature. — 2010-02. — Vol. 463, iss. 7284. — P. 1075—1078. — ISSN 1476-4687 0028-0836, 1476-4687. — doi:10.1038/nature08740.
- R. A. Wogelius, P. L. Manning, H. E. Barden, N. P. Edwards, S. M. Webb. Trace Metals as Biomarkers for Eumelanin Pigment in the Fossil Record (англ.) // Science. — 2011-09-16. — Vol. 333, iss. 6049. — P. 1622—1626. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1205748.
- Quanguo Li, Julia A. Clarke, Ke-Qin Gao, Jennifer A. Peteya, Matthew D. Shawkey. Elaborate plumage patterning in a Cretaceous bird (англ.) // PeerJ. — 2018-11-02. — Vol. 6. — P. e5831. — ISSN 2167-8359. — doi:10.7717/peerj.5831.
- L. Schmitz, R. Motani. Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology (англ.) // Science. — 2011-05-06. — Vol. 332, iss. 6030. — P. 705—708. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1200043.
- Zhou Z. and Farlow, J.O. (2001) “Flight capability and habits of Confuciusornis”. In: Gauthier and Gall (eds). New perspectives on the origin and early evolution of birds: proceedings of the international symposium in honor of John H. Ostrom. Peabody Museum of Natural History. Yale University, New Haven. pp. 237—254
- Hembree, D. (1999). “Re-evaluation of the posture and claws of Confuciusornis”. Journal of Vertebrate Paleontology. 19: 50A. DOI:10.1080/02724634.19.99.10011202.
- X. Zheng, X. Xu, Z. Zhou, D. Miao, F. Zhang. Comment on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability” (англ.) // Science. — 2010-10-15. — Vol. 330, iss. 6002. — P. 320—320. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1193223.
- R. L. Nudds, G. J. Dyke. Response to Comments on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability” (англ.) // Science. — 2010-10-15. — Vol. 330, iss. 6002. — P. 320—320. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1193474.
- Zhonghe Zhou, Paul M. Barrett, Jason Hilton. An exceptionally preserved Lower Cretaceous ecosystem (англ.) // Nature. — 2003-02. — Vol. 421, iss. 6925. — P. 807—814. — ISSN 1476-4687 0028-0836, 1476-4687. — doi:10.1038/nature01420.
- Robert J. Knell, Darren Naish, Joseph L. Tomkins, David W. E. Hone. Sexual selection in prehistoric animals: detection and implications (англ.) // Trends in Ecology & Evolution. — 2013-01. — Vol. 28, iss. 1. — P. 38—47. — doi:10.1016/j.tree.2012.07.015.
- Anusuya Chinsamy, Luis M. Chiappe, Jesús Marugán-Lobón, Gao Chunling, Zhang Fengjiao. Gender identification of the Mesozoic bird Confuciusornis sanctus (англ.) // Nature Communications. — 2013-06. — Vol. 4, iss. 1. — P. 1381. — ISSN 2041-1723. — doi:10.1038/ncomms2377.
- Winfried S. Peters, Dieter Stefan Peters. Life history, sexual dimorphism and ‘ornamental’ feathers in the mesozoic bird Confuciusornis sanctus (англ.) // Biology Letters. — 2009-12-23. — Vol. 5, iss. 6. — P. 817—820. — ISSN 1744-957X 1744-9561, 1744-957X. — doi:10.1098/rsbl.2009.0574.
- Kaiser, Gary W. The inner bird: anatomy and evolution : [англ.]. — UBC Press, 2010. — ISBN 9780774813440.
- Gareth J. Dyke, Gary W. Kaiser. Cracking a developmental constraint: egg size and bird evolution. In Proceedings of the VII International Meeting of the Society of Avian Paleontology and Evolution, ed. W.E. Boles and T.H. Worthy (англ.). Records of the Australian Museum (26 мая 2010).
- D. Charles Deeming, Gerald Mayr. Pelvis morphology suggests that early Mesozoic birds were too heavy to contact incubate their eggs (англ.) // Journal of Evolutionary Biology. — 2018-05. — Vol. 31, iss. 5. — P. 701—709. — ISSN 1420-9101 1010-061X, 1420-9101. — doi:10.1111/jeb.13256.
- David J. Varricchio, Frankie D. Jackson. Reproduction in Mesozoic birds and evolution of the modern avian reproductive mode (англ.) // The Auk. — 2016-10. — Vol. 133, iss. 4. — P. 654—684. — ISSN 1938-4254 0004-8038, 1938-4254. — doi:10.1642/AUK-15-216.1.
- Zhang, F.; Hou, L.; Ouyang, L. (1998). “Osteological microstructure of Confuciusornis: preliminary report”. Vertebrata PalAsiatica [англ.]. 36: 126—135.
- Zhang, F.-C.; Xu, X.; Lü, J.; Ouyang, L. (1999). “Some microstructure difference among Confuciusornis, Alligator and a small theropod dinosaur, and its implications”. Paleoworld [англ.]: 296—308.
- Anusuya Chinsamy, Luis M. Chiappe, Peter Dodson. Growth rings in Mesozoic birds (англ.) // Nature. — 1994-03. — Vol. 368, iss. 6468. — P. 196—197. — ISSN 1476-4687 0028-0836, 1476-4687. — doi:10.1038/368196a0.
- Kevin Padian, Armand J. de Ricqlès, John R. Horner. Dinosaurian growth rates and bird origins (англ.) // Nature. — 2001-07. — Vol. 412, iss. 6845. — P. 405—408. — ISSN 1476-4687 0028-0836, 1476-4687. — doi:10.1038/35086500.
- A. J. De Ricqlès, K. Padian, J. R. Horner, E.-T. Lamm, N. Myhrvold. Osteohistology of Confuciusornis sanctus (Theropoda: Aves) (англ.) // Journal of Vertebrate Paleontology. — 2003-06-17. — Vol. 23, iss. 2. — P. 373—386. — ISSN 1937-2809 0272-4634, 1937-2809. — doi:10.1671/0272-4634(2003)023[0373:OOCSTA]2.0.CO;2.
- L. M. Chiappe, J. Marugán-Lobón, A. Chinsamy. Palaeobiology of the Cretaceous bird Confuciusornis : a comment on Peters & Peters (2009) (англ.) // Biology Letters. — 2010-08-23. — Vol. 6, iss. 4. — P. 529—530. — ISSN 1744-957X 1744-9561, 1744-957X. — doi:10.1098/rsbl.2009.1057.
- Estelle Bourdon, Jacques Castanet, Armand de Ricqlès, Paul Scofield, Alan Tennyson. Bone growth marks reveal protracted growth in New Zealand kiwi (Aves, Apterygidae) (англ.) // Biology Letters. — 2009-10-23. — Vol. 5, iss. 5. — P. 639—642. — ISSN 1744-957X 1744-9561, 1744-957X. — doi:10.1098/rsbl.2009.0310.
- Winfried S. Peters, Dieter Stefan Peters. Sexual size dimorphism is the most consistent explanation for the body size spectrum of Confuciusornis sanctus (англ.) // Biology Letters. — 2010-08-23. — Vol. 6, iss. 4. — P. 531—532. — ISSN 1744-957X 1744-9561, 1744-957X. — doi:10.1098/rsbl.2010.0173.
- Jingmai O’Connor, Gregory M. Erickson, Mark Norell, Alida M. Bailleul, Han Hu. Medullary bone in an Early Cretaceous enantiornithine bird and discussion regarding its identification in fossils (англ.) // Nature Communications. — 2018-12. — Vol. 9, iss. 1. — P. 5169. — ISSN 2041-1723. — doi:10.1038/s41467-018-07621-z.
- Elżanowski, A. (2002) “Biology of basal birds and the origin of avian flight”. In: Zhou Z., Zhang F. (eds) Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1—4 June 2000. Science, Beijing, pp 211—226
- Zhonghe Zhou, Fucheng Zhang. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China (англ.) // Canadian Journal of Earth Sciences. — 2011-02-09. — doi:10.1139/e03-011.
- J. Dalsätt, Z. Zhou, F. Zhang, P. G. P. Ericson. Food remains in Confuciusornis sanctus suggest a fish diet (англ.) // Naturwissenschaften. — 2006-09. — Vol. 93, iss. 9. — P. 444—446. — ISSN 1432-1904 0028-1042, 1432-1904. — doi:10.1007/s00114-006-0125-y.
- A. V. Zinoviev. An attempt to reconstruct the lifestyle of confuciusornithids (Aves, Confuciusornithiformes) (англ.) // Paleontological Journal. — 2009-07. — Vol. 43, iss. 4. — P. 444—452. — ISSN 1555-6174 0031-0301, 1555-6174. — doi:10.1134/S0031030109040145.
- Case Vincent Miller, Michael Pittman, Thomas G. Kaye, Xiaoli Wang, Jen A. Bright. Disassociated rhamphotheca of fossil bird Confuciusornis informs early beak reconstruction, stress regime, and developmental patterns (англ.) // Communications Biology. — 2020-09-21. — Vol. 3, iss. 1. — P. 1—6. — ISSN 2399-3642. — doi:10.1038/s42003-020-01252-1.
Литература
- Dalsätt, J.; Zhou, Z.; Zhang, F. & Ericson, Per G. P. (2006). Food remains in Confuciusornis sanctus suggest a fish diet. // Naturwissenschaften 93(9): 444—446. doi:10.1007/s00114-006-0125-y (HTML abstract)
- Hou, L.; Zhou, Z.; Gu, Y. & Zhang, H. (1995). [Description of Confuciusornis sanctus]. // Chinese Science Bulletin 10: 61—63.
- Hou, L.-H.; Zhou, Z.; Martin, L.D. & Feduccia, A. (1995): A beaked bird from the Jurassic of China. // Nature 377: 616—618. doi:10.1038/377616a0 (HTML abstract)
- de Ricqlès, A.J.; Padian, K.; Horner, J.R.; Lamm, E.-T. & Myhrvold, N. (2003): Osteohistology of Confuciusornis sanctus (Theropoda: Aves). // Journal of Vertebrate Paleontology 23(2): 373—386. DOI:10.1671/0272-4634(2003)023[0373:OOCSTA]2.0.CO;2 HTML abstract
- Mayr, G.; Pohl, B. & Peters, D. S. (2005). A well-preserved Archaeopteryx specimen with theropod features. // Science 310(5753): 1483—1486. doi:10.1126/science.1120331 (HTML abstract) Supporting Online Material
- Senter, Phil (2006): Scapular orientation in theropods and basal birds, and the origin of flapping flight. // Acta Palaeontologica Polonica 51(2): 305—313. PDF fulltext
- Zhou, Z. & Zhang, F. (2003): Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. // Naturwissenschaften 90: 220—225. PDF fulltext
| Таксономия |
|---|