Tethyshadros
Tethyshadros (лат.) — род растительноядных орнитоподовых динозавров из надсемейства гадрозавроидов, чьи окаменелые остатки найдены в верхнемеловых отложениях Европы. Типовой и единственный вид Tethyshadros insularis назвал и описал итальянский палеонтолог Фабио Марко Далла Веккья в 2009 году. Родовое название образовано из названия древнего океана Тетис с добавлением слова лат. hadros, которое является отсылкой к группе гадрозавроидов. Видовое название insularis значит «островной». Голотип представляет первый обнаруженный полный и сочленённый скелет гадрозавроида[1].
| † Tethyshadros | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: † Цераподы Инфраотряд: Парвотряд: Надсемейство: Род: † Tethyshadros |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Tethyshadros Dalla Vecchia, 2009 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
Dalla Vecchia, 2009 |
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 83,6—66,6 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
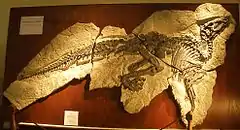

История исследования
Голотип SC 57021, представляющий собой целый сочленённый скелет, и относимый материал обнаружены в слоях формации Liburnia, датированных верхним кампаном — нижним маастрихтом, провинция Триест, Италия.
Дополнительные образцы: 57022 — неполные сочленённые передние конечности, SC 57023 — отдельная левая лобковая кость, SC 57025 — отдельный шейный позвонок с правым ребром, SC 57026 — неполный повреждённый череп и часть посткрания, SC 57247 — неполный сочленённый скелет, повреждённый во время раскопок, SC 57256 — отдельное спинное ребро[1].
Описание
Tethyshadros имеет следующие аутапоморфии: большой и удлинённый череп (длиннее плечевой кости в 1,6—1,65 раз; соотношение длины черепа к высоте около 2,6); предчелюстные зубы очень крупные, тонкие и заострённые; маленькие парные гребни в каудальной части теменных костей; скуловая кость очень длинная, тонкая, без вентрального выступа, часть от каудального до дорсального отростка в 2 раза длиннее рострального отростка; межвисочное отверстие почти прямоугольное, примерно в 2 раза больше глазницы; латеральный дистальный мыщелок квадратной кости расширенный и плоский вентрально (в форме шляпки гвоздя); длина тела первого хвостового позвонка больше его высоты; остистые отростки 1—6-го проксимальных хвостовых позвонков и, возможно также, дистальных спинных и крестцовых имеют форму топора; рёбра 1—5-го хвостовых позвонков имеют форму языка, краниокаудально расширенные и дорсовентрально уплощённые; длинный, хлыстовидный хвост с удлинёнными телами позвонков с 18-го по 32-й, в 2,4—3,4 раза длиннее высоты; тела хвостовых позвонков с 23 по 33 полуцилиндрической формы; длинный сегмент проксимальных хвостовых позвонков без гемапофизов (первый шеврон появляется между 7-м и 8-м хвостовыми позвонками), образующий промежуток в вентральной части, занимающий 44 % хвоста; дистальные концы гемапофизов 15—20-го позвонков имеют длинный задний отросток; лопаточная пластина асимметрично расширяется дистально (как у примитивных игуанодонтов камптозавра и дриозавра); завертлужный отросток подвздошной кости длинный, низкий, клиновидный (отсутствует низкий выступ) и сужается в латеральной проекции; очень длинная седалищная кость имеет сигмовидное тело и тупой, нерасширенный дистальный конец, не изогнутый и не сужающийся; только 3 пальца кисти (палец 5 отсутствует); плоские суставные концы пястных костей; 2 фаланги в 4-м пальце кисти, дистальная фаланга сильно редуцирована (2-я фаланга отсутствует у других гадрозавроидов); большеберцовая кость значительно длиннее бедренной. Tethyshadros имеет также следующие уникальные сочетания признаков: низкая подвздошная кость с большим и свисающим надвертлужным отростком, робустный предвертлужный отросток слабо изогнут, отсутствует дорсальная депрессия на надвертлужном отростке.
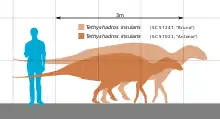
Длина скелета голотипа Tethyshadros — 3,62 м. Это маленький размер для гадрозавроида. Дополнительные экземпляры SC 57247 и SC 57022 относятся к крупным особям и более робустным, чем голотип, SC 57026 примерно того же размера, а SC 57023 принадлежит маленькой особи. Сращение тел позвонков с дугами, наличие едва заметных швов между большинством элементов черепа, степень оссификации, удлинённый череп и относительный размер глазниц не говорят в пользу неполовозрелости животного.
Длина черепа голотипа — 475 мм, высота — около 185 мм (соотношение — 2,57). Его череп длиннее черепов большинства гадрозавридов (за исключением Edmontosaurus annectens, у которого соотношение — 3,4). Среди базальных гадрозавроидов длина черепа Tethyshadros сопоставима с Dollodon bampingi (соотношение — 2,6) и только череп уранозавра превышает длину черепа Tethyshadros (соотношение — 3,2). По общему плану череп Tethyshadros напоминает базальных Dollodon bampingi и Mantellisaurus atherfieldensis. В заглазничной части в дорсальной проекции череп относительно широк в поперечнике, как у игуанодона, а также у него отсутствует характерное каудальное сужение, наблюдаемое у большинства гадрозавридов и уранозавра.
Позвоночный столб состоит из 11 шейных, 16 спинных, возможно, 8 крестцовых и более 33 хвостовых позвонков[1].
Филогения
По результатам филогенетического анализа с использованием 121 признака для 25 таксонов Tethyshadros находится за пределами семейства гадрозавриды и является родственным Telmatosaurus, однако с последним, а также со всеми продвинутыми гадрозавроидами на древе строго консенсуса, он образует неразрешимую политомию. В 55 одинаково экономных деревьях Tethyshadros либо сестринский таксон телматозавра (образуют кладу), либо продвинутее телматозавра, образует кладу со всеми продвинутыми гадрозавроидами, либо базальнее телматозавра, является сестринским таксоном к кладе телматозавр + все продвинутые гадрозавроиды. В дереве согласия по большинству Tethyshadros продвинутее телматозавра, и является сестринским таксоном к гадрозавридам.
Кладограмма на основе анализа[1]:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Особенности копытохождения гадрозавридов коритозавра и завролофа, когда часть веса животного, помимо копыта, несёт и предпоследняя фаланга (subunguligrady), обнаруживаются также и у Tethyshadros: трёхпалость, укорочение пальцев, копытовидные когти, фаланги значительно шире длины, отсутствие ямок коллатеральных связок, относительно уплощённые суставные поверхности.
У Tethyshadros имеются признаки, встречающиеся у бегающих амниот: большеберцовая кость длиннее бедренной, боковые пальцы утеряны, а кости метаподия длинные, тонкие и сцеплены вместе, действующие как единый элемент, уменьшение массы стопы (благодаря потере или укорочению фаланг и сужения костей метаподия) и копытохождение (subunguligrady). Tethyshadros имеет относительно низкое соотношение длин 3-й плюсневой кости к большеберцовой (0,35; схоже с коритозавром) и очень высокое — большеберцовой кости к бедренной (1,31; схоже с мелким двуногим гетеродонтозавром).
Особенности проксимальной части хвоста (длина отрезка хвоста без шевронов, удлинение тел позвонков, форма ребёр и шевронов) могут быть связаны с сильным развитием хвостово-бедренной мышцы — главного ретрактора бедра, что также может говорить о бегающем характере передвижения.
Запястье редуцировано до одной маленькой кости, 1-й и 5-й пальцы кисти и фаланга 4-го утрачены. Пястные кости тонкие, удлинённые, плотно прижатые друг к другу, с плоскими дистальными суставными поверхностями, как у большинства фаланг, поэтому невозможны были сгибания в плюсно-фаланговом и межфаланговых суставах. Предпоследние фаланги кисти 2-го и 3-го пальцев клиновидные, а 2-я и 3-я когтевые фалагни копытовидные. Проксимальные фаланги выступают, как продолжение пястных костей и вся передняя лапа выглядит, как столб. Способность к хватанию была, очевидно, сильно ограничена, и кисть вместе с маленькой терминальной частью действовала как единое целое. По-видимому, она могла использоваться только для квадрупедальной локомоции, хотя грацильность кисти, редуцированная терминальная часть и возможная ориентация может быть связана с ролью кисти как опоры во время отдыха и баланса во время бега на двух ногах. Возможно, это адаптация к своеобразному образу жизни (передвижение по пересечённой местности, характерной для карстовых ландшафтов)[1].
Палеобиогеография
Tethyshadros жил на Адриатико-Динарском острове (Adriatic-Dinaric Island), площадь поверхности которого равнялась примерно 100 000 км², что сравнимо с современной Кубой. Остров являлся частью Европейского архипелага. Все обнаруженные особи Tethyshadros, не являясь ювенильными, гораздо меньше взрослых гадрозавридов Северной Америки. Их маленький размер может объясняться островной карликовостью — феноменом, уже наблюдавшимся у других островных динозавров. Осевой и периферический скелет континентальных гадрозавридов и более базальных гадрозавроидов довольно консервативный, но у Tethyshadros он демонстрирует несколько апоморфий. Наличие необычных признаков также может бысть связано с островной изоляцией.
Примитивность телматозавра и других гадрозавроидов Европейского архипелага, в сравнении с одновозрастными в страгиграфическом плане и даже более древними гадрозавридами Северной Америки, может объясняться локальной эволюцией (начиная с середины мела) в условиях изоляции (модель викарианса). Тем не менее, Адриатико-Динарская карбонатная платформа (Adriatic-Dinaric Carbonate Platform, ADCP) была изолирована от других массивов суши с апта и была затоплена в сеномане — туроне во время трансгрессии. Перед аптом она могла соединяться с Африкано-Аравийским континентом, на котором до сих пор ещё не обнаруживались остатки послеаптских гадрозавроидов. До позднего мела на ADCP проживали в основном завроподы и тероподы. Однако из местонахождения Villaggio del Pescatore известны только гадрозавроиды, а в близком и примерно одновозрастном местонахождении Козина в Словении были извлечены зубы гадрозавроидов и тероподов. В позднесантонских динозавровых комплексах Венгрии и в раннекампанских — Франции и Австрии гадрозавроиды отсутствуют. Но они появляются в верхнем кампане — маастрихте Испании, Франции, Нидерландов, Бельгии, Германии, Словении, Италии, Румынии и Украины. Гадрозавроиды распространились в течение маастрихта, в то время как в позднем кампане они были достаточно редки. Доминирующими орнитоподами в течение сантона — раннего маастрихта на Европейском архипелаге были эндемичные рабдодонтиды.
Сестринские таксоны Tethyshadros и телматозавра азиатские Bactrosaurus, Gilmoreosaurus и Tanius из кампана — маастрихта и наиболее базальные гадрозаврины, известные на момент публикации, из маастрихта происходят из северного Китая. Североамериканские базальные гадрозавроиды датируются только барремом — туроном, в то время как хорошо известные гадрозавроиды кампана — маастрихта Северной Америки — это гадрозавриды. В отличие от Северной Америки, негадрозавридовые гадрозавроиды могли широко распространиться в Евразии в течение позднего мела. Всё это поддерживает гипотезу азиатского происхождения европейских таксонов. Гадрозавроиды могли колонизировать Европейский архипелаг из Азии (модель дисперсного видообразования), где негадрозавридовые гадрозавроиды всё ещё жили в кампане — раннем маастрихте. Коллизии микроплит вдоль северной окраины окена Тетис, имевшие место на первом этапе Альпийской складчатости, привели к образованию Трансильванского острова, где жил телматозавр, Адриатико-Динарского острова, где жил Tethyshadros и перешейков в восточных частях Европейского архипелага. В дополнение, тектонические поднятия земной коры и вулканизм вдоль геодинамически активной северной окраины Тетиса могли также создать миграционный путь из южной Азии. Гадрозавроиды, скорее всего, достигли восточной части Европейского архипелага «прыжками» с острова на остров вдоль юго-западной окраины Азии. Это расселение, вероятно, было первым в череде, имевшей место в позднем кампане[1].
Примечания
- Dalla Vecchia, F. M. (2009). «Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy». Journal of Vertebrate Paleontology. 29 (4): 1100—1116.

.jpg.webp)

