Arenysaurus
Аренизавр (лат. Arenysaurus) — род травоядных орнитоподовых динозавров семейства гадрозаврид из позднего мела Европы. Типовой и единственный вид Arenysaurus ardevoli назван и описан Хавьером Переда-Субербиолой и коллегами в 2009 году. Родовое имя ссылается на название деревни Арен, недалеко от которой были найдены останки динозавра. Видовое имя дано в честь геолога Ллуиса Ардеволя, открывшего местонахождение раскопок[1].
| † Arenysaurus | ||||||
|---|---|---|---|---|---|---|
 Реконструкция аренизавра | ||||||
| Научная классификация | ||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: † Цераподы Инфраотряд: Парвотряд: Надсемейство: Семейство: Подсемейство: Триба: Род: † Arenysaurus |
||||||
| Международное научное название | ||||||
|
Arenysaurus Pereda-Suberbiola et al., 2009 |
||||||
| Единственный вид | ||||||
|
||||||
| ||||||
История исследования
Голотип MPZ 2008/1, представляющий собой частично сочленённый череп, включающий крышу черепа и черепную коробку, многочисленные паратипы и дополнительные образцы обнаружены в слое формации Тремп, датированном поздним маастрихтом (около 67,6—65,5 млн лет назад), деревня Арен, провинция Уэска, Испания. Местонахождения, содержащие останки позвоночных, обнажились вдоль северного края синклинали Тремп в Центрально-Южных Пиренеях. Они расположены к западу от деревни Арен и недалеко от Blasi Hill. Местонахождения ископаемых останков пронумерованы Бласи 1 — Бласи 5. Бласи 1 располагается в верхней части формации Арен, остальные местонахождения находятся в нижней части формации Тремп, залегающей выше. Эти местонахождения происходят из отложений передних частей дельт, а также лагунных и прибрежных отложений осадочного комплекса Арен 4. Останки аренизавра были обнаружены в местонахождении Бласи 3, самом богатом на ископаемых позвоночных.
Большинство костей посткраниального скелета, причисленных аренизавру, были найдены в расчленённом состоянии, но в достаточно тесной связи с черепом. Они включают части осевого скелета: 7 шейных позвонков и 3 ребра и один спинной позвонок с двумя рёбрами, неполный крестец, состоящий из 5 сросшихся позвонков, и большую часть хвостового ряда, состоящую из 14 передних хвостовых позвонков; периферический скелет представлен костями грудного пояса (правая лопатка и коракоид), передней конечности (правая плечевая кость), тазовым поясом (правая подвздошная и лобковая кости) и задними конечностями (левая и правая бедренные кости).
Черепная коробка аренизавра полностью окостенела; отсутствие открытых черепных швов указывает, что череп голотипа не принадлежит молодой особи. Развитие лобной платформы, отсутствие предлобно-лобного «хомута», зубчатый шов между лобными и теменными костями и сращение верхнезатылосной кости с крышей черепа поддерживают эту трактовку. С другой стороны, череп, вероятно, не принадлежит старой особи, поскольку лобные кости не полностью срослись. Характер большинства швов вместе с размером черепа предполагает, что это взрослая особь. В дополнение к черепу, поперечные отростки и дуги позвонков срослись с их телами, а тела крестцовых позвонков срослись друг с другом, как это обычно бывает у зрелых особей[1].
В 2013 году Пенелопа Крузадо-Кабальеро с коллегами представила полное описание новых черепных элементов и посткраниального скелета, что повлияло на филогенетическое положение аренизавра[2]. В том же году Прието-Маркес с коллегами представил пересмотренную диагностику и совершенно другую филогенетическую позицию динозавра[3].
В 2015 году краниальный материал аренизавра с целью получения эндокрана был подвергнут компьютерной томографии в Лаборатории эволюции человека в университете города Бургос, Испания, с использованием промышленного томографа[4].
Описание
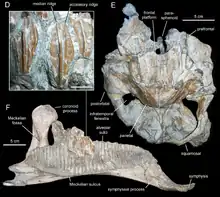
Длина аренизавра примерно 5—6 м.
Аренизавр диагностируется по следующим признакам: ярко выраженный купол лобной кости, более развитый, чем у других взрослых экземпляров; почти вертикальные предквадратный отросток чешуйчатой кости и скуловой отросток заглазничной кости; дельтопекторальный гребень плечевой кости ориентирован вперёд. Отличие от других ламбеозавринов проявляется в уникальном сочетании следующих признаков: короткая лобная кость с отношением длины задней части к ширине около 0,5; срединный бугор теменной кости находится приблизительно на уровне заглазнично-чешуйчатой дуги; теменная кость не вклинивается между чешуйчатыми костями и затылочной поверхностью черепа; латеральная сторона чешуйчатой кости расположена относительно низко над чашеобразной впадиной[1].
В пересмотренной Прието-Маркесом и коллегами диагностике всего одна аутапоморфия: сильно выступающий купол лобной кости, который поднимается по крайней мере на уровень дорсального края срединной ветви чешуйчатой кости.
Предполагаемая почти вертикальная ориентация скуловой ветви заглазничной кости и предквадратного отростка чешуйчатой кости возникает по причине субгоризонтального положения чешуйчатой ветви заглазничной кости. В действительности, однако, у сочленённых черепов ламбеозавринов каудальная часть крыши черепа, в большей или меньшей степени, обычно наклонена каудовентрально относительно длинной оси максиллярного зубного ряда, поэтому предквадратный и скуловой отростки выступают ростровентрально. Тем не менее, у многих, хотя не у всех образцов ламбеозавринов предквадратный отросток чешуйчатой кости и проксимальная часть скуловой ветви заглазничной кости почти перпендикулярны чешуйчатой ветви заглазничной кости над подвисочным отверстием. Эти сведения указывают, что ориентация предквадратного отростка и скуловой ветви заглазничной кости аренизавра не является аутапоморфией.
Переда-Субербиола также рассматривал краниальную ориентацию дельтопекторального гребня плечевой кости в качестве аутапоморфии. Однако, такая ориентация не уникальна для аренизавра; точно такая имеется у канардии, Parasaurolophus cyrtocristatus и зауролофина (по Прието-Маркесу) улагазавра. Более того, дельтопекторальный гребень сильно пострадал от эрозии. Этот эродированный останок соответствует наиболее проксимальному основанию гребня. Плохая сохранность и отсутствие большей его части препятствует точному установлению реальной ориентации дельтопекторального гребня.
Наконец, комбинация признаков, представленная Переда-Субербиолой и коллегами, для диагностики аренизавра не уникальна для этого таксона. Например, те же самые признаки мы видим у Hypacrosaurus stebingeri и H. altispinus. Более того, такое определение: латеральная сторона чешуйчатой кости, находящаяся относительно низко над чашеобразной впадиной, подходит большинству черепов ламбеозавринов[3].
Филогения
Анализ Переда-Субербиолы и коллег (2009)
Для определения систематического положения аренизавра была построена матрица с использованием 57 признаков для 22 таксонов. Для этого использовался набор данных из исследования Паскаля Годфруа и коллег 2008 года для сахалиянии и улагазавра. По мнению авторов, на тот момент это был самый всеобъемлющий анализ филогении гадрозавридов. В анализ помимо аренизавра был добавлен также телматозавр и исключена сахалияния, поскольку её включение приводит к неопределённости в полученных результатах. По результатам филогенетического анализа аренизавр занимает довольно базальную позицию, являясь сестринским таксоном амурозавру и монофилетической группе, образованной трибами Corythosaurini (синоним Lambeosaurini) и Parasaurolophini. Аренизавр не продвинутый ламбеозаврин, поскольку его дентарная батарея (37 зубов) не соответствует состоянию, наблюдаемому у Parasaurolophini и Corythosaurini (42—45 зубов). Наличие очень короткой лобной кости с отношением длины задней части к максимальной ширине меньше 0,6 рассматривается, как признак приобретённый конвергентно у аренизавра и продвинутых ламбеозавринов.
Клада Parasaurolophini, образованная паразавролофом и харонозавром, определяется по четырём синапоморфиям, в том числе: платформа лобной кости тянется над передней частью надвисочного отверстия и дорсальный промонторий на заглазничной кости. У аренизавра теменная кость не входит в затылочную норму черепа, как и у паразавролофа и харонозавра. Это состояние рассматривается, как конвергентно развитое у аренизавра, поскольку у него отсутствуют две другие синапоморфии Parasaurolophini. Клада Corythosaurini, состоящая из ламбеозавра, коритозавра, гипакрозавра и олоротитана, характеризуется латеральным предчелюстным отростком, расширяющимся назад до слёзной кости[1].
Кладограмма, построенная на основе анализа Переда-Субербиолы и коллег (2009):
| Hadrosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Анализ Крузадо-Кабальеро и коллег (2013)
В 2013 году Крузадо-Кабальеро с коллегами провели новый филогенетический анализ на основе матрицы, предложенной Годфруа и коллегами в 2012 году, подкорректировав матрицу и включив новый материал, описанный в своей работе. Полученная матрица включала 119 признаков для 22 таксонов. По итогам анализа аренизавр и его ближайший испанский родственник бласизавр (из того же местонахождения) занимают более продвинутое положение в противоположность предыдущим работам Переда-Субербиолы и коллег, 2009, и Годфруа и коллег, 2012. Выяснилось, что клада аренизавр — бласизавр располагается внутри трибы Parasaurolophini. Несмотря на то, что три синапоморфии Parasaurolophini не могут быть кодированы для этих испанских ламбеозавринов, у аренизавра имеются следующие общие с паразавролофом признаки: каудальный отросток заглазничной кости проходит над подвисочным отверстием; короткая диастема между первым дентарным зубом и предзубной костью (также у бласизавра); широкие медиолатерально дистальные мыщелки плечевой кости; дельтовидный бугор лопатки дорсовентрально широкий и краниокаудально вытянутый с хорошо заметным вентральным краем. Тем не менее, оба таксона отличаются от других представителей Parasaurolophini широким язычным выступом симфизной части зубной кости[2].
Кладограмма, построенная на основе анализа Крузадо-Кабальеро и коллег (2013):
| Hadrosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Анализ Прието-Маркеса и коллег (2013)
В 2013 году Прието-Маркес с коллегами провёл филогенетический анализ для выяснения систематического положения четырёх европейских ламбеозавринов: аренизавра, бласизавра, парарабдодона и канардии в соответствии с их пересмотренными диагнозами. В анализе использовалось 265 признаков (179 краниальных и 86 посткраниальных) для 34 таксонов (9 внешних таксонов, гадрозавр, 2 представителя завролофинов и 22 ламбеозаврина). В результате было получено 5 самых экономных деревьев. Аренизавр и бласизавр оказываются глубоко вложенными внутри ламбеозавринов. Они образуют сестринскую кладу, поддержанную тремя однозначными синапоморфиями зубной кости: относительно короткая диастема с отношением между её длиной и расстоянием между самой ростральной позицией зуба и каудальным краем венечного отростка меньше 0,2 (конвергентно с велафронсом); угол наклона вентрального края зубной кости до 20 градусов; каудальный конец зубной батареи прилегает вплотную к каудальному краю венечного отростка.
Включение этих двух испанских родов в кладу ламбеозавринов, состояющую из последнего общего предка гипакрозавра, амурозавра и всех его потомков, поддерживается одной неоднозначной синапоморфией: отсутствие или очень плохое развитие вентрального поперечного каудального гребня между базиптеригоидными отростками базисфеноида (неизвестно для олоротитана). На более инклюзивном уровне клада аренизавр — бласизавр однозначно помещается внутри трибы Lambeosaurini на основе следующих признаков: сочленовая поверхность лобной кости с носовой имеет форму наклонённой ростровентрально платформы; раздвоение ростромедиального края лобных костей в сагиттальной плоскости крыши черепа; средняя длина отростка перед чашеобразной впадиной чешуйчатой кости от 0,95 до 1,25 ширины чашеобразной впадины. Эти синапоморфии не могут наблюдаться в имеющемся материале бласизавра, однако, поскольку он находится в сестринских отношениях с аренизавром, эти признаки были оптимизированы для клады испанских динозавров в целом.
Полученная филогения в корне отличается от предыдущих исследований Переда-Субербиолы и Крузадо-Кабальеро. Такой результат объясняется вследствие использования исследователями матриц признаков, содержащих, по существу, различную информацию. А именно, пармисонический анализ Переда-Субербиолы использовал только 57 признаков (на 208 меньше, чем в данном анализе); в матрице отсутствовали посткраниальные признаки. Матрица Крузадо-Кабальеро содержит едва ли половину признаков матрицы Прието-Маркеса.
Далее Прието-Маркес ссылается на не опубликованную работу Крузадо-Кабальеро от 2012 года, где аренизавр и бласизавр также формируют кладу внутри трибы Parasaurolophini на основе признаков: равносторонний краниально поднимающийся отросток таранной кости и расширенный дистальный конец малоберцовой кости. Однако, малоберцовая и таранная кости не сохранились у рассматриваемых испанских динозавров. Признаки, поддерживающие близкие отношения аренизавра и бласизавра с паразавролофом, включают: широкие дистальные мыщелки плечевой кости; относительно короткое сужение предлобковой кости, которая расширяется у основания проксимального конца предлобкового отростка; заглазничная кость с короткой чешуйчатой ветвью, показывающей наузкий дорсальный края подвисочного отверстия. Несмотря на сильно эродированную дистальную часть плечевой кости аренизавра, имеется много внутритаксонных вариантов ширины дистальных мыщелков плечевой кости гадрозавридов, поэтому Прието-Маркес не считает данный признак филогенетически информативным. Относительно короткое проксимальное сужение предлобкового отростка безусловно имеется у Parasaurolophus walkeri и Parasaurolophus cyrtocristatus (у Parasaurolophus tubicen лобковая кость не найдена), но также и у Hypacrosaurus altispinus, Hypacrosaurus stebingeri и Magnapaulia laticaudus. Вентральное расширение предлобкового отростка встречается у всех ламбеозавринов. Более того, вентральная область предлобкового отростка у аренизавра слишком плохо сохранилась для того, чтобы выяснить величину и геометрию вентрального расширения этого отростка. Однако, Прието-Маркес согласен, что аренизавр демонстрирует глубокое рострокаудальное сужение чешуйчатой ветви заглазничной кости с сопутствующим укорочением дорсального края подвисочного отверстия, который приобретает латерально дугообразную форму. Это состояние безусловно наблюдается у Parasaurolophus walkeri и Parasaurolophus tubicen, но отсутствует у Parasaurolophus cyrtocristatus. Более того, последний демонстрирует относительно длинный чешуйчатый отросток и прямой, почти прямоугольный дорсальный край подвисочного отверстия, что более похоже на коритозавра, ламбеозавра или гипакрозавра. Стоит отметить, что базальный ламбеозаврин яксартозавр также обладает коротким чешуйчатым отростком заглазничной кости. В анализе Прието-Маркеса и коллег это состояние заглазничной кости представлено, как однозначная синапоморфия Parasaurolophus walkeri и Parasaurolophus tubicen, а у аренизавра и яксартозавра выступает, как независимо приобретённый признак. Таким образом, с учётом имеющихся анатомических данных двух испанских ламбеозавринов, исследование Прието-Маркеса не поддерживает присутствие представителей Parasaurolophini на Иберо-Армориканском острове Европейского архипелага[3].
Кладограмма, построенная на основе анализа Прието-Маркеса и коллег (2013):
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
С помощью компьютерной томографии был получен почти полный трёхмерный эндокран аренизавра. Структуры на левой стороне эндокрана хорошо сохранились и поддались цифровой визуализации, в то время, как правая сторона плохо сохранилась и в некоторых случаях не может быть восстановлена. В целом, возможно наблюдать неполные обонятельные луковицы, полушарие головного мозга, мозжечок, начало продолговатого мозга, гипофизарную ямку, внутреннее ухо и каналы почти для каждого нерва со II по XII.
Эндокран аренизавра типичный для гадрозавридов и удлинён в передне-заднем направлении. Он имеет длину 116,5 мм от основания обонятельного тракта до каудальной ветви подъязычного нерва. Максимальная ширина поперёк полушария мозга — 48,4 мм, а предполагаемый объём эндокрана (включая обонятельные луковицы) — 126,2 см³. Общий объём полушария — 65,4 см³, составляя 53,3 % от общего объёма эндокрана (за исключением обонятельных луковиц). Этот результат схож с результатом, полученным для образца взрослого амурозавра. С другой стороны эндокран аренизавра значительно сужен латеромедиально на уровне мозжечка с максимальной шириной в этой части — 31,3 мм и слегка сужен в продолговатом мозге (26,3 мм).
Угол главной оси полушария мозга по отношению к горизонтали близок к 45 градусам. Такая высота угла соответствует подсемейству ламбеозавринов, в противоположность гадрозавринам и остальным орнитоподам, у которых полушарие мозга располагается почти горизонтально. Угол изгиба между мозжечком и полушарием очень маленький, почти 10 градусов. В этом отношении эндокран близок к взрослым лавразийским ламбеозавринам (Hypacrosaurus altispinus и амурозавр).
В первоначальной работе Переда-Субербиолы указывалось, что образец аренизавра по нескольким остеологическим характеристикам принадлежит взрослой особи. Палеонейроанатомические данные, включающие очень маленький угол изгиба между мозжечком и полушарием мозга и плохо различимые на срезе КТ черепные швы поддерживают этот вывод. Однако, несколько признаков говорят о подростковой онтогенетической стадии развития этого образца: общий объём эндокрана (без обонятельных луковиц); объём полушария мозга (без обонятельных луковиц); максимальная ширина полушария. Это запутывающее сочетание признаков взрослой и подростковой стадий развития может говорить, возможно, о первом случае карликовости.
Более того, Farke с коллегами в 2013 году предположили, что у гадрозавридов, а также маленького орнитопода Dysalotosaurus lettowvorbecki имеется дуральный пик (угол дорсального края мозжечка, не его выступ), который не меняется в ходе онтогенетического развития. Далее, эти авторы предполагают, что филогенетические различия между трибами Lambeosaurini и Parasaurolophini могли бы быть оценены по углу дурального пика. Lambeosaurini имеют широкий угол (около 120 градусов), а Parasaurolophini — более острый (примерно 90 градусов). У всех наблюдаемых в данном исследовании гадрозавринов и ламбеозавринов угол был не меньше 100 градусов. В случае аренизавра этот угол равен примерно 114 градусам. Как итог, угол дурального пика может быть действительно информативным: большой угол может являться базальным признаком, а угол меньше 100 градусов может быть особенностью рода паразавролофов. Касательно внутреннего уха, хотя общий вид похож на других гадрозавридов, можно наблюдать небольшие различия в полукружных каналах. Морфология внутреннего уха аренизавра сближает его с трибой Parasaurolophini. В целом, различия в вестибулярной системе (соотношение между передним и задним полукружным каналом и размер ампулы) могут быть использованы в качестве филогенетического сигнала для дифференциации эдмонтозавра, паразавролофа и аренизавра от других гадрозавридов. Уменьшение разницы между длиной переднего и заднего полукружных каналов и возможно также высоты этих каналов может отражать снижение компенсаторных движений глаз и головы у эдмонтозавра, паразавролофа и аренизавра. Это может быть связано с поведенческими моделями, которые требуют меньшую подвижность головы[4].
Палеобиогеография
Самые базальные и ранние ламбеозаврины встречаются в Азии, которая может рассматриваться в качестве их предковой области: аралозавр из турона — сантона Казахстана, циньтаозавр из кампана восточного Китая, яксартозавр из сантона Казахстана и амурозавр из маастрихта восточной России. Другие азиатские ламбеозаврины являются продвинутыми формами из маастрихта восточной России и северо-восточного Китая (харонозавр, олоротитан, сахалияния). Ниппонозавр известен из сантона — раннего кампана острова Сахалин, Россия, но его филогенетическое положение и статус остаются спорными. На западе Северной Америки все известные ламбеозаврины являются продвинутыми формами, состоящими в кладе, образованной Corythosaurini и Parasaurolophini (коритозавр, гипакрозавр, ламбеозавр, велафронс и паразавролоф). Старейшие таксоны датированы кампаном.
Открытие ламбеозаврина из маастрихта Пиренейского полуострова и палеобиогеографическая история ламбеозавринов может говорить о позднем расселении из Азии в Европу в течение второй половины позднего мела. Палеогеографические реконструкции, тем не менее, указывают, что появление Тургайского пролива, эпиконтинентального моря, располагавшегося с востока Уральских гор, образовало препятствие для расселения между Европой и Азией после сеномана. Тесные отношения между европейскими и азиатскими ламбеозавринами показывают, что такой барьер, кажется, не препятствовал фаунистическому обмену ламбеозавринов между Азией и Европой в течение позднего мела[1].
Глобальный подъём уровня моря начался в апте и достиг высшей точки в позднем сеномане — раннем туроне, создав архипелаг между Африкано-Аравийской плитой и надводной частью Фенносарматского щита (Североевропейский континент). В позднем мелу встречные движения Африкано-Аравийской и Евразийской плит, возникшее вследствие формирования Южной Атлантики, и последующие столкновения микроплит, находящихся между ними, привело к локальному тектоническому подъёму и образованию массивов суши. Европейский архипелаг состоял из Английского, Иберо-Армориканского, Рейнско-Богемского, Адриатического, Австро-Альпийского и Трансильванского (остров Тисия-Дакия или Хатег) и других островов. У разных авторов разные реконструкции количества и площадей этих островов.
Европейская гадрозавриновая фауна позднего маастрихта характеризовалась ламбеозавринами (аренизавр, бласизавр, парарабдодон и канардия из Иберо-Армориканского острова) и негадрозавридовыми гадрозавроидами. Тем не менее, на сегодняшний момент в Европе нет домаастрихтских останков однозначно относимых к ламбеозавринам или гадрозавридам. Таксономический состав фауны европейских ламбеозавринов более похож на одновозрастную фауну Азии, чем Северной Америки, поскольку в маастрихте североамериканские ламбеозаврины представлены исключительно Hypacrosaurus altispinus, который может быть датирован нижним—нижним верхним маастрихтом. В противоположность, ламбеозаврины достигли многообразия и относительно широко распространились в верхнем маастрихте восточной Азии.
Для реконструкции предковых ареалов для клады, состоящей из Hypacrosaurus altispinus и ламбеозавринов из Бласи аренизавра и бласизавра, сначала был проведён Statistical Dispersal-Vicariance анализ. Неустойчивое положение олоротитана в пяти филогенетических деревьях внутри Lambeosaurini приводит к двум различным реконструкциям предковых ареалов для этой клады. В частности, когда олоротитан выступает в качестве сестринского таксона ламбеозаврам из Бласи, их последний общий предок реконструирован, как живший в Евразии; во всех остальных случаях предок заселял бы Северную Америку и Европейский архипелаг. Тем не менее, во всех случаях присутствие ламбеозавринов из Бласи на Иберо-Армориканском острове будет представлять собой викарианс, который имел место в маастрихте.
Вышеприведённые сценарии контрастируют с результатами анализов Bayesian Binary MCMC и Dispersal Extinction Cladogenesis. Ещё более неопределённой выглядит история клады ламбеозавринов, включающей два вида из Бласи и их общего сестринского таксона, усложнённая, опять же, нестабильным филогенетическим положением олоротитана. С одной стороны, результат BBM анализа указывает на Северную Америку, как наиболее вероятный предковый ареал этой клады во всех пяти самых экономных деревьев. Следуя этой реконструкции, аренизавр и бласизавр проникли на Иберо-Армориканский остров посредство расселения из Северной Америки в течение маастрихта. С другой стороны, результаты DEC анализа существенно варьируются в зависимости от положения олоротитана, изображая, либо Евразию, Северную Америку + Европейский архипелаг, либо исключительно Северную Америку в качестве наиболее вероятного предкового ареала для этой клады ламбеозавринов. Эти выводы предусматривают несколько биогеографических сценариев для таксонов из Бласи, появившихся в течение маастрихта, от викарианса, приводящего к разделению Азиатского или Северо Американского ареала и Европейского, до переселения из Северной Америки на Европейский архипелаг.
Викариантный сценарий, предложенный S-DIVA, находится в противоречии с текущими ископаемыми европейских ламбеозавринов. Как указывалось выше, не имеется окаменелостей ламбеозавринов, однозначно датированных временем до верхнего маастрихта, несмотря на останки гадрозавроидов, датированных верхним кампаном — маастрихтом из отложений Бельгии, Нидерландов, Германии, Словении, Италии, Болгарии, Румынии и Украины. Ламбеозаврины также отсутствуют в верхнем кампане — нижнем маастрихте среди местонахождений наземных позвоночных на Пиренейском полуострове. Они также не зафиксированы в верхнем кампане — нижнем маастрихте в южной Франции. Единственный зуб из местонахождения Lano — одно лишь предполагаемое свидетельство о гадрозавридах в Европе в позднем кампане; местонахождения позвоночных в Европе, датированные сантоном — верхним кампаном, лишены останков гадрозавридов, хотя в этих местонахождениях присутствуют рабдодонтиды. Поэтому предковые арелы, полученные посредством анализов BBM и DEC и соответствующие сценариям расселения, больше согласуются с известными ископаемыми ламбеозавринов в Европе[3].
Примечания
- Pereda-Suberbiola, Xabier; Jose Ignacio Canudo; Penelope Cruzado-Caballero; Jose Luis Barco; Nieves Lopez-Martinez; Oriol Oms; Jose Ignacio Ruiz-Omenaca (2009). «The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain)». Comptes Rendus Palevol 8 (6): 559—572.
- Cruzado-Caballero, P. L.; Canudo, J. I.; Moreno-Azanza, M.; Ruiz-Omenaca, J. I. (2013). «New material and phylogenetic position of Arenysaurus ardevoli, a lambeosaurine dinosaur from the late Maastrichtian of Aren (northern Spain)». Journal of Vertebrate Paleontology 33 (6): 1367—1384.
- Prieto-Marquez, A.; Dalla Vecchia, F. M.; Gaete, R.; Galobart, A. (2013). Dodson, Peter, ed. «Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis». PLoS ONE 8 (7): e69835.
- Cruzado-Caballero P, Fortuny J, Llacer S, Canudo J. (2015) Paleoneuroanatomy of the European lambeosaurine dinosaur Arenysaurus ardevoli. PeerJ 3:e802

.jpg.webp)

