Halobacteriaceae
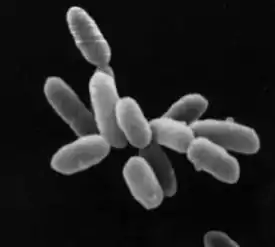
Halobacteriaceae (лат.) — семейство архей из класса галобактерий[1] (Halobacteria), единственное в порядке Halobacteriales. Включает около двадцати родов, в том числе Halobacterium, Halococcus, Haloarcula, Natrococcus, Natrobacterium.
| Halobacteriaceae | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Тип: Класс: Порядок: Halobacteriales Grant and Larsen 1989 Семейство: Halobacteriaceae |
||||||||||
| Международное научное название | ||||||||||
| Halobacteriaceae Gibbons 1974 | ||||||||||
| ||||||||||
Представители семейства живут в средах с высоким содержанием солей[2], в том числе в Мёртвом море, где концентрация соли достигает 26—27 %, а в некоторые годы повышается до 31 %, а также в Большом Солёном Озере штата Юта (США), где концентрация соли достигает 32 %[3] (при 36 % NaCl выпадает в осадок), на кристаллах соли в прибрежной полосе, в солончаках, на солёной рыбе, на засолённых шкурах животных, на рассольных сырах, в капустных и огуречных рассолах.
Строение, метаболизм
Halobacteriaceae — кокковидные или палочковидные, подвижные или неподвижные аспорогенные микроорганизмы. Большинство из них окрашиваются грамположительно. У некоторых имеются газовые вакуоли для контроля плавучести[4].
Преимущественно аэробы, но могут переносить и очень низкое содержание кислорода в среде, свободноживущие сапрофиты. По типу источника энергии это фототрофы, по донору электронов — органотрофы и по источнику углерода — гетеротрофы. При наличии кислорода и органических соединений, которые можно использовать в качестве источника энергии, галобактерии способны развиваться и в темноте (то есть в зависимости от условий фотоорганогетеротрофы либо хемоорганогетеротрофы). Однако при недостатке или даже при полном отсутствии кислорода и ярком освещении в оболочке клеток синтезируется бактериородопсин, позволяющий использовать энергию Солнца[5]. Из-за большого содержания каротиноидов галобактерии окрашены в красные, оранжевые и жёлтые тона. Наиболее распространённый каротиноид — бактериоруберин.
Мембранный бислой построен из изопрениловых диэфиров фосфоглицерина, с небольшим содержанием неполярных липидов — C30-изопреноидов и с очень большим содержанием белков. В мембране различают участки пурпурного и красно-оранжевого цветов. В пурпурной мембране 75 % массы приходится на один белок — бактериородопсин[5]. Бактериородопсин функционирует как светочувствительный протонный насос. После абсорбции фотона конформация белка изменяется таким образом, что новая конформация позволяет транспорт одного протона из клетки в среду. Создаваемый таким образом градиент концентрации протонов используется для синтеза АТФ[6]. В мембране также содержится два так называемых сенсорных родопсина, которые обеспечивают положительный и отрицательный фототаксис[7][8]. Молекулы сенсорных родопсинов абсорбируют фотоны различных длин волн, что запускает каскад сигналов, управляющих жгутиковым двигателем. Кроме того, в мембране имеется галородопсин, представляющий собой светозависимый насос ионов хлора[9]. Его основная функция — транспорт в клетку Cl− против градиента электрохимического потенциала. Этот транспорт необходим для поддержания осмотического давления внутри клетки, растущей в среде с высоким содержанием соли[10]. Жизнь в гипоксическом рассоле привела к выработке у галобактерий мощной системы активного транспорта, благодаря которой концентрация Na+ в цитоплазме поддерживается на низком уровне, несмотря на колоссальный концентрационный градиент Na+ на клеточной мембране.

Следует отметить, что чем выше концентрация натрия в среде, тем выше содержание K+ внутри клеток. Известно, что калий необходим для работы большинства внутриклеточных ферментов, тогда как натрий подавляет активность многих из них. При существенном снижении концентрации соли в среде, клетки бактерий разрушаются. Посредником синтеза АТФ служит концентрационный градиент H+ на мембране галобактерий[5]. Он создаётся и поддерживается системой активного транспорта, переносящей протоны из среды в цитоплазму. За счёт этого pH в цитоплазме стабилен, очень низок — около 3 и мало зависит от щелочности водоёма, где pH может достигать 12.
Экология
Судя по их строению, галобактерии — одни из древнейших обитателей нашей планеты. Человечеству они известны довольно давно по красноватому налёту на продуктах, консервируемых с использованием больших количеств поваренной соли. Впервые галобактерии были выделены в начале прошлого столетия из микрофлоры лиманной грязи, однако их систематическое изучение началось только в конце второго десятилетия двадцатого века. У них практически нет врагов или конкурентов, способных жить в таких же условиях, и поэтому галобактерии свободно эволюционировали на протяжении всей истории жизни на Земле. Галобактерии не наносят никакого существенного вреда народному хозяйству. Очевидно, что внутренняя среда человека непригодна для жизнедеятельности галобактерий, поэтому среди них нет ни одного патогена.
Примечания
- Галобактерии // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов. — М. : Большая российская энциклопедия, 2004—2017.
- Tortora, Funke, Case, 2007, p. 316, 340.
- Reece J. B., Urry L. A., Cain M. L., Wasserman S. A., Minorsky P. V., Jackson R. B., Rawle F., Durnford D., Moyes C., Walde S., Wilson K. 2014. Campbell Biology, Canadian edition, Pearson Canada Inc., pp. 595, 606. ISBN 978-0-321-77830-7.
- Tortora, Funke, Case, 2007, p. 96.
- Tortora, Funke, Case, 2007, p. 147.
- Alberts B., Johnson A., Lewis J., Morgan D., Raff M., Roberts K., Walter P. 2015. Molecular biology of the cell, 6th edition. Garland Science: New York, pp. 586—587. ISBN 978-0-8153-4464-3.
- Grote M., Engelhard M., Hegemann P. 2014. Of ion pumps, sensors and channels - perspectives on microbial rhodopsins between science and history. Biochimica et Biophysica Acta 1837(5): 533—545. doi:10.1016/j.bbabio.2013.08.006.
- Spudich J. L., Sineshchekov O. A., Govorunova E. G. 2014. Mechanism divergence in microbial rhodopsins. Biochimica et Biophysica Acta 1837(5): 546—452. doi:10.1016/j.bbabio.2013.06.006.
- Song Y., Gunner M. R. 2014. Halorhodopsin pumps Cl- and bacteriorhodopsin pumps protons by a common mechanism that uses conserved electrostatic interactions. Proceedings of the National Academy of Sciences of the USA 111(46): 16377—16382. doi:10.1073/pnas.1411119111
- Pfisterer C., Gruia A., Fischer S. 2009. The mechanism of photo-energy storage in the Halorhodopsin chloride pump. Journal of Biological Chemistry 284(20): 13562—13569. doi:10.1074/jbc.M808787200.
Литература
- Tortora G. J., Funke B. R., Case C. L. Microbiology: An introduction : [англ.]. — 9th edition. — San Francisco : Pearson Education, 2007. — ISBN 0-8053-4791-7.
| Брожение | |
|---|---|
| Фотосинтез | |
| Хемосинтез | |
| Анаэробное дыхание |
|
| Категории |  | ||||||
|---|---|---|---|---|---|---|---|
| Примечательные экстремофилы |
| ||||||
| Связанные статьи |
| ||||||