Тарбозавр
Тарбоза́вр[1] (лат. Tarbosaurus, от др.-греч. τάρβος — «ужас, предмет боязни» и σαῦρος — «ящерица») — род гигантских (по одной из оценок, до 14 метров длиной) хищных ящеротазовых динозавров семейства тираннозаврид, живших во времена позднемеловой эпохи (83,6—66,0 млн лет назад[2]) на территории нынешних Монголии и Китая.
| † Тарбозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Клада: Клада: Надсемейство: Семейство: Род: † Тарбозавр |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Tarbosaurus Maleev, 1955 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
† Tarbosaurus bataar (Maleev, 1955) |
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 83,6—66,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||


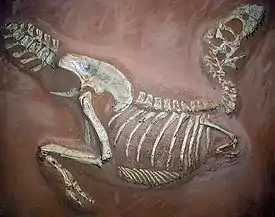
Обнаруживаемые с 1946 года ископаемые остатки нескольких десятков особей тарбозавра, включая целые черепа и скелеты, позволяют воссоздать его внешний облик и сделать выводы об образе жизни, а также проследить его эволюцию. С 1955 года, когда название Tarbosaurus было впервые использовано Е. А. Малеевым, исследователи дали обнаруженным ископаемым остаткам несколько видовых названий. Однако к началу XXI века в научном сообществе сомнений не вызывает существование только одного из этих видов, Tarbosaurus bataar (реже, из-за значительного сходства с североамериканскими тираннозаврами, их классифицируют как Tyrannosaurus bataar). Близкими родичами тарбозавра также являются обнаруженные в Монголии алиорамы.
Тарбозавры были крупными двуногими хищниками с массой тела, сопоставимой с массой тираннозавров, и непропорционально маленькими относительно остального тела двупалыми передними конечностями. В пасти тарбозавра располагались около шести десятков зубов по меньшей мере до 85 мм длиной. Хотя ряд исследователей считает тарбозавров падальщиками, более распространена точка зрения, согласно которой они были активными хищниками и занимали верхнее положение в экосистеме своей эпохи и региона, охотясь на крупных травоядных динозавров во влажных речных поймах.
Классификация и систематика
| Кладограммы тираннозаврид | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Тарбозавра как правило относят к семейству тираннозаврид подотряда теропод, к подсемейству тираннозаврин[3]. Последнее утверждение разделяют только часть систематиков, например, Люй с коллегами в работе 2014 года включили род непосредственно в семейство тираннозаврид[5].
По своему внешнему виду тарбозавр ближе к тираннозавру, чем к альбертозавру и горгозавру. Его отличает более массивное телосложение, пропорционально больший череп и пропорционально более длинные подвздошные кости, чем у представителей второй эволюционной ветви семейства, включающей альбертозавра и горгозавра; в то же время у этой второй ветви более длинные относительно общих размеров тела большеберцовые кости и кости плюсны[6]. Часть исследователей рассматривают T. bataar как вид тираннозавров; эта точка зрения высказывалась как вскоре после открытия[7], так и в более новых исследованиях[3][8]. В частности, Кен Карпентер оспаривает даже устоявшуюся точку зрения, согласно которой тарбозавры были мельче, чем тираннозавры, а все отличия в строении черепа между тарбозавром и тираннозавром, по его мнению, укладываются в рамки вариаций в строении черепа разных особей внутри вида Tyrannosaurus rex[8]. Другие авторы выделяют тарбозавров в отдельный вид, хотя по прежнему признают их близкое родство с тираннозаврами[9].
Из исследования 2003 года следовало, что наиболее близки к тарбозаврам алиорамы, разделяющие с ними особенности черепа, отсутствующие у других видов тираннозаврин (см. Строение черепа). Если эта точка зрения найдёт подтверждение, это будет свидетельствовать о том, что эволюция тираннозаврин в Америке и Азии шла разными путями и что тарбозавра нельзя отождествлять с тираннозаврами[4][10]. При этом единственный известный представитель алиорамов — по ряду характеристик молодая особь, — по всей видимости, не является молодым тарбозавром, так как обладает бо́льшим количеством зубов (от 76 до 78) и специфической цепочкой костяных бугорков вдоль верхней части морды[11]. В то же время в исследовании 2013 года наиболее близким родственником тарбозавра назван ещё один азиатский тираннозаврид — Zhuchengtyrannus, а тираннозавр представлен как следующий по близости морфологических характеристик[12].
Место тарбозавра в кладистике относительно тираннозавра в XXI веке имеет точки пересечения с теориями о процессе распространения тираннозаврид в Северной Америке и Азии. В частности, в статье 2013 года, рассматривающей древний материк Ларамидия как место возникновения тираннозаврид, предполагается, что они затем распространились на территории Азии одной волной — в период падения уровня поверхности мирового океана в конце кампанского яруса. Дальнейшая эволюция тираннозаврид проходила параллельно в Америке и Азии, и таким образом американский тираннозавр эволюционно отстоит достаточно далеко от более близких друг к другу азиатских таксонов Tarbosaurus и Zhuchengtyrannus[12]. Напротив, исследование 2016 года, в числе авторов которого сторонники теории о большей близости тираннозавра и тарбозавра, предполагает, что именно Tyrannosaurus rex является потомком сформировавшихся в Азии гигантских тираннозаврид, проникших в Америку ещё позже — ближе к концу мелового периода, и именно поэтому это единственный гигантский вид, до сих пор открытый в Северной Америке, где помимо него находят только тираннозаврид средних размеров[13].
История открытия и наименования

В 1946 году Монгольская палеонтологическая экспедиция АН СССР под руководством И. А. Ефремова в Гоби, в Южно-Гобийском аймаке, обнаружила череп и несколько позвонков в Нэмэгэтской свите. В 1955 году советский палеонтолог Е. А. Малеев определил эти находки как голотип ранее не описанного вида, которому он дал название Tyrannosaurus bataar[14], представляющее собой искажённое монг. баатар (рус. герой)[15]. В этом же году Малеевым были описаны ещё три черепа тероподов, найденных в процессе той же экспедиции в 1948 и 1949 годах. С каждым из этих черепов были найдены другие детали скелета, и каждый был классифицирован Малеевым как принадлежащий отдельному виду. Первый комплект получил название Tarbosaurus efremovi — новое родовое название, образованное от др.-греч. τάρβος (рус. ужас, почтение) и σαῦρος (рус. ящерица), и видовое имя, данное в честь советского палеонтолога и писателя-фантаста И. А. Ефремова. Остальные два черепа были классифицированы как новые виды известного по Северной Америке рода горгозавр (соответственно, G. lancinator и G. novojilovi). Все эти экземпляры были менее крупными, чем первый[14].
В 1965 году А. К. Рождественский опубликовал статью, в которой все четыре экземпляра, описанные ранее Малеевым, были определены как представители одного и того же вида на разных стадиях роста. По мнению Рождественского, этот вид не был тождественным североамериканскому Tyrannosaurus rex. В этот вид, для которого Рождественский предложил имя Tarbosaurus bataar, он включал не только описанные в 1955 году образцы, но и новые материалы[16]. В более поздних публикациях, включая работы самого Малеева[17], выводы Рождественского были признаны верными, хотя некоторые авторы предпочитали использовать видовое название Tarbosaurus efremovi, а не T. bataar[18]. Однако в 1988 году Грегори С. Пол в своём обзоре хищных динозавров вновь относит Tarbosaurus efremovi к роду Tyrannosaurus[19]. Четырьмя годами позже американский палеонтолог Кеннет Карпентер вновь проанализировал исследованные Рождественским образцы и пришёл к выводу, что ископаемые остатки принадлежат представителям рода Tyrannosaurus, согласившись с первоначальными выводами Малеева. Карпентер отнёс к виду Tyrannosaurus bataar все экземпляры, кроме одного, который был описан Малеевым как Gorgosaurus novojilovi. По мнению Карпентера, этот экземпляр представлял отдельный, более мелкий вид тираннозаврид, который он предложил назвать Maleevosaurus novojilovi[8]. Ещё одна точка зрения была высказана в 1995 году популяризатором палеонтологии Джорджем Ольшевским, который предложил новое родовое имя Jenghizkhan (в честь Чингисхана) для Tarbosaurus bataar, а тарбозавра Ефремова и малеевозавра Новожилова рассматривал как ещё два отдельных рода, современных первому и обитавших в той же местности[20]. Канадский палеонтолог Томас Карр в 1999 году снова описывает малеевозавра как подростковую особь тарбозавра[21]. После 1999 года все публикации описывают только один вид либо под названием Tarbosaurus bataar[4][22][23], либо, реже, Tyrannosaurus bataar[3].
В 1963 году началась совместная польско-монгольская экспедиция в Гоби. В ходе экспедиции, продолжавшейся до 1971 года, было обнаружено много новых ископаемых остатков, включая несколько экземпляров тарбозавров в Нэмэгэтской свите[24]. Японско-монгольские экспедиции с 1993 по 1998 год[25], а также частные экспедиции ещё одного канадского палеонтолога, Филипа Карри, в начале XXI века также принесли новый костный материал, классифицированный как принадлежащий тарбозаврам[26][27]. В общей сложности обнаружены кости, принадлежащие более чем 30 экземплярам, включая свыше 15 черепов и несколько относительно полных скелетов[22].
Возможные синонимы
В 1976 году советский палеонтолог С. М. Курзанов по несколько более древним ископаемым остаткам, также обнаруженным в Монголии, описал ещё один новый род тираннозаврид, Alioramus[28]. Более поздний анализ показал близкое родство алиорама с тарбозавром[4][10]. Хотя найденный экземпляр был описан как взрослый, вытянутый и низкий череп характерен для подростковых особей, что заставило Карри предположить, что алиорам является всего лишь неполовозрелым тарбозавром; однако он же отметил большее количество зубов и наличие костяных гребней на морде алиорама, не позволяющие однозначно сделать подобный вывод[11].
В середине 1960-х годов китайские палеонтологи обнаружили в Синьцзян-Уйгурском автономном районе (формация Субаши) ископаемые остатки (неполный череп и скелет) небольшого теропода, получившего кодовый номер IVPP V4878. В 1977 году китайский палеонтолог Дун Чжимин описал этот экземпляр как представителя нового рода Shanshanosaurus huoyanshanensis[29]. В 1988 году в работе американского исследователя Грегори Пола шаньшанозавр был описан как представитель вида Aublysodon (вид тираннозаврид, позже исключённый из биологической типологии)[19]; впоследствии Дун и Карри, вновь проанализировав ископаемые остатки шаньшанозавра, пришли к выводу, что они принадлежат подростковой особи крупного тираннозаврида, но не сумели определить, какой именно вид они представляют, хотя тарбозавр был указан как один из возможных вариантов[30]; к таким же выводам пришла в 2011 году японско-монгольская исследовательская группа после изучения ископаемых остатков детёныша тарбозавра в возрасте 2—3 лет, имеющих значительное сходство с аналогичными ископаемыми остатками шаньшанозавра[31]. В последующие годы разрозненные ископаемые остатки тираннозаврид были найдены также в других районах КНР, и многим из них местные исследователи дали собственные названия; вместе с тем, ископаемые остатки, описанные как Albertosaurus periculosis, Tyrannosaurus luanchuanensis, Tyrannosaurus turpanensis и Chingkankousaurus fragilis, возможно, принадлежат тарбозаврам[22].
В 2009 году в Гоби были обнаружены ископаемые остатки хищного динозавра, несомненно относящегося к тираннозавридам. Вначале ископаемые остатки были описаны как подростковая особь тарбозавра, но позже определены уже как принадлежащие новому, намного меньшему по размерам, динозавру, рапторексу, и датированы более ранним геологическим периодом — нижним мелом[32][33]. В 2011 году группа американских палеонтологов сообщила о результатах нового анализа, согласно которым датировка костей неоднозначна и ископаемые остатки и всё же принадлежат детёнышу более крупного тираннозаврида — возможно, тарбозавра[34].
Внешний вид и строение
Известные ископаемые остатки тарбозавров, как и некоторых других крупнейших тираннозаврид, многочисленны, и многие из них хорошо сохранились. Только в Нэмэгэтской свите четверть всех обнаруженных костей принадлежит тарбозаврам[35]. Скелеты, черепа и отдельные кости тарбозавров представлены в собраниях музеев всего мира, в том числе в коллекциях Палеонтологического института РАН[36] и Института Палеобиологии Польской академии наук в Варшаве[37], в Музее естественной истории[38] и Палеонтологическом музее Академии наук Монголии[39] в Улан-Баторе (в июне 2013 года на центральной площади Улан-Батора был открыт временный музей, где экспонируется целый скелет тарбозавра[40]), Музее естественной истории Университета Токай (префектура Сидзуока, Япония), Национальном музее естественной истории в Париже и Мельбурнском музее (подростковая особь)[41]. Хотя тарбозавры изучены меньше, чем тираннозавриды Северной Америки[42], доступного материала достаточно, чтобы учёные пришли к определённым выводам об их организме.


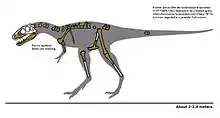
Уступая в размерах тираннозавру, тарбозавр всё равно оставался одним из самых крупных представителей семейства[43][44][45]. Длина черепа типового образца (PIN 551-1) составляет 122[7] или 135 см[19][44][46] — больше, чем у любого другого тираннозаврида, кроме собственно тираннозавра[46][45]. При этом голова у тарбозавра была пропорционально крупнее по отношению к телу, чем у других тираннозаврид, включая тираннозавра[44][47]. Известны почти полные скелеты молодых тарбозавров: PIN 552-1 (череп 114 см) и ZPAL MgD-1/3 (череп 74,5 см), длиной 7,7 и 5,5 метров, а их масса оценивается в 1650—2100 и 760 кг соответственно[47]. Другие хорошо известные образцы включают в себя ZPALMgD−I/4 — самый крупный образец (длина черепа более 110 см) из института палеобиологии в Варшаве, который был найден в 1964—1965 годах польско-монгольской экспедицией; а также MPC-D 107/2 (GIN 107/2) — скелет взрослой особи с сочленённым черепом длиной 122 см, который был найден японско-монгольской экспедицией и описан в 1984 году[48][49]. В 1988 году палеонтолог Грегори С. Пол указал PIN 551-1 в качестве крупнейшего известного образца тарбозавра, оценив его общую длину в 10 метров, а массу — в 5 тонн[19], но в 2016 году скорректировал эту оценку до 9,5 метров и 4 тонн[44]. С другой стороны, Зофья Келан-Яворовская сообщает о полных скелетах тарбозавров длиной до 13 метров, что сопоставимо с крупнейшими полными скелетами тираннозавров[50], а в издании 1983 года даже приводится максимальная возможная длина тарбозавра в 14 метров[51]. Масса тарбозавра считается такой же, или несколько меньшей, чем у тираннозавра сопоставимой длины[19][43][44][45].
Тираннозавриды мало отличались друг от друга внешне, и тарбозавр не был исключением из этого правила. Голова тарбозавра сидела на S-образной шее, а остальная часть позвоночника, включая длинный хвост, располагалась горизонтально. Крошечные передние конечности тарбозавра были ещё меньше по сравнению с остальным телом, чем у других представителей семейства. На каждой передней конечности, как и у других близкородственных видов, было по два когтистых пальца; у некоторых экземпляров имеется также третий палец, лишённый когтя. Т. Хольц указывает, что пальцы у тарбозавра были редуцированы сильней, чем у других тираннозаврид[52], поскольку вторая пястная кость у тарбозавра менее чем вдвое короче первой, тогда как у остальных видов это соотношение примерно равно 2:1. Третья пястная кость у тарбозавра также пропорционально меньше, чем у прочих тираннозаврид, будучи короче, чем первая, в то время как, например, у альбертозавра и дасплетозавра она длиннее, чем первая[53].
В отличие от передних, трёхпалые задние конечности тарбозавра были длинными и мощными, неся на себе всю массу тела. Длинный тяжёлый хвост служил противовесом голове и торсу, так что центр тяжести располагался в районе бёдер[14].
Строение черепа
.jpg.webp)
.jpg.webp)
Череп высокий, как и у тираннозавра, но не такой широкий, особенно в затылочной части. То, что череп не расширяется к затылку, означает, что глаза тарбозавра не глядели прямо вперёд и что, возможно, в отличие от тираннозавра, он не обладал бинокулярным зрением. Масса черепа была снижена за счёт больших окон (отверстий) в черепных костях. В челюстях размещалось 56—64 зуба, несколько больше, чем у тираннозавра, но не так много, как у тираннозаврид меньших размеров, таких как горгозавр или алиорам[4][54]. Большинство зубов были овальными в сечении, за исключением полукруглых в сечении зубов, располагавшихся на предчелюстной кости. Подобный гетеродонтизм характерен для всего семейства. Самые длинные зубы, до 85 миллиметров длиной, располагались в верхней челюсти.
Первое полное описание черепа тарбозавра было сделано в 2003 году. Было обращено внимание на ключевые отличия в строении черепа тарбозавра и североамериканских тираннозаврид, и многие из этих отличий связаны с тем, как распределяется давление по черепу в момент укуса. В момент укуса давление передаётся через верхнюю челюсть на соседние кости черепа. При этом у североамериканских тираннозаврид основная часть нагрузки передавалась через верхнюю челюсть в спаянные носовые кости в верхней части морды, прочно соединённые со слёзными костями костяными перемычками; кости были соединены достаточно прочно, чтобы можно было сделать вывод, что давление передавалось через носовые кости дальше, на слёзные. В то же время у тарбозавра эти костяные распорки отсутствовали, и связь между носовыми и слёзными костями была непрочной. С другой стороны, у тарбозавра, в отличие от североамериканских тираннозаврид, была массивная, хорошо развитая задняя часть верхней челюсти, входившая в «карман», образованный слёзной костью. Таким образом, видимо, давление при укусе передавалось у тарбозавра от верхней челюсти на слёзные кости напрямую. В свою очередь слёзные кости тарбозавра были прочнее, чем у его североамериканских родичей, соединены с лобной и переднелобной костью, в целом делая общую конструкцию верхней челюсти более жёсткой[55].
Ещё одним принципиальным различием между тарбозавром и североамериканскими тираннозавридами была жёсткая конструкция нижней челюсти. В то время как у многих теропод, включая тираннозаврид Северной Америки, соединение задних и передних костей челюсти обладало известной гибкостью, нижнюю челюсть тарбозавра характеризует наличие запирающего механизма, образованного костной перемычкой между угловой костью и задней кромкой передней зубной кости. Существует гипотеза, согласно которой большая прочность нижней челюсти тарбозавра связана с тем, что добычей ему служили крупные зауроподы титанозавры, кости которых также найдены в Нэмэгэтской свите. Аналогичные по размерам травоядные ящеры практически не встречались в Северной Америке на протяжении позднего мелового периода[10].
Специфика строения черепа также играет роль в определении места тарбозавра в биологической типологии. Похожие особенности можно найти также у черепа алиорама, также обнаруженного в Монголии, что указывает на то, что именно алиорам, а не тираннозавр, является ближайшим родичем тарбозавра. Возможно, что схожие черты в облике тарбозавра и тираннозавра развились независимо, как прямое следствие их гигантских размеров, и представляют собой пример конвергентной эволюции[10].
Строение мозга
Один из найденных в 1948 году советско-монгольской экспедицией черепов тарбозавра (первоначально получивший название Gorgosaurus lancinator) характеризует полость, в которой находился мозг. Гипсовый слепок внутреннего рельефа полости (эндокрана) позволил Малееву сделать ряд первичных выводов о форме мозга тарбозавра[56]. В 2005 году на основе более совершенного слепка на основе полиуретановой резины был проделан новый детальный анализ строения и функционирования мозга тарбозавра[57].
Строение мозга тираннозавра также было объектом анализа несколько ранее[58], и результаты анализа мозга тарбозавра оказались похожими, за исключением расположения мест отхождения некоторых черепных нервов, включая тройничный и добавочный нерв. Мозг тираннозаврид ближе по строению к мозгу крокодилов и других пресмыкающихся, чем к мозгу птиц. Общий объём мозга 12-метрового тарбозавра, по оценке исследователей, составлял только 184 см³. Большая обонятельная луковица и мощные обонятельный и концевой нервы свидетельствуют о хорошо развитом чувстве обоняния у тарбозавра — как и у тираннозавра. Добавочная обонятельная луковица также отличается большими размерами, что указывает на наличие хорошо развитого якобсонова органа, отвечавшего за чувствительность к феромонам. Это, в свою очередь, может указывать на развитую систему ухаживания у тарбозавров. Большой слуховой нерв означает, что слух у тарбозавров был хорошим, позволяя вести звуковую коммуникацию и обеспечивать хорошую ориентацию в пространстве. Вестибулярный аппарат был хорошо развит; иными словами, тарбозавры обладали хорошим чувством равновесия и координацией движений. С другой стороны, нервы и мозговые отделы, связанные со зрением, были маленькими и недоразвитыми. Крыша среднего мозга, отвечающая у пресмыкающихся за обработку зрительной информации, у тарбозавра была совсем маленькой, а зрительный и глазодвигательный нервы — слабыми. Если у тираннозавра, с его широким черепом, глаза смотрели вперёд, обеспечивая бинокулярное зрение, то череп тарбозавра, как и у большинства других тираннозаврид, был у́же, и глаза располагались по разные стороны головы. Всё это позволяет сделать вывод о том, что основными чувствами, развитыми у тарбозавра, были обоняние и слух, но не зрение[57].
Палеоэкология
Местом обнаружения подавляющего большинства известных ископаемых остатков тарбозавра является Нэмэгэтская свита в пустыне Гоби, южная Монголия. Возраст этой геологической формации никогда не определялся радиоизотопным методом, но фауна Нэмэгэтской свиты характерна для маастрихтского яруса верхнего мела[35]. Этот ярус покрывает эпоху между 70 и 66 миллионами лет назад[59]. Формация Субаши, в которой обнаружены ископаемые остатки шаншанозавра, также соответствует по возрасту маастрихтскому ярусу[60].
Исследование изотопов кислорода зубной эмали тарбозавра показало, что в формации Нэмэгэт наблюдалась сезонность со средними годовыми температурами на 10 °C выше, чем в современной Монголии. Сезонная структура δ18O продемонстрировала сходство с современной глобальной сетью изотопов Шицзячжуан в осадочной станции, Северный Китай, показав, что в формации Нэмэгэт был прохладный умеренный муссонный климат. Значения изотопов углерода растительноядных и плотоядных (δ13C) из зубной эмали указывают на наличие в прошлом на территории формации Нэмэгэт лесной экосистемы, в которой преобладали хвойные деревья, такие как араукарии, что согласуется с гипотезой о том, что крупные завроподы и гадрозавриды были наиболее предпочтительной добычей тарбозавра. Среднее годовое количество осадков, основанное на взаимосвязи между современными голосеменными и местным среднегодовым количеством осадков, оценивается в 775—835 мм/год[61].
В нэмэгэтских отложениях сохранились русла больших рек и почвенные наносы, свидетельствующие о том, что климат в это время был намного более влажным, чем в более ранних Барун-Гойотской и Джадохтской формациях, однако встречающиеся горизонты каличе указывают на то, что периодические засухи всё же имели место. В формации попадаются окаменелые моллюски и ископаемые остатки других водных организмов, таких как рыбы и черепахи[35]. Найдены несколько видов крокодиломорф, в первую очередь относящихся к роду Shamosuchus, зубы которых приспособлены к разгрызанию раковин[62]. Ископаемые остатки млекопитающих в Нэмэгэтской свите крайне редки, но было обнаружено много птиц, включая гурилиний из подкласса энанциорнисовых и принадлежащих к гесперорнисообразным юдинорнисам, а также древних гусеобразных тевиорнисов.

Исследователями описаны многочисленные динозавры из Нэмэгэтской свиты, в том числе такие анкилозавриды, как Tarchia, и такие пахицефалозавриды, как Homalocephale и Prenocephale[35]. Взрослые тарбозавры, будучи крупнейшими хищными динозаврами в данной формации, вероятнее всего охотились на крупных гадрозавров — зауролофов и барсболдий, или на таких титанозавров, как немегтозавр[10]. Тарбозаврам не могли составить конкуренцию более мелкие тероподы, такие как троодонтиды (бороговии, заурорнитоидесы и прочие), овирапторозавры (Elmisaurus, Nemegtomaia, Rinchenia) или Bagaraatan (которого Хольц называет базальным родом в семействе тираннозаврид, но который по размеру был в несколько раз меньше). Среди других гигантских тероподов некоторые (в частности, теризинозавр) могли быть травоядными, а орнитомимозавры — ансеримим, галлимим, дейнохейрус — всеядными и охотиться за более мелкой дичью, не конкурируя за пищу с тарбозавром. Молодые особи тарбозавров, как это, вероятно, происходило с другими крупными тираннозавридами и происходит в настоящее время с крокодилами и комодскими варанами, возможно, занимали промежуточные экологические ниши между взрослыми тарбозаврами и более мелкими тероподами[63].
Наряду с точкой зрения, согласно которой тарбозавры, как и другие гигантские тираннозавриды, были доминантными хищниками своей эпохи, новейшие исследования также рассматривают гипотезу, отводящую им другое место в экологическом балансе. С середины 1990-х годов появились оценки, основанные на массе и строении тела, по которым тираннозавр мог развивать скорость не более 30 километров в час, поскольку, если животное подобного веса и роста споткнётся, голова ударится о землю с такой силой, что череп расколется. Поэтому крупные тираннозавры не рисковали быстро бегать, так как это могло окончиться для них смертельным падением[64][65]. Скорее всего, питались тираннозавры и тарбозавры падалью. Для такого образа жизни не требовалось умения быстро бегать, и уменьшался риск разбиться при падении. Дополнительные аргументы в пользу того, что крупные тираннозавриды не могли быть эффективными охотниками, включают слабое зрение, короткие относительно длины бедра большеберцовые кости, редуцированные передние конечности и толстые зубы, непохожие на плоские саблеобразные зубы, типичные для хищных динозавров. Все эти доводы в разной степени опровергались сторонниками более распространённой точки зрения, согласно которой тираннозавриды были хищниками[66]. В то же время в 2010 году учёные из КНР и Японии также пришли к выводу о падальщицком образе жизни тарбозавра, исследовав кости скелета зауролофа, найденного в 1995 году в западной части Гоби. Особенности расположения и размеры отметин позволили учёным предположить, что их оставил тарбозавр, ископаемые остатки которого были найдены рядом со скелетом зауролофа[67].
Тарбозавр в культуре
В конце XX века, с появлением новых компьютерных технологий, стало возможным создание натуралистичных трёхмерных моделей давно вымерших или никогда не существовавших животных. Технология была с успехом применена в киноиндустрии, и динозавры были «оживлены» одними из первых. Трёхмерные изображения тарбозавров можно видеть в следующих фильмах и компьютерных играх:
- В 2003 году Би-би-си был выпущен документальный фильм «В стране гигантов»[68]. Тарбозавр появляется во второй части фильма — «Гигантский коготь», рассказывающей о меловых динозаврах Монголии.
- В 2006 году в Германии немецкой компанией SEK была разработана компьютерная игра ParaWorld. Тарбозавр появляется в дополнении boosterpack.
- В 2009 вышел двухсерийный документальный фильм «Баллада о тарбозавре» («Tarbosaurus the Mightiest Ever»). Режиссёр Хан Санг Хо (Han Sangho), EBS, Корея.
- В 2012 году тарбозавры стали главными героями художественного фильма «Тарбозавр 3D» («Jeombaki: Hanbandoeui Gongryong 3D»), созданного тем же южнокорейским режиссёром[69].
Примечания
- Тарбозавр // Струнино — Тихорецк. — М. : Советская энциклопедия, 1976. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 25).
- Tarbosaurus (англ.). Paleobiology Database Classic. (Дата обращения: 15 ноября 2016).
- Carr, Thomas D., Williamson, Thomas E., & Schwimmer, David R. A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama // Journal of Vertebrate Paleontology. — 2005. — Vol. 25, № 1. — P. 119—143. — doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- Currie, Philip J., Hurum, Jørn H., & Sabath, Karol. Skull structure and evolution in tyrannosaurid phylogeny // Acta Palaeontologica Polonica. — 2003. — Vol. 48, № 2. — P. 227—234.
- Lü J., Yi L., Brusatte S. L., Yang L., Li H. and Chen L. A new clade of Asian Late Cretaceous long-snouted tyannosaurids // Nature Communications. — 2014. — P. 1—10. — doi:10.1038/ncomms4788.
- Holtz, 2004, p. 130.
- Малеев Е. А. Гигантские хищные динозавры Монголии // Доклады Академии наук СССР. — 1955. — Т. 104, № 4. — С. 634—637.
- Ken Carpenter. Tyrannosaurids (Dinosauria) of Asia and North America // Aspects of Nonmarine Cretaceous Geology / Mateer, Niall J.; and Chen Peiji (eds.). — Beijing: China Ocean Press, 1992. — P. 250—268.
- Holtz, 2004, pp. 112—113.
- Hurum & Sabath, 2003, p. 188.
- Currie, Philip J. Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta // Acta Palaeontologica Polonica. — 2003. — Vol. 48, № 2. — P. 191—226.
- Lowen, M. A., Irmis, R. B., Sertich, J. J. W., Currie, P. J., & Sampson, S. D. Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans // PLoS One. — 2013. — Vol. 8, № 11. — P. e79420. — doi:10.1371/journal.pone.0079420.
- Brusatte, S. L., & Carr, T. D. The phylogeny and evolutionary history of tyrannosauroid dinosaurs // Scientific Reports. — 2016. — Vol. 6. — P. 20252. — doi:10.1038/srep20252.
- Малеев, 1955.
- Hurum & Sabath, 2003, p. 162.
- А. К. Рождественский. Возрастная изменчивость и некоторые вопросы систематики динозавров Азии // Палеонтологический журнал. — 1965. — № 3. — С. 95—109.
- Малеев Е. А. Гигантские карнозавры семейства Tyrannosauridae // Фауна и биостратиграфия мезозоя и кайнозоя Монголии / Труды ССМПЭ. — 1974. — № 3. — С. 132—191.
- Барсболд Р. Хищные динозавры мела Монголии // Труды Совместной советско-монгольской палеонтологической экспедиции. — 1983. — № 19. — С. 96—97. Архивировано 14 июля 2014 года.
- Gregory S. Paul. Predatory Dinosaurs of the World. — N. Y.: Simon & Schuster, 1988. — P. 464.
- George Olshevsky & Tracy L. Ford. The origin and evolution of the tyrannosaurids, part 1 (яп.) // Dinosaur Frontline. — 1995. — Vol. 9. — P. 92—119.
- Thomas D. Carr. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria) (англ.) // Journal of Vertebrate Paleontology. — 1999. — Vol. 19, no. 3. — P. 497—520. — doi:10.1080/02724634.1999.10011161.
- Holtz, 2004, p. 113.
- Xu Xing, Mark A. Norell, Kuang Xuewen, Wang Xiaolin, Zhao Qi, & Jia Chengkai. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids (англ.) // Nature. — 2004. — Vol. 431, no. 7009. — P. 680—684. — doi:10.1038/nature02855.
- Hurum & Sabath, 2003, p. 163.
- Masato Watabe & Shigeru Suzuki. Cretaceous fossil localities and a list of fossils collected by the Hayashibara Museum of Natural Sciences and Mongolian Paleontological Center Joint Paleontological Expedition (JMJPE) from 1993 through 1998 // Hayashibara Museum of Natural Sciences Research Bulletin. — 2000. — Vol. 1. — P. 99—108.
- Philip J. Currie. Nomadic Expeditions, Inc., report of fieldwork in Mongolia, September 2000 // Alberta Palaeontological Society, Fifth Annual Symposium, Abstract Volume. — Calgary: Mount Royal College, 2001. — P. 12—16.
- Philip J. Currie. Report on fieldwork in Mongolia, September 2001 // Alberta Palaeontological Society, Sixth Annual Symposium, "Fossils 2002," Abstract Volume. — Calgary: Mount Royal College, 2002. — P. 8—12.
- Курзанов С. М. Новый позднемеловой карнозавр из Ногон-Цава, Монголия // Палеонтология и биостратиграфия Монголии / Труды ССМПЭ. — 1976. — Т. 3. — С. 93—104.
- Dong Zhiming. On the dinosaurian remains from Turpan, Xinjiang (кит.) // Vertebrata PalAsiatica. — 1977. — Vol. 15. — P. 59—66.
- Philip J. Currie & Dong Zhiming. New information on Shanshanosaurus huoyanshanensis, a juvenile tyrannosaurid (Theropoda, Dinosauria) from the Late Cretaceous of China // Canadian Journal of Earth Sciences. — 2001. — Vol. 38, № 12. — P. 1729—1737. — doi:10.1139/cjes-38-12-1729.
- Takanobu Tsuhiji, Mahito Watabe, Khishigjav Tsogtbaatar, Takehisa Tsubamoto, Rinchen Barsbold, Shigeru Suzuki, Andrew H. Lee, Ryan C. Ridgely, Yasuhiro Kawahara, & Lawrence M. Witmer. Cranial Osteology of a Juvenile Specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia // Journal of Vertebrate Paleontology. — 2011. — Vol. 31, № 3. — P. 497—517. — doi:10.1080/02724634.2011.557116. Архивировано 6 октября 2011 года.
- Paul C. Sereno, Lin Tan, Stephen L. Brusatte, Henry J. Kriegstein, Xijin Zhao, & Karen Cloward. Tyrannosaurid Skeletal Design First Evolved at Small Body Size // Science. — 2010. — Vol. 326, № 5951. — P. 418—422. — doi:10.1126/science.1177428.
- James Clark. Becoming T. rex // Science. — 2010. — Vol. 326, № 5951. — P. 373—374. — doi:10.1126/science.1181276.
- Denver W. Fowler, Holly N. Woodward, Elizabeth A. Freedman, Peter L. Larson, & John R. Horner. Reanalysis of "Raptorex kriegsteini": A Juvenile Tyrannosaurid Dinosaur from Mongolia // PloS ONE. — 2011. — Vol. 6, № 6. — PMID 21738646.
- Jerzykiewicz, Tomasz, & Russell, Dale A. Late Mesozoic stratigraphy and vertebrates of the Gobi Basin // Cretaceous Research. — 1991. — Vol. 12, № 4. — P. 345—377. — doi:10.1016/0195-6671(91)90015-5.
- Зал V (Сектор В): Мезозой, динозавры и их современники. Палеонтологический институт РАН (2001). Архивировано 25 октября 2012 года.
- Adam T. Halamski. Mongolian collection (англ.). Institute of Paleobiology, Polish Academy of Sciences. Дата обращения: 20 сентября 2011. Архивировано 23 января 2012 года.
- Casey Hynes. The outer limits of Ulaan Baatar (англ.) (недоступная ссылка). Global Times (1 августа 2011). Дата обращения: 20 сентября 2011. Архивировано 6 октября 2012 года.
- Philip J. Currie. Paleontological Museum of the Mongolian Academy of Sciences, Ulaan Baatar // Encyclopedia of Dinosaurs / Philip J. Currie.. — Academic Press, 1997. — P. 524—525. — ISBN 0080494749.
- Ж. Болор. Т-БАТААР СҮХБААТАРЫН ТАЛБАЙД ЗАЛАРЛАА
- Tarbosaurus bataar — a ferocious theropod (англ.). Melbourne Museum. Дата обращения: 20 сентября 2011. Архивировано 23 января 2012 года.
- Hurum & Sabath, 2003, p. 161.
- Gregory S. Paul. The extreme lifestyles and habits of the gigantic Tyrannosaurid superpredators of the Late Cretaceous of North America and Asia. // Tyrannosaurus rex, the tyrant king / Peter L. Larson, & Kenneth Carpenter (eds.). — Bloomington, IN: Indiana University Press, 2008. — P. 307—352. — ISBN 978-0-253-35087-9.
- Gregory S. Paul. Tyrannosaurids // The Princeton Field Guide to Dinosaurs. — 2nd ed. — Princeton, NJ: Prnceton University Press, 2016. — P. 108—117. — ISBN 978-0-691-16766-4.
- Holtz, 2004, p. 132.
- Philip J. Currie. Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia (англ.) // Canadian Journal of Earth Sciences. — 2003-04-01. — Vol. 40. — Iss. 4. — P. 651—665. — ISSN 1480-3313 0008-4077, 1480-3313. — doi:10.1139/e02-083.
- François Therrien, Donald M. Henderson. My Theropod is Bigger Than Yours ... or Not: Estimating Body Size from Skull Length in Theropods (англ.) // Journal of Vertebrate Paleontology. — 2007. — Vol. 27. — Iss. 1. — P. 108—115. — doi:10.2307/4524671.
- Cranial Osteology of a Juvenile Specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia (PDF Download Available) (англ.). ResearchGate. Дата обращения: 27 октября 2017.
- Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared - Acta Palaeontologica Polonica (англ.). www.app.pan.pl. Дата обращения: 27 октября 2017.
- Zofia Kielan-Jaworowska. In Pursuit of Early Mammals. — Indiana University Press, 2013-07-12. — P. 47. — ISBN 9780253008244.
- David Lambert. A field guide to dinosaurs. — Rev. ed. — Avon Books, 1983. — P. 89. — 256 p. — ISBN 0380835193.
- Thomas R. Holtz Jr. The phylogeny and taxonomy of the Tyrnannosauridae // Mesozoic Vertebrate Life / Carpenter K., Tanke D. H., & Skrepnick M. W.. — Indiana University Press, 2001. — P. 71. — 577 p. — ISBN 0-253-33907-3.
- Holtz, 2004, p. 124.
- Peter Larson. Variation and sexual dimorphism in Tyrannosaurus rex. // Tyrannosaurus rex, the tyrant king / Peter L. Larson, & Kenneth Carpenter (eds.). — Bloomington, IN: Indiana University Press, 2008. — P. 103—128. — ISBN 978-0-253-35087-9.
- Hurum & Sabath, 2003, pp. 167—172.
- Малеев Е. А. О головном мозге хищных динозавров // Палеонтологический журнал. — 1965. — № 2. — С. 141—143.
- Савельев С. В., Алифанов В. Р. Новое исследование мозга хищного динозавра Tarbosaurus bataar (Theropoda, Tyrannosauridae) // Палеонтологический журнал. — 2005. — Т. 41, № 3. — С. 47—55. — doi:10.1134/S0031030107030070.
- Christopher A. Brochu. A digitally-rendered endocast for Tyrannosaurus rex // Journal of Vertebrate Paleontology. — 2000. — Vol. 20, № 1. — P. 1—6. — doi:10.1671/0272-4634(2000)020[0001:ADREFT]2.0.CO;2.
- Международная стратиграфическая шкала (версия за август 2012) на сайте Международной комиссии по стратиграфии
- Shen Y. B. and Mateer, Niall J. An outline of the Cretaceous System in northern Xinjiang, western China // Aspects of Nonmarine Cretaceous Geology / Mateer, Niall J.; and Chen Peiji (eds.). — Beijing: China Ocean Press, 1992. — P. 49—77.
- Krzysztof Owocki, Barbara Kremer, Martin Cotte, Hervé Bocherens. Diet preferences and climate inferred from oxygen and carbon isotopes of tooth enamel of Tarbosaurus bataar (Nemegt Formation, Upper Cretaceous, Mongolia) (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2020. — Vol. 537. — P. 109190. — ISSN 0031-0182. — doi:10.1016/j.palaeo.2019.05.012.
- Ефимов М. Б. Обзор ископаемых крокодилов Монголии // Ископаемые рептилии Монголии / Труды совместной советско-монгольской палеонтологической экспедиции. — 1983. — Т. 24. — С. 76—95.
- Holtz, 2004, p. 134.
- Farlow J. O, Smith M. B, & Robinson J. M. Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex (англ.) // Journal of Vertebrate Paleontology. — 1995. — Vol. 15, no. 4. — P. 713—725.
- McNeill Alexander R. Dinosaur biomechanics (англ.) // Proceedings of the Royal Society B: Biological Sciences. — 2006. — Vol. 273. — P. 1849—1855.
- Thomas R. Holtz Jr. A critical reappraisal of the obligate scavenging hypothesis for Tyrannosaurus rex and other Tyrant dinosaurs // Tyrannosaurus rex, the tyrant king / Peter L. Larson, & Kenneth Carpenter (eds.). — Bloomington, IN: Indiana University Press, 2008. — P. 371—398. — ISBN 978-0-253-35087-9.
- Hone, D. W. E., & Watabe, M. New information on scavenging and selective feeding behaviour of tyrannosaurids (англ.) // Acta Palaeontologica Polonica. — 2010. — Vol. 55, no. 4. — P. 627—634.
- The Giant Claw: A 'Walking with Dinosaurs' Special (англ.) на сайте Internet Movie Database
- «Тарбозавр 3D» (англ.) на сайте Internet Movie Database
Литература
- Малеев, Е. А. Новый хищный динозавр из верхнего мела Монголии // Доклады Академии наук СССР. — 1955. — Т. 104, № 5. — С. 779—783.
- Hurum, J. H., & Sabath, K. Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared // Acta Palaeontologica Polonica. — 2003. — Vol. 48, № 2. — P. 161—190.
- Thomas R. Holtz. Tyrannosauroidea // The Dinosauria / Weishampel, David B., Dodson, Peter, & Osmólska, Halszka.. — 2nd ed. — Berkeley: University of California Press, 2004. — P. 111—136. — ISBN 0-520-24209-2.
| Таксономия |
|---|