Бромодоменсодержащий белок 4
Бромодоменсодержащий белок 4 (англ. Bromodomain-containing protein 4, BRD4) — хроматинсвязывающий белок, который у человека кодируется геном BRD4. Он узнаёт и связывается с остатками ацетилированного лизина в составе гистонов и других белков. BRD4 принимает участие в регуляции транскрипции и передаче эпигенетической информации при делении клетки[1]. В 2016 году было установлено, что BRD4 обладает собственной гистонацетилтрансферазной активностью[2].
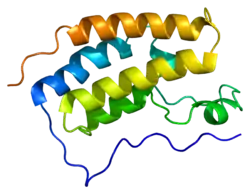
BRD4 принадлежит к семейству белков BET, содержащих два тандемных бромодомена и экстратерминальный домен. Кроме BRD4, к этому семейству относятся ещё три белка человека: BRD2, BRD3 и BRDT[3].
Распространение и внутриклеточная локализация
РНК BRD4 и белок присутствуют в подавляющем большинстве типов клеток и тканей[4][3].
BRD4 локализован главным образом в ядре клетки, но может присутствовать и в цитоплазме в небольшом количестве[1].
Структура гена и белка
В результате альтернативного сплайсинга формируются две изоформы белка. Более длинная и более распространённая изоформа состоит из 1362 аминокислотных остатков и имеет молекулярную массу около 152 кДа. Биологическую активность BRD4 связывают именно с этой изоформой. Вторая изоформа укорочена с C-конца, состоит из 723 аминокислотных остатков и имеет молекулярную массу 88 кДа. Короткую изоформу удаётся обнаружить гораздо реже, чем длинную, и её функции неизвестны[3][1].
BRD4 содержит два консервативных бромодомена на N-конце молекулы. Бромодомены состоят из четырёх α-спиралей, соединённых подвижными петлями. Вместе спирали и петли формируют гидрофобный карман, который распознаёт остатки ацетиллизина в белках[5].
C-концевой экстратерминальный домен BRD4 принимает участие в связывании фактора транскрипции P-TEFb[5].
Функции
Роль в транскрипции
BRD4 выступает в качестве глобального регулятора транскрипции, при этом он может действовать несколькими способами, пассивно и активно.
Транскрипция существенной доли генов многоклеточных организмов останавливается на стадии ранней элонгации, когда длина синтезированной пре-мРНК составляет около 60 нуклеотидов[6]. Одной из причин такой остановки является последовательное присоединение к транскрипционному комплексу негативных регуляторных факторов DSIF (англ. DRB sensitivity inducing factor) и NELF (англ. negative elongation factor), которые блокируют дальнейшее продвижение РНК-полимеразы II. Повторный запуск транскрипции осуществляется при участии фактора P-TEFb. P-TEFb представляет собой гетеродимер, состоящий из циклинзависимой киназы 9 и её регуляторной субъединицы — циклина T1, T2 или K. P-TEFb фосфорилирует DSIF и NELF и тем самым подавляет их активность. Кроме того, P-TEFb фосфорилирует гептадные повторы в С-концевом домене РНК-полимеразы II по положению Ser-2, что также способствует активной элонгации[5]. За привлечение P-TEFb в транскрипционный комплекс отвечает BRD4, который связывается с околопромоторными областями генов за счёт взаимодействия с остатками ацетиллизина в гистонах[7][8].
BRD4 является атипичной протеинкиназой: он способен самостоятельно фосфорилировать РНК-полимеразу II по положению Ser-2 при отсутствии явной структурной гомологии с другими протеинкиназами[9].
Роль в транскрипции отдельных генов
BRD4 стимулирует транскрипцию гена CD274, кодирующего белок PD-L1, независимо от MYC. На примере рака яичника и B-клеточной лимфомы было продемонстрировано, что применение ингибиторов BRD4, таких как JQ1, приводит к снижению количества PD-L1 на поверхности опухолевых клеток и вызывает T-клеточный иммунный ответ на них[10][11].
Роль в регуляции структуры хроматина
В 2016 году было установлено, что BRD4 является гистоацетилтрансферазой[2]. Он ацетилирует остатки лизина в N-концевых фрагментах гистонов 3 и 4, а также остаток Lys-122 в гистоне 3. Ацетилирование последнего остатка оказывает более существенное влияние на дестабилизацию нуклеосомы, потому что он находится в глобулярном C-концевом домене гистона и в непосредственном контакте с ДНК. Известны ещё только две гистоацетилтрансферазы, способные ацетилировать Lys-122 в гистоне 3: p300 и CBP. Гистонацетилтрансферазная активность BRD4 вызывает декомпактизацию хроматина (а следовательно, стимулирует транскрипцию) во многих участках генома, в том числе в районе генов MYC, FOS и AURKB.
Роль в заболеваниях человека
Впервые особая роль BRD4 в заболеваниях человека была описана при анализе редкого типа карциномы, характеризующегося хромосомной транслокацией t(15;19)[12]. При данной транслокации происходит слияние первой половины гена BRD4 и практический всей кодирующей области гена NUT[13]. В результате формируется химерный онкобелок BRD4-NUT, который отвечает за необычно агрессивный характер данной карциномы[14].
Примечания
- BRD4 в базе данных UniProt. Дата обращения: 11 сентября 2014.
- Devaiah B. N., Case-Borden C., Gegonne A., Hsu C. H., Chen Q., Meerzaman D., Dey A., Ozato K., Singer D. S. BRD4 is a histone acetyltransferase that evicts nucleosomes from chromatin. (англ.) // Nature structural & molecular biology. — 2016. — doi:10.1038/nsmb.3228. — PMID 27159561.
- Wu S. Y., Chiang C. M. The double bromodomain-containing chromatin adaptor Brd4 and transcriptional regulation. (англ.) // The Journal of biological chemistry. — 2007. — Vol. 282, no. 18. — P. 13141—13145. — doi:10.1074/jbc.R700001200. — PMID 17329240.
- BRD4 на сайте Human Protein Atlas. Дата обращения: 11 сентября 2014.
- Shi J., Vakoc C. R. The mechanisms behind the therapeutic activity of BET bromodomain inhibition. (англ.) // Molecular cell. — 2014. — Vol. 54, no. 5. — P. 728—736. — doi:10.1016/j.molcel.2014.05.016. — PMID 24905006.
- Kwak H., Lis J. T. Control of transcriptional elongation. (англ.) // Annual review of genetics. — 2013. — Vol. 47. — P. 483—508. — doi:10.1146/annurev-genet-110711-155440. — PMID 24050178.
- Jang M. K., Mochizuki K., Zhou M., Jeong H. S., Brady J. N., Ozato K. The bromodomain protein Brd4 is a positive regulatory component of P-TEFb and stimulates RNA polymerase II-dependent transcription. (англ.) // Molecular cell. — 2005. — Vol. 19, no. 4. — P. 523—534. — doi:10.1016/j.molcel.2005.06.027. — PMID 16109376.
- Yang Z., Yik J. H., Chen R., He N., Jang M. K., Ozato K., Zhou Q. Recruitment of P-TEFb for stimulation of transcriptional elongation by the bromodomain protein Brd4. (англ.) // Molecular cell. — 2005. — Vol. 19, no. 4. — P. 535—545. — doi:10.1016/j.molcel.2005.06.029. — PMID 16109377.
- Devaiah B. N., Lewis B. A., Cherman N., Hewitt M. C., Albrecht B. K., Robey P. G., Ozato K., Sims R. J. 3rd, Singer D. S. BRD4 is an atypical kinase that phosphorylates serine2 of the RNA polymerase II carboxy-terminal domain. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2012. — Vol. 109, no. 18. — P. 6927—6932. — doi:10.1073/pnas.1120422109. — PMID 22509028.
- Zhu H., Bengsch F., Svoronos N., Rutkowski M. R., Bitler B. G., Allegrezza M. J., Yokoyama Y., Kossenkov A. V., Bradner J. E., Conejo-Garcia J. R., Zhang R. BET Bromodomain Inhibition Promotes Anti-tumor Immunity by Suppressing PD-L1 Expression. (англ.) // Cell reports. — 2016. — Vol. 16, no. 11. — P. 2829—2837. — doi:10.1016/j.celrep.2016.08.032. — PMID 27626654.
- Hogg S. J., Vervoort S. J., Deswal S., Ott C. J., Li J., Cluse L. A., Beavis P. A., Darcy P. K., Martin B. P., Spencer A., Traunbauer A. K., Sadovnik I., Bauer K., Valent P., Bradner J. E., Zuber J., Shortt J., Johnstone R. W. BET-Bromodomain Inhibitors Engage the Host Immune System and Regulate Expression of the Immune Checkpoint Ligand PD-L1. (англ.) // Cell reports. — 2017. — Vol. 18, no. 9. — P. 2162—2174. — doi:10.1016/j.celrep.2017.02.011. — PMID 28249162.
- French C. A., Miyoshi I., Aster J. C., Kubonishi I., Kroll T. G., Dal Cin P., Vargas S. O., Perez-Atayde A. R., Fletcher J. A. BRD4 bromodomain gene rearrangement in aggressive carcinoma with translocation t(15;19). (англ.) // The American journal of pathology. — 2001. — Vol. 159, no. 6. — P. 1987—1992. — doi:10.1016/S0002-9440(10)63049-0. — PMID 11733348.
- French C. A., Miyoshi I., Kubonishi I., Grier H. E., Perez-Atayde A. R., Fletcher J. A. BRD4-NUT fusion oncogene: a novel mechanism in aggressive carcinoma. (англ.) // Cancer research. — 2003. — Vol. 63, no. 2. — P. 304—307. — PMID 12543779.
- French C. A. Pathogenesis of NUT midline carcinoma. (англ.) // Annual review of pathology. — 2012. — Vol. 7. — P. 247—265. — doi:10.1146/annurev-pathol-011811-132438. — PMID 22017582.