Альфа-мотонейрон
Альфа-мотонейроны (α-мотонейроны) — большие, мультиполярные нижние мотонейроны ствола мозга и спинного мозга. Они иннервируют экстрафузальные мышечные волокна скелетных мышц и напрямую ответственны за инициализацию их сокращения. Альфа-мотонейроны отличаются от гамма-мотонейронов, которые иннервируют интрафузальные мышечные волокна мышечных веретен.
В то время как тела альфа-мотонейронов располагаются в центральной нервной системе (ЦНС), α-мотонейроны также считаются частью соматической нервной системы — отдела периферической нервной системы (ПНС) — поскольку их аксоны идут в составе периферических нервов для иннервации скелетной мускулатуры.
Альфа-мотонейрон и мышечные волокна, которые он иннервирует, образуют моторную единицу. Пул мотонейронов объединяет все мотонейроны, иннервирующие одну мышцу.
Расположение
Альфа-мотонейроны, иннервирующие голову и шею располагаются в стволе головного мозга; α-МН, иннервирующие остальное тело, располагаются в спинном мозге. В спинном мозге α-МН больше чем в стволе мозга, т.к. количество α-МН прямо пропорционально точности контроля за работой отдельной мышцы. Например, мышцы пальцев имеют большее число α-МН на волокно, и большее общее число α-МН, чем четырёхглавая мышца бедра, что позволяет более тонко контролировать работу пальцев.
Обычно, α-МН, расположенные на одной стороне ствола мозга или спинного мозга, иннервируют мышцы, расположенные на этой же стороне тела. Исключением является ядра блокового нерва, расположенные в стволе мозга, которые иннервируют верхнюю косую мышцу глаза на противоположной стороне лица.
Ствол мозга
В стволе головного мозга, α-МН и другие нейроны находятся внутри скоплений клеток, называемых ядрами, некоторые из которых содержат тела нейронов, принадлежащих к черепным нервам. Не все ядра черепных нервов содержат α-МН; на этом основании ядра делят на двигательные и сенсорные.
Как правило, двигательные ядра, расположенные выше в стволе мозга (т.е., более рострально) иннервируют мышцы, расположенные выше на лице. Например, ядра глазодвигательного нерва, содержащие α-МН, которые иннервируют мышцы глаза, располагаются в среднем мозге — наиболее ростральном компоненте ствола мозга. С другой стороны, ядро подъязычного нерва, которое содержит α-МН, иннервирующие язык, находится в продолговатом мозге — наиболее каудальной (т.е. расположенном к низу) из стволовых структур.
Спинной мозг
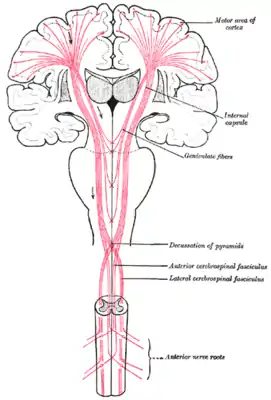
В спинном мозге, α-МН располагаются в сером веществе передних рогов. Эти α-МН обеспечивают моторный компонент спинномозговых нервов, которые иннервируют мышцы тела.
Как и в стволе мозга, вышележащие сегменты спинного мозга содержат α-МН, которые иннервируют мышцы, расположенные выше на теле. Например, двуглавая мышца плеча, мышца руки, иннервируется α-МН, расположенными в сегментах спинного мозга С5, С6 и С7, которые расположены в ростральной (верхней) части спинного мозга. С другой стороны, икроножная мышца, одна из мышц ноги, иннервируется α-МН, расположенными в пределах сегментов S1 и S2, которые находятся в каудальной (нижней) части спинного мозга.
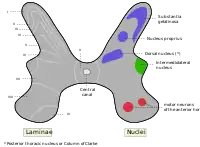
Альфа-мотонейроны располагаются в определенной области серого вещества спинного мозга. Эта область относится к пластине IX по системе пластин Рекседа, которая классифицирует регионы серого вещества на основе их цитоархитектоники. Пластина IX расположена преимущественно в медиальной части вентрального (переднего) рога, хотя некоторые её части лежат внутри VII и VIII пластин. Как и другие регионы спинного мозга, клетки этой пластины организованы соматотопически, это означает, что положение нейронов в спинном мозге взаимосвязано с тем, какие мышцы они иннервируют. В частности, α-МН в медиальной зоне пластины ІХ чаще иннервируют проксимальные мышцы тела, в то время как нейроны латеральной зоны обычно иннервируют более дистальные мышцы. Кроме того, имеется соматотропия связанная с α-МН, иннервирующими сгибатели и разгибатели: α-МН, иннервирующие мышцы-сгибатели как правило расположены в дорсальной части пластинки ІХ; те, что иннервируют разгибатели как правило расположены более вентрально.
Связи
Как и другие нейроны, нижние двигательные нейроны обладают как афферентными (входящими) и эфферентными (исходящими) связями. Альфа-мотонейроны получают афферентную иннервацию из ряда источников, в том числе верхних мотонейронов, сенсорных нейронов и интернейронов. В свою очередь α-МН иннервируют экстрафузальные мышечные волокна. Эти афферентные и эфферентные связи участвуют в координации мышечной активности.
Афферентный вход
| Расположение ВМН | Расположение α-МН | Имя пути |
|---|---|---|
| Головной мозг | Ствол мозга | Корково-ядерный путь |
| Головной мозг | Спинной мозг | Пирамидный тракт |
| Красное ядро | Спинной мозг | Красноядерно-спинномозговой тракт |
| Вестибулярные ядра | Спинной мозг | Преддверно-спинномозговой тракт |
| Крыша среднего мозга | Спинной мозг | Крышечно-спинномозговой тракт |
| Ретикулярная формация | Спинной мозг | Ретикулярно-спинномозговой тракт |
Верхние мотонейроны (ВМН) посылают аксоны к α-МН через несколько путей, включая (но не ограничиваясь) корково-ядерный, пирамидный и красноядерно-спинномозговой проводящие пути.
Корково-ядерный путь соединяет кору больших полушарий с ядрам черепных нервов. (Корково-ядерный путь также называют кортикобульбарным путём.) Он представляет собой пучок аксонов верхних двигательных нейронов, который спускается из коры и оканчивается синапсами на α-МН ствола мозга. Аналогично, ВМН коры головного мозга напрямую контролируют α-МН в спинном мозге через латеральные и вентральные корково-спинномозговые тракты.
Афферентный (сенсорный) вход для α-МН обширен и берет своё начало в сухожильных органах Гольджи, мышечных веретенах, механорецепторах, терморецепторах, и других сенсорных нейронах на периферии. Эти связи образуют структуру нейронных цепей, которые лежат в основе рефлексов. Существует несколько типов рефлекторных схем, простейшая из которых состоит из одного синапса между сенсорным нейроном и α-МН. Коленный рефлекс является примером такого моносинаптического рефлекса.
Наиболее значительная афферентация приходит на α-МН от локальных интернейронов, которые являются наиболее многочисленным типом нейронов в спинном мозге. Помимо множества других функций которые они выполняют, интернейроны посылают аксоны к α-МН, что создаёт более сложные рефлекторные схемы. Одним из типов интернейронов являются клетки Реншоу, которые будут рассмотрены ниже.
Эфферентный выход
Альфа-мотонейроны посылают аксоны, которые в основном оканчиваются синапсами на экстрафузальных мышечных волокнах. Некоторые аксоны α-МН оканчиваются на клетках Реншоу, т. е. тормозных интернейронах, которые посылают аксоны к α-МН и ограничивают их активность с целью предотвращения повреждения мышц.
Сигналы
Как и другие нейроны, α-МН передают сигналы в виде потенциалов действия, быстрых изменений электрической активности, распространяющихся от тела клетки к окончанию аксона. Чтобы увеличить скорость, с которой потенциалы действия распространяются, аксоны α-МН имеют большой диаметр и сильно миелинизированы как олигодендроцитами, так и Шванновскими клетками. Олигодендроциты миелинизируют часть аксона альфа-мотонейрона, которая находится в центральной нервной системе (ЦНС), тогда как Шванновские клетки миелинизируют ту часть, которая лежит в периферической нервной системе (ПНС). Переход между ЦНС и ПНС происходит на уровне мягкой мозговой оболочки.
Аксон α-МН соединяется с экстрафузальным мышечным волокном при помощи нервно-мышечного синапса, специализированного типа химического синапса, который отличается как по структуре, так и функциям от химических синапсов, соединяющих нейроны между собой. Оба вида синапсов используют нейротрансмиттеры для перевода электрического сигнала в химический сигнал и обратно. Однако они отличаются тем, что синапсы между нейронами, как правило, используют глутамат или ГАМК, в качестве нейротрансмиттеров, в то время как нервно-мышечные синапсы используют исключительно ацетилхолин. Ацетилхолин связывается с никотиновыми ацетилхолиновыми рецепторами на экстрафузальных мышечных волокнах, вызывая их сокращение.
Как и другие моторные нейроны, α-МН названы в соответствии со свойствами их аксонов. Альфа-мотонейроны имеют аксоны Аα-типа, большого диаметра, сильно миелинизированые, обладающие большой скоростью проведения потенциалов действия. Напротив, гамма-мотонейроны имеют аксоны типа Aγ, тонкие, маломиелинизированные, с малой скоростью проведения.
Роль в заболеваниях
Поражение α-МН является наиболее распространенным типом дисфункций нижних (периферических) мотонейронов. Повреждение может быть вызвано травмой, ишемией, инфекцией и др. Кроме того, некоторые заболевания связаны с избирательной потерей α-МН. Например, полиомиелит вызывается вирусом, который убивает мотонейроны в передних рогах спинного мозга. Боковой амиотрофический склероз также связан с гибелью мотонейронов.
Паралич является одним из наиболее выраженных эффектов повреждения α-МН. Поскольку только α-МН обеспечивают произвольную иннервацию экстрафузальных мышечных волокон, потеря α-МН эффективно разрушает связь между стволом мозга, спинным мозгом и мышцами, которые они иннервируют. Без этой связи, произвольный и непроизвольный (рефлекторный) контроль над мышцами невозможен. Произвольный контроль над мышцами теряется, поскольку α-МН переключают произвольную иннервацию от верхних мотонейронов к мышечным волокнам. Потеря непроизвольного контроля ведет к прерыванию рефлекторных дуг, таких как тонический рефлекс растяжения. Следствием прерывания рефлекторных дуг является снижение мышечного тонуса, в результате чего возникают вялые парезы. Другим следствием является угнетение глубоких сухожильных рефлексов, вызывая арефлексию.
Мышечная слабость и атрофия также являются неизбежными последствиями поражения α-МН. Поскольку размер мышц и их сила связаны со интенсивностью их использования, денервированные мышцы склонны к атрофии. Вторичной причиной атрофии мышц является то, что к денервированным мышцам больше не поступают трофические факторы из иннервирующих их α-МН. Поражение альфа-мотонейронов также проявляется на электромиограмме (например, в виде потенциалов фибрилляции), а также в виде фасцикуляций - спонтанных, непроизвольных сокращений мышц.
Заболевания, которые ухудшают передачу сигнала между α-МН и экстрафузальными мышечными волокнами, называемые болезнями нервно-мышечного соединения, имеют схожие признаки с теми, которые возникают при заболеваниях α-МН. Например, миастения является аутоиммунным заболеванием, которое препятствует передаче возбуждения по нервно-мышечным синапсам, что приводит к функциональной денервации мышц.
Развитие
Альфа-мотонейроны происходят из базальной пластинки, расположенной в вентральной части нервной трубки развивающегося эмбриона. Белок "ёжик Соник" (SHH) секретируется близлежащей хордой и другими вентральными структурами (например, пластинкой дна), при этом создаётся градиент с высокой концентрацией SHH в базальной пластинке и меньшей концентрацией SHH в крыловидной пластинке. Под влиянием SHH и других факторов, некоторые нейроны из базальной пластинки дифференцируются в α-МН.
Как и другие нейроны, α-МН посылают аксоны, к своим целевым экстрафузальным мышечным волоконам используя аксональное наведение, процесс в регуляции которого принимают участие нейротрофические факторы, которые секретируются целевым мышечным волокном. Нейротрофические факторы также гарантируют, что каждое мышечное волокно иннервируется соответствующим числом α-МН. Как и большинство типов нейронов в нервной системе, α-МН более многочисленны в начале развития, по сравнению со взрослым периодом. Мышечные волокна выделяют ограниченное количество нейротрофических факторов, способных поддерживать лишь незначительную часть α-МН, из тех что изначально проецируются на мышечное волокно. Таким образом, α-МН, которые не получают достаточного количества нейротрофических факторов подвергаются апоптозу, одной из форм запрограммированной клеточной смерти.
Некоторые кластеры α-МН получают высокую концентрацию нейротрофических факторов, так как они иннервируют множество мышц, следовательно, они выживают на этом этапе развития эмбриона. Это верно для α-МН, иннервирующих верхние и нижние конечности: эти α-МН располагаются в шейном и пояснично-крестцовом утолщениях спинного мозга. В свою очередь, α-МН также секретируют ряд трофических факторов для поддержания мышечных волокон, которые они иннервируют. Снижение уровня трофических факторов способствует атрофии мышц, которая следует за поражением α-МН.
См. также
- Бета-мотонейрон
- Экстрафузальное мышечное волокно
- Гамма-мотонейрон
- Интрафузальное мышечное волокно
- Нервно-мышечное веретено
- Клетка Реншоу
Литература
- John A. Kiernan. Barr's the Human Nervous System: An Anatomical Viewpoint (англ.). — 8th. — Hagerstown, MD: Lippincott Williams & Wilkins, 2005. — ISBN 0-7817-5154-3.
- Duane E. Haines. Neuroanatomy: An Atlas of Structures, Sections, and Systems (англ.). — 6th. — Hagerstown, MD: Lippincott Williams & Wilkins, 2004. — ISBN 0-7817-4677-9.
Ссылки
- NIF Search - Alpha Motor Neuron via the Neuroscience Information Framework