Дендритная пластичность
Дендритная пластичность — это характерный для ЦНС фундаментальный механизм, который лежит в основе синаптической потенциации и является ключевым для формирования памяти, обучения и когнитивных способностей, для нормального функционирования мозга.
В основе дендритной пластичности лежит динамическая природа дендритов, которые могут быть смоделированы через изменение их объёма, наклона, с потерей или добавлением других дендритов и дендритных шипиков, изменениями длины шеи шипиков, действием на дендритные потенциал-зависимые ионные каналы. Все эти формы дендритной пластичности связаны с обучением и памятью и являются основой уникальных биологических, вычислительных функций одиночных нейронов.
Влияние дендритов на нейронную пластичность
Хотя способность нейронных дендритов к генерации локальных потенциалов действия (дендритных спайков) известна уже несколько десятилетий, относительно мало внимания уделялось способности дендритов стимулировать синаптическую пластичность. Впрочем, поскольку синапсы (особенно возбуждающие) больше всего сосредоточены в дендритах, то дендритные свойства должны существенно влиять на появление и формирование различных типов синаптической и несинаптической (внутренней) пластичности.
Изменения в морфологии дендритов во взрослом мозге происходят очень медленно. К примеру, длительная визуализация дендритов у мыши не показала каких-либо добавлений или удалений дендритных ветвей на протяжении одного месяца[1]. Поэтому считается, что не столько сами дендриты, сколько дендритные шипики являются элементарными единицами хранения памяти и формирования синаптической пластичности, обусловленной изменениями в синаптической эффективности через долговременную потенциацию (ДВП) и долговременное ослабление (или долговременную депрессию, ДВД)[2]. В отличие от дендритов, шипики непрерывно и спонтанно формируются в мозге, двигаясь, вырастая и отходя на протяжении нескольких минут, а их размер коррелирует с их функциональной синаптической эффективностью, как in vivo, так in vitro[3][4][5].
Прогресс в технологии in vivo нейровизуализации позволил выявить пластические изменения в нейрональных свойствах, связанных с обучением и памятью[6]. Такие изменения включают перестройку в морфологии уже существующих синапсов, в частности увеличение дендритных шипиков в течение ДВП и уменьшение шипиков течение ДВД[7]. Другие структурные изменения могут включать появление новых синаптических контактов, обусловленных появлением новых дендритных шипиков вследствие стимуляции синаптической пластичности. Одним из доказательств того, что подобные структурные изменения связаны с памятью и обучением, является недавнее исследование, которое показало рост дендритных шипиков в нейронах двигательной коры как следствие двигательного обучения[8].
Независимо от механизмов стимулирования, важным признаком наличия синаптической пластичности является увеличение внутриклеточной концентрации кальция в течение стимулирования[9][10][11]. От амплитуды и длительности постсинаптического временного увеличения кальция в течение стимулирования зависит то, усилится или ослабится синаптическая передача. Длительное и умеренное увеличение кальция ведет к ДВД, тогда как короткое и сильное увеличение — к ДВП[12]. Активация только одного возбуждающего синапса ведёт к накоплению кальция в индивидуальных дендритных шипиках, привлекая для этого NMDA-рецепторы, которые благодаря современным достижениям в технологии нейровизуализации могут быть увидены и измерены in vivo[13][14][15]. Впрочем, на сегодняшний день количественных оценок кальциевых импульсов, лежащих в основе этих механизмов, не получено.
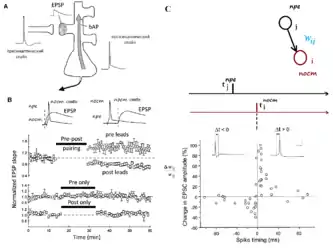
А. Стимуляция STDP путём синхронизации пресинаптических спайков и ВПСП (EPSP) с постсинаптическими спайками. bAP = потенциал действия обратного распространения.
B. «Пре-пост» спайки стимулируют ДВП, а «пост-пре» спайки стимулируют ДВД. Сами по себе пре- или пост-синаптические спайки не меняют синаптическую силу (вес).[16]
С. STDP: изменение синаптических связей как функция относительной синхронизации пре- и пост синаптических спайков после повторяющейся корреляции 60 спайков (с частотой 1 Гц)[17]
Пластичность Хэбба
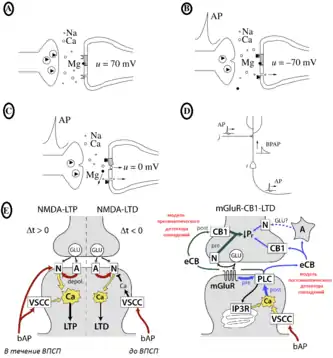
A. Везикулы в пресинаптических окончаниях содержат глутамат как нейромедиатор (заполненные треугольники). В состоянии потенциала покоя, канал NMDA-рецепторов блокируется магнием.
B. Когда потенциал действия (AP) прибывает в пресинаптическое окончание, везикула присоединяется к мембране клетки, глутамат входит в синаптическую щель и связывает NMDA и не-NMDA рецепторы на постсинаптической мембране. В состоянии потенциала покоя канал NMDA-рецепторов остаётся заблокированным магнием, тогда как не-NMDA каналы открываются (внизу).
C. Когда мембрана постсинаптического нейрона деполяризуется, то прекращается блокировка магния, и кальциевые ионы входят в клетку.
D. Деполяризация постсинаптической мембраны может быть вызвана обратным распространением потенциала действия (BPAP)[18]
E.Клеточные механизмы зависимой от времени пластичности. Биохимические сигнальные пути для большинства форм STDP (англ. Spike-timing dependent plasticity, рус. Модель пластичности, зависимая от времени спайка). Красным указана деполяризация. Для mGluR-CB1-LTD пресинаптический детектор совпадений показан зелёной стрелкой, а постсинаптический — синей. (mGluR = метаботропный глутамат-рецептор; CB = каннабиноиды; N и A = NMDA- и AMPA-рецепторы; A — астроцит)[16]
Открытие потенциалов действия обратного распространения было одним из первых свидетельств синаптической пластичности в дендритах, поскольку они делают возможными временную координацию и выявление совпадений[19][20]. Результаты многих исследований подтверждают, что Ca2+ импульсы, связанные с bAP и синаптическими входными импульсами, суммируются суперлинейно, когда синхронность двух событий происходит в достаточно узком временном интервале (<50 мкс)[21]. Это взаимодействие между входом и выходом позволяет спайкам, которые следуют за синаптическим входным импульсом в течение нескольких десятков микросекунд (мкс), сгенерировать постсинаптический Ca2+ импульс, который является значительно большим, чем если бы спайк предшествовал входному импульсу или слишком сильно отставал от него (>50 мкс). Эта временная синхронность обусловливается НМДА-рецептором и потенциалзависимыми каналами и является одним из механизмов пластичности, зависящим от времени спайка (англ. spike-timing dependent plasticity, STDP), что является расширенной версией модели Хэбба для синаптической пластичности[22][23].
Правило обучения Хэбба требует выявления совпадений в пресинаптической и постсинаптической активности[24]. Ответ детектора совпадений в присутствии обоих импульсов должен быть качественно отличным от ответа к каждому из этих импульсов отдельно. Как было упомянуто перед этим, лучше всего на роль такого детектора подходит уникальный макромолекулярный комплекс — NMDA-рецептор (NMDA-R), который может лежать в основе ДВП. Имея ряд необычных биофизических свойств, NMDA-R открывается только когда оба, и пресинаптические и постсинаптические нейроны, активированы. То есть, открытие рецептора требует и пресинаптического химического импульса (глутамат-нейромедиатора), и постсинаптического электрического импульса (локальную мембранную деполяризацию). Когда эти два условия выполняются в то же время, открытие NMDA-R позволяет Ca2+ ионам войти в постсинаптический нейрон и активировать Ca2+-восприимчивые ферменты, которые в конечном итоге и приводят к синаптическому усилению или ослаблению.
Как и другой ионотропный рецептор, AMPA-рецептор, NMDA-R активируется возбуждающим нейромедиатором глутаматом, но его восприимчивость к этому медиатору значительно большая (EC50 в диапазоне 1 мкм), что делает этот рецептор более восприимчивым к его меньшим концентрациям. Он также имеет более медленную частоту деактивации после кратковременного применения глутамата, что означает, что будучи один раз пресинаптически высвобожденным, глутамат связывается с рецептором и остаётся соединённым с ним в течение долгого времени. Благодаря этому NMDA-R действует как долговременный индикатор пресинаптической активности[25]. Именно эти свойства делают NMDA-рецептор идеальной основой для синаптической пластичности (напр., STDP) и выявления совпадений[9][26].
Кластерная пластичность
Поскольку внутриклеточная концентрация кальция является необходимым условием для возникновения синаптической пластичности, то синаптически порождённые локальные дендритные спайки могут быть одной из причин синаптической пластичности, и даже сами по себе вызывать ДВП и ДВД. Так, к примеру, в одном из недавних исследований[11] было сделано неожиданное открытие, что синаптически вызванные дендритные спайки стимулируют ДВП в CA1 нейронах гиппокампа, однако эта синаптическая пластичность не требует активного обратного распространения натриевых соматических потенциалов действия, поскольку они блокируются локальным применением к ближайшей части апикального дендрита антагониста натриевого канала — TTX. Скорее всего, формирование прочной ДВП в отдалённых дендритах требует относительно сильной стимуляции и, как следствие, локальных дендритных спайков, а не ослабленного потенциала обратного распространения.
В этом же исследовании[11] было показано, что ДВП в близких дендритах также может быть вызвана локальными дендритными спайками при отсутствии соматических потенциалов действия. Значит, стимуляция пластичности не обязательно должна быть связана с аксонным возвратным потенциалом действия[27]. А поскольку пространственно вместе локализованные синаптические входные импульсы имеют большую вероятность вызвать локальные дендритные спайки и ДВП, из этого следует важный вывод, что аксоны пресинаптических нейронов с коррелированной активностью образуют синаптические кластеры в дендритном дереве, к которому посылают импульсы. Подобная синаптическая кластеризация (т. н. «гипотеза кластерной пластичности») была теоретически предсказана ещё до её открытия, как эффективный способ обработки и хранения информации нейроном[28][29].
Кластерная пластичность может связывать вместе функционально объединённые входные импульсы, которые поступают к дендритам, и увеличивать в индивидуальных нейронах способность хранения путём привлечения нелинейных потенциал-зависимых каналов[30][31]. Более того, кластерная пластичность может увеличивать вероятность генерации локального спайка через усиление дендритной возбудимости[32], что в свою очередь укрепляет связь между дендритной ветвью и сомой[33]. Подобное увеличение силы дендритной ветви делает возможным порождение точного по времени и надёжного соматического выходного сигнала, являясь основой достоверной передачи информации на уровне единичных нейронов[34].
С появлением технологии двух-фотонного высвобождения глутамата возникла возможность точного направления глутамата к множеству или даже одному шипику. Применение этой техники к дендритам обнаружило бимодальное распределение дендритных ветвей: одна популяция ветвей со слабыми ответами на синхронные синаптические стимуляции, а другая — с более сильными[33]. То есть, информация в нейронах может храниться не только в форме синаптических весов входных синапсов, а дендритные ветви способны активно выявлять специфические свойства входных сигналов, напр., их синхронность.
Более того, внутренние свойства дендритных ветвей могут варьироваться в результате активности и в течение накопления опыта. Недавнее уникальное исследование[33], — новизна результатов которого породила целый ряд новых гипотез и моделей дендритной пластичности, — показало, что при одновременной стимуляции кластеров синапсов на дендритной ветке, их повторная активация приводит к постепенным изменениям в ответе мембраны на стимулы. В частности, было выявлено, что в CA1 пирамидальных нейронах гиппокампа появление холинергического агониста приводит к тому, что «слабые» ветви превращаются в ветви с сильной спаечной активностью путём сочетания синаптического возбуждения с постсинаптическим потенциалом действия. Такое превращение слабой дендритного ветви значительно усиливает распространение локальных дендритных Na+ спайков к соме и обусловлено новой формой пластичности — «branch-strength potentiation», (потенциация силы ветви, BSP), которая способна значительно увеличивать напряжение в соме по сравнению с непотенциированными ветвями. Увеличение напряжения, которое возникает в результате усиления распространения дендритного спайка вдоль ветки, скорее всего, является следствием подавления активности потенциал-зависимых калиевых (Kv4.2) каналов, которые ограничивают возбудимость дендритной ветви[33].
Эта специфическая для дендритной ветви пластичность, которая повышает её силу, превращает темпорально коррелированные и сгруппированные синаптические входные сигналы в долговременные изменения возбудимости ветви, будучи вероятным механизмом сохранения свойств входных сигналов, основой памяти.
Таким образом, связь между локальным дендритным спайком и сомой является пластичной и может быть изменена в специфический для данной ветки способ через обусловленную НМДА-рецептором регуляцию дендритных калиевых (Kv4.2) каналов. Тогда дендритная ветка предстаёт как единый интегративный сегмент («фундаментальной функциональной единицей в нервной системе»[35][36]) и влияние дендритного спайка на соматический потенциал будет существенно отличаться между отдельными дендритными ветвями.
Данные результаты были подтверждены этой же группой исследователей, которые обнаружили, что CA1 пирамидальные нейроны у животных, которые находились в обогащённой среде, проявляют облегчённое распространение дендритных спайков в подгруппе дендритных ветвей, пластично соединённых с сомой[37].
Хотя авторы этих исследований и выдвинули гипотезу, что потенциация дендритной ветки может лежать в основе долговременного сохранения паттернов входных сигналов, механизм этой пластичности оставался неизвестным. Только недавно была предложена детальная вычислительная модель[38], которая предсказала один из таких возможных механизмов и дала более обобщающее объяснение роли дендритов в нейронных вычислениях.
Эта модель впервые показала, как нейроны, благодаря дендритным спайкам и сочетанию классической (STDP) и неклассической (BSP) пластичности, способны выполнять сложные нелинейные вычисления путём самоорганизации. Оба механизма пластичности могут породить конкуренцию между дендритными ветвями, что позволяет одному нейрону, на клеточном уровне, выполнять такие нелинейные вычисления, которые ранее считались возможными только для нейронных сетей, напр. способность связывать множество входных признаков (т .н «проблема связывания», англ. binding problem). В частности, модель на примере нескольких десятков симуляций демонстрирует, что только при условии нелинейности дендритных вычислений нейрон сможет выполнять разделение образов (англ. pattern separation): двойная пластичность, вследствие конкуренции дендритов, позволяет сформировать образы в кластеры и затем, используя простые логические операции AND и OR, их классифицировать и связать (хотя такой нейрон и не в состоянии реализовать функцию XOR, для чего нужны ингибирующие синапсы)[38].
Итак, данная модель подтвердила гипотезы, которые возникли на основе экспериментов, что сила ветки может сохранять черты многих паттернов входных сигналов, помнить их[39].
Кроме найденных показаний in vitro исследований в пользу кластерной гипотезы[40], есть ряд исследований, которые показывают формирование кластеров синаптических входных импульсов in vivo, наиболее частое формирование соседних или дочерних шипиков в группе дендритов[41][42][43][44].
Так, вполне однозначно было показано, что активность дендритных шипиков часто синхронизируется в пределах группы шипиков, которые находятся близко друг к другу и объединены общими синаптическими входными сигналами от синхронизированных пресинаптических нейронов[45]. Кластерная синхронизация, которая возникнет в результате этого, может служить основой ассоциативного обучения на клеточном уровне, поскольку соседние шипики здесь кодируют разную информацию[15][46].
Одно из распространённых объяснений целесообразности синаптической кластеризации в дендритах заключается в том, что общая активация синапсов в пределах определённых дендритов приведёт к большей дендритной деполяризации, которая в свою очередь запустит регенеративные события, тем самым обеспечивая уверенный соматический ответ. Таким образом, синаптическая кластеризация может служить средством для гарантирования, что импульсы, которые состоят из множества одновременно активированных синапсов, надежно распространятся к соме или сохранятся в дендритах, тогда как некоррелируемые синапсы не будут иметь влияния на пластичность и потенциалы действия[47]. Хотя это довольно привлекательная гипотеза и она имеет ряд экспериментальных свидетельств, существуют исследования с совершенно противоположными результатами, когда входные импульсы не кластеризуются, но широко рассеяны по дендритному дереву и не сходятся в единый дендрит, линейно суммируясь[14].
Нехэббовская пластичность
Подобно тому, как внутренне нестабильной является классическая пластичность Хэбба, формы ДВП, которые опираются на дендритные локальные спайки, также будут дестабилизировать распределение нагрузок дендритного дерева[49]. Поэтому, как и в случае с хэббовской пластичностью, стойкость дендритной информации должна обеспечивать долговременная депрессия. Недавно было найдено свидетельство того, что НМДА-подобные спайки в неокортексе также способны вызвать ДВД, но к большому удивлению исследователей, эта форма ДВД для своего возникновения нуждается лишь в «одном выстреле» (т. н. однострельное ДВД) и не требует соматического потенциала действия[50], хотя, возможно, этот феномен может зависеть от определённого региона мозга. Такое «однострельное» синаптическое правило обучения кардинально отличается от классического протокола стимуляции ДВД, которое обычно включает сотни повторений, и должно опираться на совершенно иные механизмы, чем классические и синхронизационные формы ДВД. Принимая во внимание, как быстро вызывается это однострельное ДВД, исследователи считают, что оно может служить для других вычислительных целей, чем классические формы ДВД, например, быть основой для «однострельного» быстрого обучения[51].
Ещё более затруднило понимание природы дендритного высчисления открытие формы «однострельного» ДВП, хотя уже в CA1 нейронах гиппокампа. Этот тип ДВП требует только одной пачки (англ. burst) пресинаптической активности и зависит от локальных дендритных спайков. Относительно большие ВПСП (возбуждающий постсинаптический потенциал), использованные исследователями для стимуляции однострельного ДВП, предполагают, что несколько десятков пресинаптических CA3 пирамидальных нейронов были бы одновременно активированы in vivo с достаточно высокой временной точностью, что, впрочем, является вполне возможным в течение гиппокампных острых волн, которые видны на ЭЭГ[52]. Следовательно, открытие этого быстрого ДВП, даже если оно ограничивается только определённым регионом мозга, показывает скорость и неклассичность сугубо дендритного высчисления, которое лежит в основе сохранения информации, будучи вполне независимым от аксонного потенциала действия обратного распространения.
Поэтому, всё больше накапливается данных, что локальные дендритные спайки могут вызвать двустороннюю синаптическую пластичность. В отличие от STDP, которая требует совпадения между пре- и постсинаптическими спайками, обусловленная локальными спайками пластичность включает совпадение синаптических активаций тесно сгруппированных входных импульсов к одной и той же части дендрита[33]. Это является свидетельством нового, ещё неизвестного правила обучения, согласно которому нейроны делятся на крупные независимые модули интеграции и обучения, которые не зависят от привычного импульса обратной связи от сомы (bAP).
Важное различие между STDP и вызванной локальными спайками пластичностью заключается в скорости стимуляции: в то время, как STDP требует много повторений совпадающих пре-пост стимуляций[53], то лишь одного локального спайка достаточно для создания ДВД или ДВП[54] (однако, см. также[11]). Исследования CA1 пирамидальных нейронов обнаружили, что даже одна активация единственного синапса способна вызвать локальную деполяризацию, достаточную для активации НМДА-рецепторов[55]. Подобное увеличение в скорости стимуляции связано в значительной степени со взаимной активацией соседних НМДА-рецепторов, находящихся на одном дендрите. Следовательно, зависимая от локального спайка пластичность может быть решающим механизмом, который лежит в основе быстрого получения воспоминаний, рабочей памяти[56].
Дендритные вычисления
Избирательность дендритов в направлении
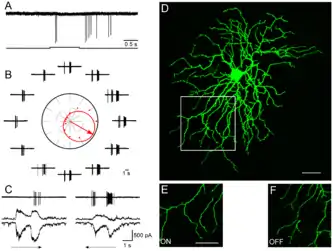
А. Ответ DSGC на появление и прекращение неподвижного вспыхивающего пятнышка.
В. Направленный ответ на треугольник, который движется в 12 направлениях. Направление, которое клетка предпочитает, показано как стрелка в середине полярных координат и соответствующими потенциалами действия.
С. Асимметричные синаптические входные импульсы, регистрируемые, когда мембранный потенциал был между −65 мВ и 0 мВ соответственно. Значительно больший ингибирующий импульс зарегистрирован, когда визуальный стимул двигался в NULL направлении.
D. Дендритная морфология клетки. Сегмент дендритного поля был увеличен, чётко показывая ON (E) и OFF (F) разделение дендритов.[57]
.png.webp)
Одним из наиболее ярких и наиболее изученных примеров того, насколько сложными могут быть дендритные вычисления, является избирательность к направлению (англ. directional selectivity, DS). Избирательные в направлении нейроны способны реагировать на движение изображений в одном направлении (PREF), но не в другом (NULL). Эти нейроны существуют у многих видов, от глаз мух до коры млекопитающих, и в большинстве из них проявляется активная роль дендритных вычислений.
Одно из первых убедительных экспериментальных подтверждений[59] привлечения дендритов в избирательность направления было сделано на зрительной системе мухи in vivo, в которой существует популяция больших интернейронов, т. н. тангенциальных клеток (TC), которые пространственно интегрируют выходные импульсы многих тысяч цилиндрических клеток, каждая из которых чувствительна только к очень малой части зрительного поля. Все TC клетки восприимчивы к движению: они возбуждаются движением в одном направлении и тормозятся движением в противоположном направлении. Соединив внутриклеточную регистрацию и кальциевую визуализацию дендритов in vivo, исследователи обнаружили два главных шага в обработке информации, которые выполняют дендриты TC клеток. Благодаря обработке входных элементов противоположных клеток, имеющих другие преимущественные направления, направленная избирательность пресинаптических нейронов в ТС клетках является существенно расширенной. В этом же исследовании было также показано, что дендритная фильтрации помогает отличить изменение в контрасте вследствие движения стимула от изменений, связанных с чисто локальными формами стимула. Результатом такой интеграции стимулов является постепенная деполяризация в аксоне тангенциальных клеток, которая с высокой точностью предоставляет информацию о скорости изображения[59].
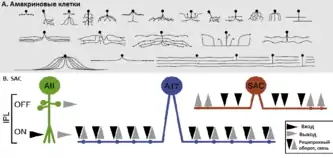
Классической моделью изучения механизмов дендритного высчисления направленной избирательности является исследование «избирательности в направлении ганглионарных клеток сетчатки» (англ. Direction-Selective Retinal Ganglion Cell, DSRGC или DSGC)[60]. Среди ганглиозных клеток направленно-выборочными являются т. н. ON/OFF ганглиозные клетки. Они действуют, как локальные детекторы движения, реагируя на двигающиеся точки, решётки и линии в широком диапазоне скоростей. При появлении или исчезновении стимула, ON возбуждаются, когда он попадает в центр их рецептивного поля, и ингибируются, когда на периферию; OFF клетки проявляют прямо противоположную реакцию. ON/OFF DS клетки имеют двухслойное дендритное дерево, с одним разветвлением в наружной половине (OFF sublamina) внутреннего сплетениевидного слоя (IPL), а другое ветвление — во внутренний половине IPL (ON sublamina)[61].
Такая организация DSGC позволяет отвечать на направление движения изображения тёмных объектов на светлом фоне — выполняется OFF ветвью — как и на объекты, более светлые, чем фон — осуществляется ON ветвью. Существуют четыре подтипа DS клетки, каждый из которых предпочитает одно конкретное направление движения. Каждый подтип покрывает сетчатку таким образом, что образуется небольшое перекрытие дендритов, таким образом получая информацию о направлении для каждого из преобладающих направлений, которые доступны в любом регионе сетчатки[62].
Вилфрид Ролл в 1969 году предложил первую простую модель того, как дендриты могут воплощать подобную DS[63]. Пока же идут дебаты относительно определения первичного уровня нейронов, который обнаруживает такую избирательность. Одни исследователи считают, что нелинейная взаимосвязь между возбуждением и ингибированием является вероятным механизмом для объяснения DS в ганглиозных клетках[64][65]. Другие исследователи, на основе теоретической модели, считают, что входной импульс к DSRGC уже является выборочным по направлению, а первично эта избирательность может быть вычислена в индивидуальных дендритных ветвях звёздчатых амакриновых нейронов (англ. starburst amacrine cells, SAC), которые являются пресинаптическими к ганглионарным клеткам сетчатки[66].
Если большинство классических теорий «направленной избирательности» рассматривали её происхождения как результат взаимодействия между нейронами, строя модели нейронных сетей, то с углублением изучения дендритов всё чаще возникают модели, которые предлагают рассматривать направленную избирательность на клеточном уровне как результат внутренних свойств нейронов. Одним из таких нейронов, где первично осуществляется избирательность, и считаются звёздчатые амакриновые клетки.
Амакриновые клетки лучше всего подходят для подобных вычислений движения. Состоя из 40 морфологически отличающихся типов, эти клетки являются крупнейшим классом интернейронов сетчатки[67]. Многие из них не имеют аксонов, а их дендриты служат как для получения входных синаптических импульсов, так и передачи выходных сигналов, поэтому они не являются дендритами в классическом понимании.
Наиболее характерную морфологию среди амакриновых клеток имеют SAC. Дендриты SAC расположены пучком вместе с DSGC-дендритами и именно поэтому рассматриваются как наиболее вероятные кандидаты на роль первичных детекторов движения. В отличие от других нейронов, SAC характеризуются огромным перекрытием дендритов и потому способны обеспечить различные подтипы DSGC надлежащими нейронными схемами вычислений[68]. Если, к примеру, SAC изъять из нейронной сети (напр., путём удаления клетки), то направленно-выборочные ответы в DSGC исчезают[69].
Благодаря почти электрически изолированным ветвям их дендритного дерева, SAC-дендриты реагируют независимо на световую стимуляцию[70]. Следовательно, иногда их даже предлагают рассматривать как «автономные» вычислительные единицы[71]: в отличие от целой клетки, дендриты является высоко поляризованными структурами; синаптические входные и выходные сигналы по-разному распределены вдоль дендритов — входные синапсы локализованы вдоль всей длины, тогда как выходные синапсы связаны с выступлениями на отдалённой трети ветвей. Каждая главная ветвь соответствует наиболее центробежному (в направлении дендритных окончаний), чем к центростремительному (в направлении сомы), движению, тем самым проявляя дендритную направленную избирательность[72]. С использованием двухфотонной оптической визуализации концентрации кальция в дендритах SAC было выявлено, что концентрация кальция на кончиках дендритов SAC действительно является выборочной в направлении[70].
.png.webp)
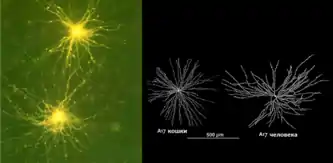
Изучение дендритной направленной избирательности позволило оценить, насколько в действительности сложными и нелинейными могут быть вычисления, которые выполняют дендриты. Новый свет на дендритную обработку информации пролило проведённое Уильямом Граймсом (Grimes) et al. новаторское исследование т. н. A17 амакриновых клеток в сетчатке крысы, в которых были обнаружены сложные формы параллельных вычислений[73].
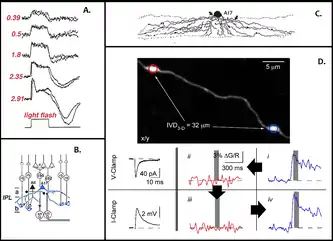
B. Формы связей А17 клеток с другими клетками сетчатки.
C. Морфология А17 клеток.
D. Экспериментальное подтверждение существования изолированных, параллельных вычислительных функциональных субъединиц в А17 клетках. Объединённый эксперимент с парой варикоз дендритов А17 клетки, удалённых на ∼30 мкм. Синаптические Ca2+ концентрации были вызваны в одном варикозе в условиях локальной фиксации потенциала (синяя кривая, i). Затем проверялась флуоресценция Ca2+ в ближайшем варикозе без изменения стимула (красная кривая, ii), однако вызванные Ca2+ транзиенты в них были редко заметны. После переключения на фиксацию тока, Ca2+ сигналы вновь были зарегистрированы в обоих варикозах (iii и iv). Левая панель показывает ВПСП (вверху) и ВПСП (внизу), вызванные синаптической стимуляцией. На основе[73].
Имея диаметр дендритного поля около 400 мкм в сетчатке крысы, A17 интернейроны относятся к подклассу амакриновых клеток. Их морфология почти одинакова для вида млекопитающих: они протягивают десятки тонких длинных (до 1 мм) дендритов с малыми варикозами, которые образуют реципрокные ГАМКергические синаптические обратные связи на аксонных окончаниях палочковых биполярных клеток[74]. Одна А17 амакриновая клетка получает синаптические входные (scotopic) импульсы от около 1000 палочковых биполярных клеток. Они не формируют синапсы с другими амакриновыми или ганглионарными клетками. Следовательно, А17, — в отличие соседних All-амакриновых клеток с узким рецептивным полем, — имеют совсем другую, более широкую площадь рецептивного поля, с которой они получают сигналы из палочек.
Граймс с коллегами, соединив двух-фотонную визуализацию кальция и вычислительное моделирование (см. изображение), неожиданно открыли, что даже одна A17 амакриновая клетка обеспечивает сетчатку примерно 150 локальными единицами (микросхемами) обратных связей, которые способны действовать совершенно независимо друг от друга, представая как отдельные дендритные компартменты. В частности, в сочетании с предыдущими данными[75] было выявлено, что поступление кальция в глутаматные рецепторы, активируемые биполярными клетками, усиливается высвобождением кальция из внутренних запасников клетки. Этого кальция вполне достаточно для того, чтобы вызвать реципрокное высвобождение ГАМК в аксонные окончания биполярных клеток без необходимости приведения в действие потенциал-зависимых кальциевых каналов. Так избегается значительное изменение мембранного напряжения, которое могло бы распространиться на соседние варикозы и помешать изоляции.
То есть, вместо того, чтобы интегрировать широкие пространственные сигналы, — как действуют дендриты во многих других нейронах, — большие А17 амакриновые клетки изолируют реципрокные синапсы обратной связи, и дендритные сегменты тогда действуют как параллельные, независимые субъединицы, каждая из которых вычисляется локально. На основе полученных данных исследователи построили модель, согласно которой дендритная ветка А17 может изолировать синаптические процессы в различных варикозах для потока фотонов, характерного для низко-освещенных (scotopic) условий. С увеличением интенсивности света (и потока фотонов), вероятность синхронных входных сигналов в соседних варикозах вдоль дендрита растёт, что ведёт к взаимодействиям, которые усиливают синаптический входной импульс.
Подобная дендритная сложность и широкое рецептивное поле А17 клеток может быть объяснено в контексте т. н. теории «оптимизации нейронных связей»[76] тем, что нейронная архитектура эволюционировала для обеспечения потребности в функциональных нейронных сетях, вместе с тем минимизируя «стоимость» связей (генетические, метаболические и пространственные ресурсы, необходимые для нейронных систем). Крупные нейроны (например в коре) могут распределять ресурсы вдоль большего числа синаптических связей, но они требуют больше энергии для транспортировки белков и имеют гораздо большее электротоническое затухание импульса. Но это не касается А17 клеток, которые используют скорее распределённое, параллельное вычисление, чем интегративное, как пирамидальные нейроны. Такие уникальные свойства дендритов А17 клеток, сохраняя пространственную остроту и динамический диапазон, лучше всего подходят именно для сетчатки, поскольку она для надлежащего выполнения своих функций должна быть достаточно тонкой и оптически прозрачной.
Дендритные вычисления в слуховой системе
Другой яркий пример влияния дендритов на нейронные высчисления был обнаружен при изучении слуховой системы, в частности её способности локализовать звук.
В отличие от других сенсорных систем, сенсорный эпителий внутреннего уха не имеет чёткой репрезентации пространства, поскольку волосковые клетки внутреннего уха кодируют частоту звука, а не звуковое пространство. Следовательно, звуковая система должна вычислять локализации источника звука в пространстве, для чего она использует два бинауральных сигнала, которые позволяют локализовать звуки в горизонтальной плоскости: интерауральную временную разницу (ИВР, англ. interaural time difference, ITD — разница во времени поступления звука до двух ушей) и интерауральную разницу интенсивностей (ИРИ, англ. interaural level difference, ILD). ИВР используются для низко-частотной (<2 кГц) локализации, а ИРИ для высоко-частотной[77].
Если звук поступает фронтально к голове, тогда ИВР = 0 мкс. Поскольку звуки с периферии (удалённые от срединно-сагиттальной плоскости) поступают в уши с большей временной задержкой, то они порождают разницы во время поступления стимула в два уха (начало ИВР) и в течение продолжительности стимула (продолжающаяся ИВР). Даже в наиболее выраженном горизонтальном положении звука, ИВР являются слишком малыми: 700 мкс у человека, 400 мкс у котов и 135 мкс у песчанки. Человек может различить ИВР с 10-20 мкс для низкочастотных звуков, и ИРИ ~1-2 дБ[78].
Подобная точность в кодировании источника звука и различении ИВР связана с существованием биполярных нейронов в медиальной верхней оливе (МВО, англ. medial superior olive, MSO) слуховой части ствола мозга у млекопитающих и ламинарном ядре (nucleus laminaris, NL) у птиц. МВО клетки имеют биполярную морфологию и расположены в единой парасагитальной плоскости с двумя главными дендритами, которые выходят от сомы на 180 градусов друг от друга и расположены ортогонально к дорсовентральной оси ядра.
Нейроны МВО, — в некоторых из которых аксоны даже возникают из дендритов, а не сомы[79], — получают билатеральные возбуждающие входные импульсы от кустистых (bushy) клеток в улитковом ядре (cochlear nucleus) и имеют одну из самых уникальных организаций дендритов во всей ЦНС. Входные сигналы от двух ушей поступают к одной МВО-клетке через два разных дендрита: ипсилатеральные входные сигналы поступают к латеральным дендритам, а контрлатеральные сигналы — к медиальным дендритам. Такая невероятная организация делает возможным разделение входных сигналов, что приводит к их электротонической изоляции, позволяет улучшить суммацию сигналов из двух различных источников и избежать доминирования сигнала из более сильного дендрита[80].
Вследствие быстрых входных синаптических сигналов, эти входные импульсы фазово синхронизированы с формой волны стимула с точностью, большей, чем слуховые нервные волокна. МВО-нейроны также синхронизированы по фазе как с монауральними (от одного уха), так и с бинауральними (от двух ушей) стимулами, но проявляют максимальную реакцию тогда, когда фазово синхронизированные потенциалы действия с двух сторон поступают одновременно, то есть, когда разница в задержках проводимости компенсирует ИВР[81].
Определяющая роль дендритных вычислений в слуховом выявлении совпадений была изначально предусмотрена теоретически, на основе компартментного моделирования и кабельных свойств дендритов, которые расширили традиционные точечные модели нейронов, добавив к ним дендритные сегменты[82]. Эта модель не только объяснила, как дендриты усиливают выявления ИВР, но и также то, почему существует дендритный градиент.
Существование дендритов в МВО (NL) клетках ведёт к появлению двух биофизических механизмов, которые способствуют выявлению совпадений. Первый механизм — это пространственная изоляция входных сигналов, что делает возможным нелинейную интеграцию входных импульсов от двух ушей. Второй: дендритный пучок действует как сток тока для входных синаптических сигналов. Максимальный ответ МВО-нейронов возможен только тогда, когда (после компенсации ИВР с помощью опозданий) входные сигналы, которые они получают от двух ушей, совпадают по времени. Поэтому МВО слабее реагируют на импульсы, поступающие от одного уха, даже если те удвоились. Если бы МВО была линейной системой, такое различение было бы невозможно, поскольку требуется некоторый нелинейный механизм, который демпферует слуховой сигнал, если тот поступает только от одного уха, но не тогда, когда от двух ушей одновременно[82].
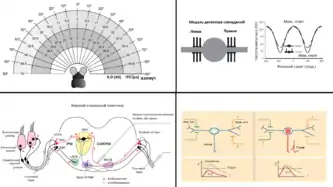
А.(вверху, слева) Интерауральная временная разница (ИВР, ITD) и интерауральная разница интенсивностей (ИРИ) в ответ на тон с частотой 35 кГц у маленькой летучей мыши (Molossus rufus), в зависимости от позиции источника звука в горизонтальной плоскости. Заметны большие ИРИ, вплоть до 40-50 дБ, в сравнении с очень малыми ИВР, до 50 мкс.
B.(внизу, слева) Схема локализации низкочастотных звуков в верхнем оливарном комплексе млекопитающих.
С. (вверху, справа) а) Модель биполярного нейрона для детектора совпадений. б) Модель ИВР кривой для стимула в 500 Гц показывает улучшение в кодировке ИВР, когда к точечной соме были добавлены (пассивные) дендриты. Разница между максимальной (0 градусов опоздания) и минимальной (180 градусов опоздания) спаечной частотой была наибольшей, когда входные сигналы локализовались на дендритах (сплошная кривая), в сравнении с моделью точечного, бездендритного нейрона (пунктирная кривая).[82]
D. (внизу, слева) Схематическое представление влияния биполярных дендритов на локализацию звука. а) Когда входной сигнал поступает только от одного уха, достигая лишь одного дендрита, следствием нелинейной суммации сигнала в дендрите является генерация менее деполяризующего синаптического тока. Как следствие, синаптический потенциал является слишком малым, чтобы сгенерировать потенциал действия в аксоне. b) Когда входные сигналы поступают одновременно от двух ушей и распределяются на разных дендритах, то нелинейность в суммировании синаптических импульсов является менее существенной. Как следствие, синаптический потенциал выше порога возбуждения и может генерировать потенциал действия в аксоне.
Повышение выявления совпадений (а следовательно и локализации звука) МВО-клетками осуществляется благодаря нелинейной суммации (т. н. «насыщение нелинейности») входных возбуждающих синаптических сигналов и увеличению нелинейной потери, когда сигналы скапливаются на одном дендрите, а не распределяются между несколькими. Когда синаптические входные сигналы активируются на том же самом дендрите, то возникает большее локальное изменение напряжения, которое снижает движущую силу для синаптического тока в этом дендрите, чем если бы синапсы формировали контакты на различных дендритах. В последнем случае синапс генерирует более деполяризующий ток и поэтому возникает большая вероятность порождения потенциала действия в аксоне, который в результате сигнализирует, что звук пришел с данного места.
Модель детектора совпадений с дендритами также предусмотрела оптимальную длину дендритов для разных частот звуков. Если частоты увеличиваются, то длина дендритов должна уменьшаться, поскольку при высоких звуковых частотах фазовая синхронизация становится слабее, и в спайках возникают фазовая дрожь (джиттер), которая препятствуют выявлению совпадений и локализации источника звука.
Таким образом МВО-нейроны проявляют микросекундную избирательность к ИВР. Когда оба возбуждающих пути к дендритам активируются в пределах узкого временного интервала, МВО-нейроны обнаруживают совпадение синаптических входных импульсов и генерируют потенциал действия. Когда же эти пути активируются асинхронно, то МВО-нейроны не генерируют ПД. Поэтому функция отклика ИВР представляет вариацию частоты разрядов МВО-клеток с относительным опозданием двух входных сигналов, а следовательно и позицию звука вдоль горизонтальной плоскости[82].
Хотя данная модель была построена для пассивных дендритов, недавние экспериментальные исследования показали, что она также хорошо подходит для объяснения активной роли дендритов для бинаурального обнаружения звуковых совпадений[83][84][85].
В одном из недавних исследований путём одновременной сома-дендритной регистрации МВО-нейронов песчанки in vitro и компартментного моделирования исследователи обнаружили потенциалзависимое обострение ВПСП ближе к соме, которое противодействует дендритному фильтрованию, что снижает временную точность импульсов[84]. Эти исследования впервые продемонстрировали неоднородное распределение низко-пороговых калиевых (KLVA) каналов в соме и дендритах, их влияние на синаптическую временную синхронизацию в слуховой системе.
Хотя Kv1 каналы присутствуют как в соме, так и дендритах, их плотность более смещена в сторону сомы. Когда ВПСП распространяются вдоль отдалённых дендритов МВО-нейронов, пассивная кабельная фильтрация, присущая этим дендритам, задерживает время нарастания ВПСП и увеличивает их продолжительность. Обусловленное Kv1 каналами обострение ВПСП не имеет на эти дендриты ощутимого влияния даже во время сильной деполяризации. Однако когда эти импульсы проникают в более близкие дендриты и сому, они дифференциально активируют значительно большие низко-пороговые калиевые токи (IKLVA) у сомы. Эта активация IKLVA является достаточно длительной в течение бинауральной стимуляции.
Таким образом, соматически смещённое распределение IKLVA компенсирует дендритную кабельную фильтрацию и сохраняет микросекундную временную точность синаптических потенциалов, возникающих в отделенных регионах дендритов, обостряя ВПСП и таким образом улучшая временную точность бинауральных детекторов совпадений. Совсем по-другому, хотя и для той же цели (выявление совпадений), используются дендриты клетками-осьминогами (octopus cells), которые находятся в «заднем кохлеарном вентральном ядре» (PCVN). Они обнаруживают совпадение активации группы слуховых нервных волокон (>60 в мыши) широкополосными кратковременными звуками. Последние порождают бегущие волны в улитке, которые вызывают возбуждение слуховых нервных волокон, настроенных на высокие частоты, на несколько миллисекунд раньше, чем волокон, настроенных на низкие частоты[86].
В отличие от МВО-клеток, клетки-осьминоги для компенсации подобного кохлеарного опоздания бегущей волны используют дендритную пассивную фильтрацию, порождая большие, быстро растущие интегрированные ВПСП у сомы[87].
Если ещё относительно недавно при моделировании и изучении нейронных вычислений в слуховой системе дендриты не принимались во внимание, то с появлением линейной и нелинейной кабельных теорий было предусмотрено, а последующие исследования это экспериментально подтвердили, что без включения дендритов в нейронные модели ряд уникальных черт слуховой системы не могут быть объяснены. Однако ещё больший прогресс в исследовании сложных дендритных вычислений был сделан с открытием их активных свойств, способности модулировать и изменять входные сигналы.
Для случая слуховой системы такое открытие активной роли дендритов было недавно сделано в новаторском исследовании in vivo кальциевой активности у пирамидальных нейронов слоя 2/3 слуховой коры мыши на уровне индивидуальных шипиков[15]. Подобно дендритам зрительной системы, было обнаружено, что сенсорная (звуковая) стимуляция может активировать даже один шипик и что различные шипики на том же самом дендрите отдают предпочтение тонам различных частот и даже способны отвечать на широкополосный шум. Это исследование впервые показало, что сенсорная активность стимулирует не дендритный ствол, а шипик, который затем предстаёт как отдельный частотно-выборочный (frequency selective) сегмент дендритного дерева. Вместе с тем, в отличие от большинства исследований, которые свидетельствуют о кластеризации синаптических входных сигналов, здесь было обнаружено, что шипики слишком неоднородно распределены на дендрите, где даже два соседних шипика отдают предпочтение разным частотам тонов[88].
Впрочем, такая же частотная селективность отдельных сегментов дендритов недавно была предсказана теоретически на основе STDP-пластичности (англ. Spike-timing dependent plasticity, рус. Модель пластичности, зависимая от времени спайка)[89]. Но, в отличие от упомянутого экспериментального исследования, эта теория моделирует дендриты как пассивные кабели, не принимая во внимание потенциалзависимые каналы и кальциевую активность. При таких посылках, теория и эксперимент приходят практически к одинаковым выводам, что синапсы в различных регионах дендритного дерева настроены на разные частоты. За исключением того, что математическая модель предусматривает кластеризацию синапсов, а эксперимент показывает их неоднородное распределение по всему дендритному дереву.
См. также
Источники
- Trachtenberg J.T., Chen B.E., Knott G.W., Feng G., Sanes J.R., Welker E., Svoboda K. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex (англ.) // Nature : journal. — 2002. — Vol. 420, no. 6917. — P. 788—794. — ISSN 0028-0836. — doi:10.1038/nature01273.
- Holtmaat Anthony and Svoboda Karel. Experience-dependent structural synaptic plasticity in the mammalian brain (англ.) // Nature Reviews Neuroscience : journal. — 2009. — Vol. 10, no. 9. — P. 647—658. — ISSN 1471-003X. — doi:10.1038/nrn2699.
- Matsuzaki M., Ellis-Davies G., Nemoto T., Miyashita Y., Iino M., and Kasai H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons (англ.) // Nature Neuroscience : journal. — 2001. — Vol. 4, no. 11. — P. 1086—1092. — ISSN 10976256. — doi:10.1038/nn736.
- Dent Erik W., Merriam Elliott B., and Hu Xindao. The dynamic cytoskeleton: backbone of dendritic spine plasticity (англ.) // Current Opinion in Neurobiology : journal. — Elsevier, 2011. — Vol. 21, no. 1. — P. 175—181. — ISSN 09594388. — doi:10.1016/j.conb.2010.08.013.
- Noguchi J., Nagaoka A., Watanabe S., Ellis-Davies G.C.R., Kitamura K., Kano M., Matsuzaki Masanori, and Kasai H. In vivo two-photon uncaging of glutamate revealing the structure-function relationships of dendritic spines in the neocortex of adult mice (англ.) // The Journal of Physiology : journal. — 2011. — Vol. 589, no. 10. — P. 2447—2457. — ISSN 00223751. — doi:10.1113/jphysiol.2011.207100.
- Hübener Mark and Bonhoeffer Tobias. Searching for Engrams (англ.) // Neuron. — Cell Press, 2010. — Vol. 67, no. 3. — P. 363—371. — ISSN 08966273. — doi:10.1016/j.neuron.2010.06.033.
- Kasai Haruo, Fukuda Masahiro, Watanabe Satoshi, Hayashi-Takag Akiko, and Noguchi Jun. Structural dynamics of dendritic spines in memory and cognition (англ.) // Trends in Neurosciences : journal. — Cell Press, 2010. — Vol. 33, no. 3. — P. 121—129. — ISSN 01662236. — doi:10.1016/j.tins.2010.01.001.
- Xu Tonghui, Yu Xinzhu, Perlik Andrew J., Tobin Willie F., Zweig Jonathan A., Tennant Kelly, Jones Theresa, and Zuo Yi. Rapid formation and selective stabilization of synapses for enduring motor memories (англ.) // Nature : journal. — 2009. — Vol. 462, no. 7275. — P. 915—919. — ISSN 0028-0836. — doi:10.1038/nature08389.
- Kampa B. M., Letzkus J. J., and Stuart G. J. Requirement of dendritic calcium spikes for induction of spike-timing-dependent synaptic plasticity (англ.) // The Journal of Physiology : journal. — 2006. — Vol. 574, no. 1. — P. 283—290. — ISSN 0022-3751. — doi:10.1113/jphysiol.2006.111062.
- Malenka Robert C. Postsynaptic factors control the duration of synaptic enhancement in area CA1 of the hippocampus (англ.) // Neuron : journal. — Cell Press, 1991. — Vol. 6, no. 1. — P. 53—60. — ISSN 08966273. — doi:10.1016/0896-6273(91)90121-F.
- Golding Nace L., Staff Nathan P. and Spruston Nelson. Dendritic spikes as a mechanism for cooperative long-term potentiation (англ.) // Nature : journal. — 2002. — Vol. 418, no. 6895. — P. 326—331. — ISSN 0028-0836. — doi:10.1038/nature00854.
- Sjöström P. and Nelson S. Spike timing, calcium signals and synaptic plasticity (англ.) // Current Opinion in Neurobiology : journal. — Elsevier, 2002. — Vol. 12, no. 3. — P. 305—314. — ISSN 09594388. — doi:10.1016/S0959-4388(02)00325-2.
- Svoboda Karel, Mainen Zachary F. and Malinow Roberto. Synaptic calcium transients in single spines indicate that NMDA receptors are not saturated (англ.) // Nature : journal. — 1999. — Vol. 399, no. 6732. — P. 151—155. — ISSN 00280836. — doi:10.1038/20187.
- Jia Hongbo, Rochefort Nathalie L., Chen Xiaowei, and Konnerth Arthur. Dendritic organization of sensory input to cortical neurons in vivo (англ.) // Nature : journal. — 2010. — Vol. 464, no. 7293. — P. 1307—1312. — ISSN 0028-0836. — doi:10.1038/nature08947.
- Chen Xiaowei, Leischner Ulrich, Rochefort Nathalie L., Nelken Israel, and Konnerth Arthur. Functional mapping of single spines in cortical neurons in vivo (англ.) // Nature : journal. — 2011. — Vol. 475, no. 7357. — P. 501—505. — ISSN 0028-0836. — doi:10.1038/nature10193.
- Feldman Daniel E. The Spike-Timing Dependence of Plasticity (англ.) // Neuron. — Cell Press, 2012. — Vol. 75, no. 4. — P. 556—571. — ISSN 08966273. — doi:10.1016/j.neuron.2012.08.001.
- Bi Guo-qiang Bi and Poo Mu-ming. Synaptic modifications in cultured hippocampal neurons: dependence on spike timing, synaptic strength, and postsynaptic cell type (англ.) // The Journal of Neuroscience : journal. — 1998. — 15 December (vol. 24). — P. 10464—10472.
- Gerstner Wulfram and Kistler Werner M. Spiking Neuron Models: Single Neurons, Populations, Plasticity (англ.). — Cambridge University Press, 2002. — P. 480. — ISBN 9780521890793.
- Magee J. C. and Johnston D. A Synaptically Controlled, Associative Signal for Hebbian Plasticity in Hippocampal Neurons (англ.) // Science : journal. — 1997. — Vol. 275, no. 5297. — P. 209—213. — ISSN 00368075. — doi:10.1126/science.275.5297.209.
- Johnston D., Christie B. R., Frick A., Gray R., Hoffman D. A., Schexnayder L. K., Watanabe S., and Yuan L.-L. Active dendrites, potassium channels and synaptic plasticity (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences : journal. — 2003. — Vol. 358, no. 1432. — P. 667—674. — ISSN 0962-8436. — doi:10.1098/rstb.2002.1248.
- Nevian T. and Sakmann B. Single Spine Ca2+ Signals Evoked by Coincident EPSPs and Backpropagating Action Potentials in Spiny Stellate Cells of Layer 4 in the Juvenile Rat Somatosensory Barrel Cortex (англ.) // Journal of Neuroscience : journal. — 2004. — Vol. 24, no. 7. — P. 1689—1699. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.3332-03.2004.
- Abbott L. F., Song Sen and Miller Kenneth D. Competitive Hebbian learning through spike-timing-dependent synaptic plasticity (англ.) // Nature Neuroscience : journal. — 2000. — Vol. 3, no. 9. — P. 919—926. — ISSN 10976256. — doi:10.1038/78829.
- Stuart Greg J. and Häusser Michael. Dendritic coincidence detection of EPSPs and action potentials (англ.) // Nature Neuroscience : journal. — 2001. — Vol. 4, no. 1. — P. 63—71. — doi:10.1038/82910.
- Kairiss Edward W., Mainen Zachary F., Claiborne Brenda J. and Brown Thomas H. Dendritic Control of Hebbian Computations (неопр.). — 1992. — С. 69—83. — doi:10.1007/978-1-4615-4010-6_7.
- Hunt David L and Castillo Pablo E. Synaptic plasticity of NMDA receptors: mechanisms and functional implications (англ.) // Current Opinion in Neurobiology : journal. — Elsevier, 2012. — Vol. 22, no. 3. — P. 496—508. — ISSN 09594388. — doi:10.1016/j.conb.2012.01.007.
- Rikhye Rajeev V., Tan Xulin , and Dongen Antonius MJ Van. Local activity in dendrites controls STDP by altering NMDA receptor kinetics (англ.) // BMC Neuroscience : journal. — 2013. — Vol. 14. — P. 399.
- Lisman John and Spruston Nelson. Postsynaptic depolarization requirements for LTP and LTD: a critique of spike timing-dependent plasticity (англ.) // Nature Neuroscience : journal. — 2005. — Vol. 8, no. 7. — P. 839—841. — ISSN 1097-6256. — doi:10.1038/nn0705-839.
- Mel, BW. The clusteron: toward a simple abstraction for a complex neuron. // Advances in neural information processing systems (англ.) / Moody J. , Hanson S. , & R. Lippmann. — Morgan Kaufmann Publishers, 1992. — P. 35—42.. — ISBN 9781558602229.
- Poirazi Panayiota and Mel Bartlett W. Impact of Active Dendrites and Structural Plasticity on the Memory Capacity of Neural Tissue (англ.) // Neuron : journal. — Cell Press, 2001. — Vol. 29, no. 3. — P. 779—796. — ISSN 08966273. — doi:10.1016/S0896-6273(01)00252-5.
- Häusser Michael and Mel Bartlett. Dendrites: bug or feature? (англ.) // Current Opinion in Neurobiology. — Elsevier, 2003. — Vol. 13, no. 3. — P. 372—383. — ISSN 09594388. — doi:10.1016/S0959-4388(03)00075-8.
- Poirazi Panayiota, Brannon Terrence and Mel Bartlett W. Pyramidal Neuron as Two-Layer Neural Network (англ.) // Neuron. — Cell Press, 2003. — Vol. 37, no. 6. — P. 989—999. — ISSN 08966273. — doi:10.1016/S0896-6273(03)00149-1.
- Frick Andreas, Magee Jeffrey, & Johnston Daniel. LTP is accompanied by an enhanced local excitability of pyramidal neuron dendrites (англ.) // Nature Neuroscience : journal. — 2004. — Vol. 7, no. 2. — P. 126—135. — ISSN 1097-6256. — doi:10.1038/nn1178.
- Losonczy Attila, Makara Judit K., and Magee Jeffrey C. Compartmentalized dendritic plasticity and input feature storage in neurons (англ.) // Nature : journal. — 2008. — Vol. 452, no. 7186. — P. 436—441. — ISSN 0028-0836. — doi:10.1038/nature06725.
- Koch Christof and Segev Idan. The role of single neurons in information processing (англ.) // Nature Neuroscience : journal. — 2000. — Vol. 3, no. Supp. — P. 1171—1177. — ISSN 10976256. — doi:10.1038/81444.
- Branco Tiago and Häusser Michael. The single dendritic branch as a fundamental functional unit in the nervous system (англ.) // Current Opinion in Neurobiology : journal. — Elsevier, 2010. — Vol. 20, no. 4. — P. 494—502. — ISSN 09594388. — doi:10.1016/j.conb.2010.07.009.
- Losonczy Attila and Magee Jeffrey C. Integrative Properties of Radial Oblique Dendrites in Hippocampal CA1 Pyramidal Neurons (англ.) // Neuron : journal. — Cell Press, 2006. — Vol. 50, no. 2. — P. 291—307. — ISSN 08966273. — doi:10.1016/j.neuron.2006.03.016.
- Makara Judit K., Losonczy Attila, Wen Quan, and Magee Jeffrey C. Experience-dependent compartmentalized dendritic plasticity in rat hippocampal CA1 pyramidal neurons (англ.) // Nature Neuroscience : journal. — 2009. — Vol. 12, no. 12. — P. 1485—1487. — ISSN 1097-6256. — doi:10.1038/nn.2428.
- Legenstein R. and Maass W. Branch-Specific Plasticity Enables Self-Organization of Nonlinear Computation in Single Neurons (англ.) // Journal of Neuroscience : journal. — 2011. — Vol. 31, no. 30. — P. 10787—10802. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.5684-10.2011.
- Lebedeva S.A., Stepanyuk A. R., and Belan P. V. Local Signalization in Dendrites and Mechanisms of Short-Term Memory (англ.) // Neurophysiology : journal. — 2013. — Vol. 45, no. 4. — P. 359—367. — ISSN 0090-2977. — doi:10.1007/s11062-013-9381-6.
- Kleindienst T., Winnubst J., Roth-Alpermann C., Bonhoeffer T., Lohmann C. Activity-Dependent Clustering of Functional Synaptic Inputs on Developing Hippocampal Dendrites (англ.) // Neuron : journal. — Cell Press, 2011. — Vol. 72, no. 6. — P. 1012—1024. — ISSN 08966273. — doi:10.1016/j.neuron.2011.10.015.
- Makino Hiroshi and Malinow Roberto. Compartmentalized versus Global Synaptic Plasticity on Dendrites Controlled by Experience (англ.) // Neuron : journal. — Cell Press, 2011. — Vol. 72, no. 6. — P. 1001—1011. — ISSN 08966273. — doi:10.1016/j.neuron.2011.09.036.
- Yadav A., Gao Yuan Z., Rodriguez A., Dickstein D.L., Wearne S.L., Luebke J.I., Hof P.R., Weaver C.M. Morphologic evidence for spatially clustered spines in apical dendrites of monkey neocortical pyramidal cells (англ.) // The Journal of Comparative Neurology : journal. — 2012. — Vol. 520, no. 13. — P. 2888—2902. — ISSN 00219967. — doi:10.1002/cne.23070.
- Fu Min, Yu Xinzhu, Lu Ju and Zuo Yi. Repetitive motor learning induces coordinated formation of clustered dendritic spines in vivo (англ.) // Nature : journal. — 2012. — Vol. 483, no. 7387. — P. 92—95. — ISSN 0028-0836. — doi:10.1038/nature10844.
- Chen T.W., Wardill T.J., Sun Y., Pulver S.R., Renninger S.L., Baohan A., Schreiter E.R., Kerr R.A., Orger M.B., Jayaraman V., Looger L.L., Svoboda K., and Kim D.S. Ultrasensitive fluorescent proteins for imaging neuronal activity (англ.) // Nature : journal. — 2013. — Vol. 499, no. 7458. — P. 295—300. — ISSN 0028-0836. — doi:10.1038/nature12354.
- Takahashi N., Kitamura K., Matsuo N., Mayford M., Kano M., Matsuki N., Ikegaya Y. Locally Synchronized Synaptic Inputs (англ.) // Science. — 2012. — Vol. 335, no. 6066. — P. 353—356. — ISSN 0036-8075. — doi:10.1126/science.1210362.
- Varga Z., Jia H., Sakmann B., and Konnerth A. Dendritic coding of multiple sensory inputs in single cortical neurons in vivo (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2011. — Vol. 108, no. 37. — P. 15420—15425. — ISSN 0027-8424. — doi:10.1073/pnas.1112355108.
- Larkum Matthew E and Nevian Thomas. Synaptic clustering by dendritic signalling mechanisms (англ.) // Current Opinion in Neurobiology : journal. — Elsevier, 2008. — Vol. 18, no. 3. — P. 321—331. — ISSN 09594388. — doi:10.1016/j.conb.2008.08.013.
- DeRoo Mathias, Klauser Paul and Muller Dominique. LTP Promotes a Selective Long-Term Stabilization and Clustering of Dendritic Spines (англ.) // PLoS Biology : journal. — 2008. — Vol. 6, no. 9. — P. e219. — ISSN 1544-9173. — doi:10.1371/journal.pbio.0060219.
- Goldberg Jesse, Holthoff Knut, adn Yuste Rafael. A problem with Hebb and local spikes (англ.) // Trends in Neurosciences : journal. — Cell Press, 2002. — Vol. 25, no. 9. — P. 433—435. — ISSN 01662236. — doi:10.1016/S0166-2236(02)02200-2.
- Holthoff K., Kovalchuk Y., Yuste R., and Konnerth A. Single-shock LTD by local dendritic spikes in pyramidal neurons of mouse visual cortex (англ.) // The Journal of Physiology : journal. — 2004. — Vol. 560, no. 1. — P. 27—36. — ISSN 0022-3751. — doi:10.1113/jphysiol.2004.072678.
- Sjostrom P. J., Rancz E. A., Roth A., Hausser M. Dendritic Excitability and Synaptic Plasticity (англ.) // Physiological Reviews : journal. — 2008. — Vol. 88, no. 2. — P. 769—840. — ISSN 0031-9333. — doi:10.1152/physrev.00016.2007.
- Kamondi Anita, Acsády László, and Buzsáki György. {{{заглавие}}} (неопр.) // The Journal of Neuroscience,. — 1998. — 15 May (т. 18). — С. 3919—3928.
- Abraham Wickliffe C. and Bear Mark F. Metaplasticity: the plasticity of synaptic plasticity (англ.) // Trends in Neurosciences : journal. — Cell Press, 1996. — Vol. 19, no. 4. — P. 126—130. — ISSN 01662236. — doi:10.1016/S0166-2236(96)80018-X.
- Noguchi Jun, Matsuzaki Masanori, Ellis-Davies Graham C.R., and Kasai Haruo. Spine-Neck Geometry Determines NMDA Receptor-Dependent Ca2+ Signaling in Dendrites (нем.) // Neuron : magazin. — Cell Press, 2005. — Bd. 46, Nr. 4. — S. 609—622. — ISSN 08966273. — doi:10.1016/j.neuron.2005.03.015.
- Rusakov D.A. and Fine A. Extracellular Ca2+ Depletion Contributes to Fast Activity-Dependent Modulation of Synaptic Transmission in the Brain (англ.) // Neuron : journal. — Cell Press, 2003. — Vol. 37, no. 2. — P. 287—297. — ISSN 08966273. — doi:10.1016/S0896-6273(03)00025-4.
- Holthoff Knut, Kovalchuk Y., and Konnerth A. Dendritic spikes and activity-dependent synaptic plasticity (англ.) // Cell and Tissue Research : journal. — 2006. — Vol. 326, no. 2. — P. 369—377. — ISSN 0302-766X. — doi:10.1007/s00441-006-0263-8.
- Chavis Pascale, Sun Le, Han Xu, and He Shigang. Direction-Selective Circuitry in Rat Retina Develops Independently of GABAergic, Cholinergic and Action Potential Activity (англ.) // PLoS ONE : journal. — 2011. — Vol. 6, no. 5. — P. e19477. — ISSN 1932-6203. — doi:10.1371/journal.pone.0019477.
- Vaney David I., Sivyer Benjamin and Taylor W. Rowland. Direction selectivity in the retina: symmetry and asymmetry in structure and function (англ.) // Nature Reviews Neuroscience : journal. — 2012. — ISSN 1471-003X. — doi:10.1038/nrn3165.
- Single S. and Borst A. Dendritic Integration and Its Role in Computing Image Velocity (англ.) // Science : journal. — 1998. — Vol. 281, no. 5384. — P. 1848—1850. — doi:10.1126/science.281.5384.1848.
- Barlow HB and Levick W. R. The mechanism of directionally selective units in rabbit's retina (англ.) // The Journal of physiology : journal. — 1965. — 1 June (vol. 178,). — P. 477—504. (недоступная ссылка)
- Kong Jee-Hyun, Fish Daniel R., Rockhill Rebecca L., and Masland Richard H. Diversity of ganglion cells in the mouse retina: Unsupervised morphological classification and its limits (англ.) // The Journal of Comparative Neurology : journal. — 2005. — Vol. 489, no. 3. — P. 293—310. — ISSN 00219967. — doi:10.1002/cne.20631.
- Masland R. Neuronal diversity in the retina (англ.) // Current Opinion in Neurobiology. — Elsevier, 2001. — Vol. 11, no. 4. — P. 431—436. — ISSN 09594388. — doi:10.1016/S0959-4388(00)00230-0.
- Rall Wilfrid. Time constants and electrotonic length of membrane cylinders and neurons (англ.) // Biophysical Journal : journal. — 1969. — December (vol. 9). — P. 1483—1508. — doi:10.1016/S0006-3495(69)86467-2.
- Koch C., Poggio T. and Torres V. Retinal Ganglion Cells: A Functional Interpretation of Dendritic Morphology (англ.) // Philosophical Transactions of the Royal Society B: Biological Sciences : journal. — 1982. — Vol. 298, no. 1090. — P. 227—263. — ISSN 0962-8436. — doi:10.1098/rstb.1982.0084.
- Taylor Rowland W. and Vaney David I. Diverse Synaptic Mechanisms Generate Direction Selectivity in the Rabbit Retina (англ.) // The Journal of Neuroscience : journal. — 2002. — 1 September (vol. 22). — P. 7712—7720.
- Borg-Graham L and Grzywacz NM. A model of the directional selectivity circuit in retina: transformations by neurons singly and in concert // Single Neuron Computation (неопр.) / McKenna T., Davis J. L., Zoenetzer S. F.. — Academic Press, 1992. — С. 347—375. — ISBN 0-12-484815-X.
- Masland Richard H. The fundamental plan of the retina (англ.) // Nature Neuroscience : journal. — 2001. — Vol. 4, no. 9. — P. 877—886. — ISSN 1097-6256. — doi:10.1038/nn0901-877.
- Macneil Margaret A., Heussy John K., Dacheux Ramon F., Raviola Elio, and Masland Richard H. The shapes and numbers of amacrine cells: Matching of photofilled with Golgi-stained cells in the rabbit retina and comparison with other mammalian species (англ.) // The Journal of Comparative Neurology : journal. — 1999. — Vol. 413, no. 2. — P. 305—326. — ISSN 0021-9967. — doi:10.1002/(SICI)1096-9861(19991018)413:2<305::AID-CNE10>3.0.CO;2-E.
- Yoshida Kazumichi, Watanabe Dai, Ishikane Hiroshi, Tachibana Masao, Pastan Ira, and Nakanishi Shigetada. A Key Role of Starburst Amacrine Cells in Originating Retinal Directional Selectivity and Optokinetic Eye Movement (англ.) // Neuron : journal. — Cell Press, 2001. — Vol. 30, no. 3. — P. 771—780. — ISSN 08966273. — doi:10.1016/S0896-6273(01)00316-6.
- Euler Thomas, Detwiler Peter B., and Denk Winfried. Directionally selective calcium signals in dendrites of starburst amacrine cells (англ.) // Nature : journal. — 2002. — Vol. 418, no. 6900. — P. 845—852. — ISSN 0028-0836. — doi:10.1038/nature00931.
- Hausselt Susanne E., Euler Thomas, Detwiler Peter B., and Denk Winfried. A Dendrite-Autonomous Mechanism for Direction Selectivity in Retinal Starburst Amacrine Cells (англ.) // PLoS Biology : journal. — 2007. — Vol. 5, no. 7. — P. e185. — ISSN 1544-9173. — doi:10.1371/journal.pbio.0050185.
- Taylor W.Rowland and Vaney David I. New directions in retinal research (англ.) // Trends in Neurosciences : journal. — Cell Press, 2003. — Vol. 26, no. 7. — P. 379—385. — ISSN 01662236. — doi:10.1016/S0166-2236(03)00167-X.
- Grimes William N., Zhang Jun, Graydon Cole W., Kachar Bechara, and Diamond Jeffrey S. Retinal Parallel Processors: More than 100 Independent Microcircuits Operate within a Single Interneuron (англ.) // Neuron : journal. — Cell Press, 2010. — Vol. 65, no. 6. — P. 873—885. — ISSN 08966273. — doi:10.1016/j.neuron.2010.02.028.
- Gollisch Tim and Meister Markus. Eye Smarter than Scientists Believed: Neural Computations in Circuits of the Retina (англ.) // Neuron : journal. — Cell Press, 2010. — Vol. 65, no. 2. — P. 150—164. — ISSN 08966273. — doi:10.1016/j.neuron.2009.12.009.
- Chávez Andrés E., Singer Joshua H., and Diamond Jeffrey S. Fast neurotransmitter release triggered by Ca influx through AMPA-type glutamate receptors (англ.) // Nature : journal. — 2006. — Vol. 443, no. 7112. — P. 705—708. — ISSN 0028-0836. — doi:10.1038/nature05123.
- Chklovskii D. Synaptic Connectivity and Neuronal Morphology (англ.) // Neuron. — Cell Press, 2004. — 2 September (vol. 43). — P. 609—617. — doi:10.1016/j.neuron.2004.08.012.
- Oertel D. THE ROLE OF TIMING IN THE BRAIN STEM AUDITORY NUCLEI OF VERTEBRATES (англ.) // Annual Review of Physiology. — 1999. — Vol. 61, no. 1. — P. 497—519. — ISSN 0066-4278. — doi:10.1146/annurev.physiol.61.1.497.
- Grothe B., Pecka M., & McAlpine D. Mechanisms of Sound Localization in Mammals (англ.) // Physiological Reviews : journal. — 2010. — Vol. 90, no. 3. — P. 983—1012. — ISSN 0031-9333. — doi:10.1152/physrev.00026.2009.
- Zhou Y., Carney Laurel H., and Colburn H. Steven. A Model for Interaural Time Difference Sensitivity in the Medial Superior Olive: Interaction of Excitatory and Inhibitory Synaptic Inputs, Channel Dynamics, and Cellular Morphology (англ.) // Journal of Neuroscience : journal. — 2005. — Vol. 25, no. 12. — P. 3046—3058. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.3064-04.2005.
- Stiefel Klaus M., Tapson Jonathan, and van Schaik André. Temporal Order Detection and Coding in Nervous Systems (англ.) // Neural Computation : journal. — 2013. — Vol. 25, no. 2. — P. 510—531. — ISSN 0899-7667. — doi:10.1162/NECO_a_00400.
- Joris Philip X., Smith Philip H., and Yin Tom C.T. Coincidence Detection in the Auditory System (англ.) // Neuron. — Cell Press, 1998. — Vol. 21, no. 6. — P. 1235—1238. — ISSN 08966273. — doi:10.1016/S0896-6273(00)80643-1.
- Agmon-Snir Hagai, Carr Catherine E. and Rinzel John. The role of dendrites in auditory coincidence detection (англ.) // Nature : journal. — 1998. — Vol. 393, no. 6682. — P. 268—272. — ISSN 00280836. — doi:10.1038/30505.
- Trussell Laurence O. Synaptic mechanisms for coding timing in auditory neurons (англ.) // Annual Review of Physiology : journal. — 1999. — Vol. 61, no. 1. — P. 477—496. — ISSN 0066-4278. — doi:10.1146/annurev.physiol.61.1.477.
- Mathews Paul J., Jercog Pablo E., Rinzel John, Scott Luisa L., and Golding Nace L. Control of submillisecond synaptic timing in binaural coincidence detectors by Kv1 channels (англ.) // Nature Neuroscience : journal. — 2010. — Vol. 13, no. 5. — P. 601—609. — ISSN 1097-6256. — doi:10.1038/nn.2530.
- Scott L.L., Mathews P.J., and Golding N.L. Perisomatic Voltage-Gated Sodium Channels Actively Maintain Linear Synaptic Integration in Principal Neurons of the Medial Superior Olive (англ.) // Journal of Neuroscience : journal. — 2010. — Vol. 30, no. 6. — P. 2039—2050. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.2385-09.2010.
- Golding N.L. and Oertel D. Synaptic integration in dendrites: exceptional need for speed (англ.) // The Journal of Physiology : journal. — 2012. — Vol. 590, no. 22. — P. 5563—5569. — ISSN 0022-3751. — doi:10.1113/jphysiol.2012.229328.
- McGinley M. J., Liberman M. C., Bal R., and Oertel D. Generating Synchrony from the Asynchronous: Compensation for Cochlear Traveling Wave Delays by the Dendrites of Individual Brainstem Neurons (англ.) // Journal of Neuroscience : journal. — 2012. — Vol. 32, no. 27. — P. 9301—9311. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.0272-12.2012.
- Grienberger Christine, Adelsberger Helmuth, Stroh Albrecht, Milos Ruxandra-Iulia, Garaschuk Olga, Schierloh Anja, Nelken Israel, and Konnerth Arthur. Sound-evoked network calcium transients in mouse auditory cortex in vivo (англ.) // The Journal of Physiology : journal. — 2011. — P. no—no. — ISSN 00223751. — doi:10.1113/jphysiol.2011.222513.
- Gilson Matthieu, Bürck Moritz, Burkitt Anthony N., and van Hemmen J. Leo. Frequency Selectivity Emerging from Spike-Timing-Dependent Plasticity (англ.) // Neural Computation : journal. — 2012. — Vol. 24, no. 9. — P. 2251—2279. — ISSN 0899-7667. — doi:10.1162/NECO_a_00331.
Литература
- Dendrites (англ.) / Stuart G., Spruston N., Hausser M.. — Oxford University Press, 1999. — P. 139—160. — ISBN ISBN 0198566565..
- The Theoretical Foundation of Dendritic Function: Selected Papers of Wilfrid Rall with Commentaries (англ.) / Segev I., Rinzel J. and Shepherd G.. — MIT Press, 1995. — P. 456. — ISBN 9780262193566.
- Yuste, Rafael. Dendritic Spines (неопр.). — MIT Press, 2010. — С. 264. — ISBN 9780262013505.
- Tuckwell, Henry C. Introduction to Theoretical Neurobiology: Volume 1, Linear Cable Theory and Dendritic Structure (англ.). — Cambridge University Press, 1988. — P. 304. — ISBN 9780521350969.
- Koch, Christof. Biophysics of Computation: Information Processing in Single Neurons (англ.). — Oxford University Press, 2004. — P. 562. — ISBN 9780195181999.
Обзорные статьи
- Magee Jeffrey C and Johnston Daniel. Plasticity of dendritic function (англ.) // Current Opinion in Neurobiology. — Elsevier, 2005. — Vol. 15, no. 3. — P. 334—342. — ISSN 09594388. — doi:10.1016/j.conb.2005.05.013.
- Sjostrom P. J., Rancz E. A., Roth A., Hausser M. Dendritic Excitability and Synaptic Plasticity (англ.) // Physiological Reviews : journal. — 2008. — Vol. 88, no. 2. — P. 769—840. — ISSN 0031-9333. — doi:10.1152/physrev.00016.2007.
- Hausser M. Diversity and Dynamics of Dendritic Signaling (англ.) // Science. — 2000. — 27 October (vol. 290). — P. 739—744. — doi:10.1126/science.290.5492.739.
- London Michael and Häusser Michael. Dendritic computation (англ.) // Annual Review of Neuroscience. — 2005. — Vol. 28, no. 1. — P. 503—532. — ISSN 0147-006X. — doi:10.1146/annurev.neuro.28.061604.135703.
- Mel Bartlett W. Information Processing in Dendritic Trees (англ.) // Neural Computation. — 1994. — Vol. 6, no. 6. — P. 1031—1085. — ISSN 0899-7667. — doi:10.1162/neco.1994.6.6.1031.
- Kasai Haruo, Fukuda Masahiro, Watanabe Satoshi, Hayashi-Takag Akiko, and Noguchi Jun. Structural dynamics of dendritic spines in memory and cognition (англ.) // Trends in Neurosciences : journal. — Cell Press, 2010. — Vol. 33, no. 3. — P. 121—129. — ISSN 01662236. — doi:10.1016/j.tins.2010.01.001.