Метод локальной фиксации потенциала
Метод локальной фиксации потенциала, patch-clamp (англ. patch — фрагмент, clamp здесь — фиксация) — электрофизиологическая методика для изучения свойств ионных каналов, состоящая в том, что фрагмент клеточной мембраны изолируется с помощью специальной микропипетки. Эта методика даёт возможность экспериментатору контролировать разность потенциалов между сторонами мембраны, а также помещать её в среду с определённым химическим составом. В этих хорошо контролируемых условиях измеряют ионные токи, проходящие через мембрану, что, в конечном итоге, позволяет делать выводы о том, как ионные каналы реагируют на электрическое и химическое воздействие. Метод настолько чувствителен, что позволяет наблюдать поведение и химические превращения единичных молекул, взаимодействующих с мембраной.[1] Разработаны экспериментальные протоколы, позволяющие измерять характеристики ионных каналов оптимальным образом. Немецкие исследователи Эрвин Неер и Берт Сакман, разработавшие эту методику, получили Нобелевскую премию в 1991 году.
Задача: исследование трансмембранных ионных токов
Живые клетки покрыты мембраной, структурную основу которой составляет двойной слой липидов, слабо проницаемый для воды и практически непроницаемый для ионов. Каждая клетка должна обмениваться с внешней средой различными веществами и, в частности, ионами. Перенос ионов через мембрану играет важную роль в процессах возбуждения клетки и передачи сигналов. Ионы проникают в клетку и выходят из неё через встроенные в мембрану белковые структуры — каналы и транспортёры[2]
Транспортёры — это мембранные белки, которые соединяются с переносимым веществом по одну сторону мембраны, переносят это вещество через мембрану и затем его освобождают. Такой перенос становится возможным потому, что в результате соединения с веществом транспортёр меняет конформацию (то есть форму, ориентацию). Важнейший транспортёр в клетках эукариот — это натрий-калиевый насос. Для работы этого насоса требуется энергия, которую он черпает из запасённой в клетке АТФ. За один цикл своей работы насос выводит из клетки 3 иона Na+ и вводит в неё 2 иона K+. Одна молекула этого транспортёра совершает примерно 10³ циклов в секунду. Сходная частота циклов характерна и для других видов транспортёров[3].
Каналы — это белки, которые выполняют функцию мембранных пор, так как формируют отверстия, сквозь которые могут проходить ионы. Мембранные каналы селективны — проницаемы только для определённых веществ. Селективность обусловлена радиусом пор и распределением заряженных функциональных групп в них. Существуют каналы, селективно пропускающие ионы натрия (натриевые каналы), а также калиевые, кальциевые и хлорные каналы. Для каждого вида ионов существует не один, а довольно много видов каналов[4]. Сквозь один канал за секунду проходит 106 — 107 ионов[3].
Несмотря на фундаментальные различия в механизме транспорта через каналы и транспортёры, они могут быть образованы высокогомологичными белками. Так, недавно получены данные, что мутация единственной аминокислоты в белке транспортёра двухвалентных металлов DMT1 приводит к его превращению в кальциевый канал. Кроме того, существует по крайней мере один транспортёр (хлорид-бикарбонатный обменник эритроцитов), осуществляющий 105 переносов в секунду[3], что очень близко к скоростям, характерным для каналов, и заставляет предположить существование у него некоего «промежуточного» между каналами и транспортёрами механизма.
Так как ионы — это электрически заряженные молекулы, при их переходе через мембранные каналы переносится и заряд, а значит, через мембрану течёт электрический ток. Этот ток можно измерить. Чем больше разность потенциалов между сторонами мембраны, тем больше ток. Проводимость (отношение тока к разности потенциалов) одиночного канала в открытом состоянии варьирует в зависимости от вида канала, от 4-5 (у CFTR[5] и ENaC[6]) до 30—50 (например, у никотин-чувствительного холинорецептора[7]) пикосименсов. Это значит, что при разности потенциалов, равной 100 мВ, через канал потечёт ток в несколько пикоампер.
Методики изучения электрических свойств клеточной мембраны
Один внутриклеточный электрод. Измерение разности потенциалов
Электрические явления на клеточных мембранах могут измеряться с помощью острых стеклянных микроэлектродов, которые вводятся в клетку. Техника с одним внутриклеточным электродом позволяет измерять разность потенциалов при фиксированном токе. Реже при помощи этой техники можно измерить ток при фиксированном потенциале при помощи методики коммутационной фиксации потенциала. Все измерения происходят исключительно на целой клетке, таким образом изучаются электрические свойства всей клеточной мембраны.
Двухэлектродная фиксация потенциала
Тот факт, что фиксация трансмембранного потенциала позволит измерять мембранную проводимость по изменениям тока при постоянном напряжении, был впервые осознан ещё в 40-х гг. XX века, и вскоре английские исследователи Алан Ходжкин и Эндрю Хаксли (Alan Hodgkin and Andrew Huxley) начали эксперименты с двухэлектродной фиксацией потенциала (2-electrode voltage clamp).[8] Суть метода состоит в следующем. В клетку вводятся два электрода, ещё один — электрод сравнения — остаётся вне клетки. Первый внутриклеточный электрод служит для измерения трансмембранной разности потенциалов (то есть разности потенциалов между ним и электродом сравнения), второй может подавать ток. Использование одного и того же острого электрода для подачи тока и измерения разности потенциалов невозможно. Дело в том, что такой электрод имеет входное электрическое сопротивление порядка 100-200 МОм, что сравнимо с сопротивлением клеточной мембраны, поэтому, если через систему "электрод-клетка" течет ток, то падение напряжения на электроде окажется соизмеримо с падением напряжения на самой мембране, при чем не существует способа выделить эти две части[8]. Далее, специальное устройство — генератор сигнала — задаёт командный потенциал, которому должен быть равен трансмембранный потенциал. Измеренный трансмембранный потенциал подаётся на вход устройства сравнения, которое вычитает измеренный потенциал из командного и, в зависимости от величины разности, подаёт ток на токовый электрод так, чтобы скомпенсировать эту разницу. Монитор тока, в свою очередь, постоянно измеряет величину тока, которая для этого необходима. В 1940-х 50-х годах, когда работали Ходжкин и Хаксли, не существовало микроэлектродов, поэтому в качестве внутриклеточных электродов использовались тонкие проволоки. Это определило выбор объекта — гигантский аксон кальмара диаметром 1 мм, внутрь которого и вводились эти проволоки. На этом объекте методом двухэлектродной фиксации потенциала исследователи выполнили эксперименты, в которых была установлена ионная природа потенциала действия и впервые постулировано существование ионных каналов (Нобелевская премия 1963 г., поделена с Дж. Экклзом, получившим её за исследования в области синаптической передачи). Двухэлектродная фиксация потенциала применяется и в настоящее время, с использованием острых стеклянных микроэлектродов, однако даже с ними эта методика имеет существенные ограничения: два электрода могут быть введены только в весьма крупную клетку (например, ооцит лягушки).
Решение: patch-clamp, его варианты и конфигурации
Гигаомный контакт

В конце семидесятых годов XX в. Эрвин Неер (E.Neher) и Берт Сакман (B.Sakmann) обнаружили, что конусообразные стеклянные пипетки с диаметром кончика 1—2 микрона могут образовывать контакты с клеточной мембраной (границе «мембрана — стекло») с сопротивлением в несколько гигаом — это так называемый гигаомный контакт. Он позволяет изолировать от внешней среды и от остальной части мембраны тот её фрагмент, который находится внутри пипетки. Отграниченный пипеткой фрагмент мембраны и называется patch — «фрагмент», слово clamp (фиксация) в названии метода можно интерпретировать и как захват и изоляцию этого фрагмента, так и как фиксацию трансмембранного потенциала в изолированном фрагменте, или, как будет описано позже, потенциала или тока на целой клетке.[9] В пипетку, заполненную раствором электролита, помещается хлор-серебряный электрод, второй электрод размещается внеклеточно, в омывающей жидкости. Отличие электрической схемы от ранее описанной для двухэлектродной фиксации заключается в том, что один и тот же электрод используется как для измерения разности потенциалов, так и для подачи тока, что оказывается возможным благодаря сравнительно низкому сопротивлению пипетки.
На фото 1 показана часть такой установки.

Огромная сфера, часть которой видна на мониторе в центре фотографии — это клетка (ооцит лягушки Xenopus laevis), находящаяся в данный момент на предметном столике микроскопа, к ней подведена patch-пипетка, её диаметр у носика — около 3 микрон. После установления гигаомного контакта она изолирует фрагмент мембраны площадью приблизительно 7 мкм², ток от ионных каналов в этом фрагменте можно будет записывать.
Конфигурация «Cell-attached»
Только что описанная конфигурация называется cell-attached patch-clamp (patch-clamp «с прикреплённой клеткой»). Она иллюстрируется схемой 2.
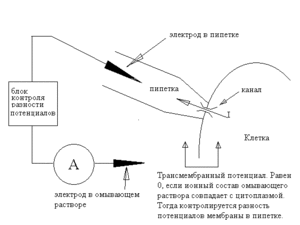
Эта конфигурация, однако, имеет два неудобства.
Во-первых, она не позволяет с достаточной надёжностью измерять, а, следовательно, и задавать трансмембранную разность потенциалов — поскольку оба электрода — как пипеточный, так и внешний, находятся по одну сторону мембраны. Вообще говоря, можно, используя омывающий раствор с ионной композицией, повторяющей состав цитоплазмы, деполяризовать мембрану вне пипетки, так что разность потенциалов между внешним электродом и цитоплазмой исчезнет, а тогда разность потенциалов между электродами окажется равна трансмембранному потенциалу — но все это весьма приблизительно, так как точный состав цитоплазмы нам неизвестен.
Во-вторых, эта конфигурация не позволяет контролировать состав среды вне пипетки — там остаётся цитоплазма, состав которой не вполне определён.
Впрочем, в определенной мере эта особенность конфигурации cell-attached является и достоинством: данная конфигурация позволяет работать в условиях, наиболее близких к физиологическим.
Конфигурация Inside-out
Однако, если пипетку быстрым движением отвести от клетки, то «внутренний» кусочек мембраны оторвется от клетки и получится конфигурация inside-out (наружная сторона внутри), названная так потому, что внутренняя, обычно обращенная к цитоплазме, сторона мембраны окажется снаружи — в омывающем растворе, а наружная — внутри пипетки. (Схема 3.) Альтернативный метод перехода в конфигурацию inside-out заключается в следующем: из конфигурации cell-attached пипетку отводят плавно, формируя с двух сторон закрытую везикулу; затем поднимают пипетку в воздух и опускают во вторую ванночку, с внутриклеточным раствором. При переносе внешняя мембрана везикулы разрушается, в результате формируется конфигурация inside-out.
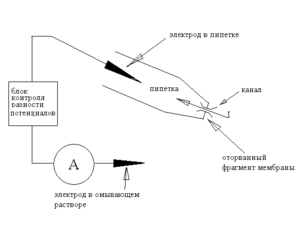
Теперь разность потенциалов на фрагменте мембраны строго равна разности потенциалов между электродами. Очевидно, что при использовании такой модификации пипетку заполняют раствором, имитирующим внеклеточную среду, тогда как омывающий раствор делают близким по составу к цитоплазме. При этом, меняя состав омывающего раствора, можно изучать, как такие изменения в цитоплазме влияют на ток интересующих нас каналов — ведь омывающий раствор контактирует с цитоплазматической стороной мембраны). При этом мы точно знаем как состав жидкости по обе стороны мембраны, так и разности потенциалов, что позволяет достаточно точно характеризовать каналы, используя биофизические уравнения Нернста и Гольдмана-Ходжкина-Каца. При этом, если в изолированный участок мембраны попал один канал, то мы наблюдаем поведение единичной молекулы и, если это лиганд-регулируемый канал-рецептор, то её взаимодействие с другими единичными молекулами.
Конфигурация Whole-cell
Если по условиям эксперимента необходимо менять состав внеклеточной среды, можно использовать конфигурацию whole cell («англ. целая клетка»). В этом случае пипетку не отводят от клетки, а подают в неё отрицательное давление и таким образом разрушают изолированный фрагмент мембраны.
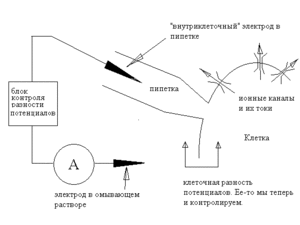
После этого пипетка соединена с внутриклеточной средой; поскольку клетка обычно маленькая, то, благодаря диффузии, состав цитоплазмы вскоре оказывается идентичным составу пипеточного раствора, поэтому мы, как и в предыдущем случае, знаем как состав жидкостей, так и разность потенциалов. Отметим, что если в конфигурации inside-out пипеточный электрод был «внеклеточным», а внутренний — «внутриклеточным», то теперь они поменялись ролями. С точки зрения изучения тока через каналы, преимущество этого метода состоит в том, что здесь оказываются сохранны клеточные структуры и регуляторные механизмы. Правда, низкомолекулярные вещества могут диффундировать из цитоплазмы в пипетку и наоборот, так что полной сохранности регуляторных механизмов в данной конфигурации достичь не удается. Определенным недостатком конфигурации можно считать то, что измеряется суммарный ток всех каналов в клетке, а не токи через одиночные каналы, то есть разрешающая способность этого метода ниже, чем у конфигураций с оторванной мембраной.
Конфигурация Outside-out
Решить проблему сниженной разрешающей способности метода позволяет конфигурация outside-out (наружная сторона снаружи). Если после перехода в whole-cell mode медленно отводить пипетку от клетки, мембрана не отрывается сразу, а начинает вытягиваться в трубку — это видно на фото 5.

Обратите внимание — в пипетке отчетливо видны кусочки цитоплазмы, попавшие туда после разрушения мембраны при переходе в whole-cell.
Следующая фаза процесса представлена на фото 6.

Мембранная трубка стала совсем тонкой и почти невидима («протуберанец» у поверхности клетки — это небольшая часть цитоплазмы, оставшаяся в мембранной трубке). В следующее мгновение трубка порвётся, а мембрана сомкнется на пипетке в «вывернутом» виде — мы окажемся в конфигурации «outside-out»
Итак, пипетка и её электрод — подобно конфигурации whole-cell — внутриклеточные, мы свободно меняем состав внеклеточного раствора, площадь исследуемой мембраны невелика, так что мы вновь можем изучать одиночные каналы.
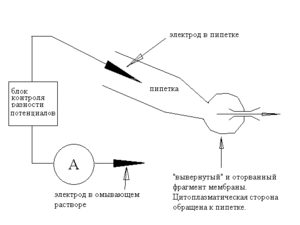
Perforated patch
Perforated patch («перфорированный patch»[10])— это специфический вариант patch-clamp в whole-cell mode. В данном случае, в пипеточном растворе содержится небольшое количество специального антибиотика, например, амфотерицина-В либо грамицидина. Антибиотики этого класса формируют ионные каналы в клеточной мембране на участке, присоединённом к микропипетке.
Такой подход позволяет избежать замещения внутренней среды клетки раствором из пипетки-электрода, то есть клетка остаётся живой с минимальными, насколько это возможно, повреждениями. Таким образом, ответы клетки на раздражители являются максимально приближёнными к естественным. В то же время, данному методу присущ ряд недостатков. Во-первых, по сравнению с классическим whole-cell mode, входное электрическое сопротивление (которое состоит главным образом из сопротивления пипетки и сопротивления в месте соединения пипетки с мембраной) является значительно более высоким. Это понижает качество фиксации потенциала, повышает уровень шума при записи, и увеличивает значения всех ошибок, связанных с изменениями сопротивления полной цепи (от электрода во внешнем растворе до электрода в пипетке). Во-вторых, для того, чтобы антибиотик подействовал, требуется довольно много времени (до 30 минут), что существенно уменьшает полезный период эксперимента. И, в-третьих, антибиотик повреждает мембрану также и в месте соединения с кончиком пипетки, что приводит к ускоренному разрушению гигаомного контакта и дополнительно уменьшает эффективное экспериментальное время. Таким образом, данный вариант метода может быть с успехом использован только в экспериментах, которые не требуют продолжительного времени для изучения исследуемых явлений.
Nucleated patch
Интересная разновидность patch-clamp — nucleated patch. (Фиксация потенциала с клеточным ядром[11]).
Метод состоит в следующем. Пипетка подводится к клетке, а затем рывком пробивает мембрану. После этого кончик пипетки подводится к клеточному ядру, на пипетку подается небольшое отрицательное давление. В результате пипетка присасывается к ядру. Затем пипетка с ядром на конце плавно отводится назад и вынимается из клетки. Пипетку необходимо вывести из клетки таким образом, чтобы участок мембраны в месте выхода «наделся» на присосавшееся ядро и оторвался, обернувшись вокруг него. В результате получается специфический вариант outside-out patch, при котором гораздо больший, чем в обычном варианте метода, участок мембраны присоединён к концу пипетки, будучи обёрнутым вокруг клеточного ядра.
Фиксация потенциала и фиксация тока
Когда изучаются изменения проводимости однотипных каналов в ответ на какие-то химические воздействия или зависимость их проводимости от разности потенциалов на мембране, удобно фиксировать потенциал и измерять ток канала — как мы до сих пор и описывали. Этот, самый частый вариант patch-clamp, называется voltage clamp (фиксация потенциала). Однако, иногда исследователя интересуют процессы трансмембранного переноса ионов, связанные с изменением мембранного потенциала — например, проведение нервного импульса. В таких случаях можно поступить противоположным образом: зафиксировать на постоянном уровне ток, и изучать изменения разности потенциалов при этом. Такой вариант называется current clamp (фиксация тока).
Примеры записей, получаемых методом patch-clamp

Запись одиночного канала рецептора глицина. Чётко видны два состояния: закрытое (ему соответствует нулевой ток) и открытое (ему соответствует ток примерно в 7 пикоампер). Канал время от времени спонтанно переходит из одного состояния в другое, промежуточных состояний нет. Запись позволяет получить две важнейшие характеристики канала: проводимость (исходя из величин трансмембранного потенциала и тока) и вероятность нахождения в открытом состоянии (определяемое как отношение времени, когда канал открыт, ко времени записи тока). К слову, чаще всего активность канала физиологически регулируется путём изменения этой вероятности.

Запись в конфигурации whole-cell. Клетка эпителия собирательной трубки. Трансмембранный потенциал −60 мВ. С интервалами в 1 минуту на 30 секунд в омывающий раствор вводится амилорид — ингибитор эпителиального натриевого канала (Время введения показано темными прямоугольниками). Кстати, чувствительность к ингибиторам — это еще одна из важнейших характеристик канала. Введение амилорида всякий раз приводит к падению тока до нуля, из чего следует, что практически вся проводимость при −60 мВ в данной клетке — это проводимость эпителиального натриевого канала. В противоположность предыдущей записи, изменения тока выглядят непрерывными — это связано с тем, что одновременно записывается много каналов (около 2000), соответственно, имеется примерно 2000 уровней тока — от «все открыты» до «все закрыты».
Planar patch-clamp

Это метод, разработанный для увеличения производительности в электрофизиологии, когда требуются массовые измерения трансмембранной проводимости. В частности, этот метод находит применение в фармакологических исследованиях. Если при классическом patch-clamp пипетка позиционируется на неподвижной клетке, то при планарном, напротив, клеточная суспензия наносится на микрочип с упорядоченными отверстиями субклеточного диаметра. Клетка оседает на отверстие, с помощью насоса подсасывается к нему, в результате чего формируется гигаомный контакт. Далее исследование проводится по тому же принципу, что и в традиционном patch-clamp. [12][13]
Основное преимущество планарного patch-clamp состоит в том, что эксперимент выполняется значительно проще, требует меньшей квалификации от исследователя и занимает меньше времени. Достаточно много измерений может быть выполнено одно за другим в автоматическом режиме. По утверждению авторов методики, этот метод позволяет легче осуществлять перфузию внутриклеточного содержимого, а также более точно дозировать исследуемые или тестовые фармакологические вещества. С помощью этого метода относительно легко работать с клетками, обычно находящимися во взвешенном состоянии — в частности, с клетками крови.
Вместе с тем, метод планарного patch-clamp имеет ряд существенных ограничений. Основное состоит в том, что исследуемые клетки должны находиться в суспензии. Соответственно, не существует возможности работать с фрагментами ткани, невозможно изучать взаимодействие клеток друг с другом. Для мобилизации клеток необходимо использовать ферменты-протеазы, которые могут модифицировать поведение изучаемых ионных каналов.
Все вышесказанное приводит к тому, что планарный patch-clamp находит применение главным образом в фармакологических скрининговых исследованиях, где требуются массовые измерения, но не столь распространен в области фундаментальной науки.
Примечания
- O. P. Hamill, A. Marty, E. Neher, B. Sakmann, and F. J. Sigworth. Improved Patch-Clamp Techniques for High-Resolution Current Recording from Cells and Cell-Free Membrane Patches. // Pfluegers Arch (1981)391:85-100
- Физиология человека. Под ред. В. М. Покровского, Г. Ф. Коротько. М.Медицина 2007. ISBN 5-225-04729-7
- Р.Геннис. Биомембраны — Молекулярная структура и функции, стр. 336-337. «Мир» 1997. ISBN 5-03-002419-0
- www.ionchannels.org (недоступная ссылка). Дата обращения: 28 августа 2006. Архивировано 17 мая 2014 года.
- Lu M, Dong K, Egan ME, Giebisch GH, Boulpaep EL, Hebert SC. Mouse cystic fibrosis transmembrane conductance regulator forms cAMP-PKA-regulated apical chloride channels in cortical collecting duct. //Proc Natl Acad Sci U S A. 2010 Mar 30;107(13):6082-7.
- Anantharam A, Palmer LG. Determination of epithelial Na+ channel subunit stoichiometry from single-channel conductances // J.Gen.Physiol. 2007, v.130, p. 55-70
- Р.Геннис. Биомембраны — Молекулярная структура и функции, стр. 349. «Мир» 1997. ISBN 5-03-002419-0
-
- Microelectrode techniques: The Plymouth workshop handbook., стр.20-27 — изд. 2-е. — Cambridge, UK: Company of Biologists, 1994. — ISBN 0-948601-49-3
- B. Sakmann, E. Neher. Single-channel recording. — 1995. ISBN 0-306-44870-X
- A. Molleman. Patch-clamping: an introductory guide to Patch-Clamp electrophysiology. P. 108-110. — John Wiley and Sons, 2003. ISBN 978-0-471-48685-5
- Uwe Windhorst,Håkan Johansson. Modern techniques in neuroscience research. P.190-192. — Springer, 1999
- Jan C. Behrends & Niels Fertig: Planar Patch Clamping. In: Neuromethods. Bd. 38, S. 411—433. PDF
- Архивированная копия (недоступная ссылка). Дата обращения: 14 ноября 2008. Архивировано 5 апреля 2008 года.
Литература
- Регистрация одиночных каналов = Single-channel recording / Под ред. Б.Сакмана, Э.Неера. — М.: «Мир», 1987. — 2750 экз.
- Numberger M., Draguhn A. Patch-Clamp Technik. — 1996.
- Jan C. Behrends & Niels Fertig. Planar Patch Clamping, Neuromethods, S. 411-433. PDF.
- The Axon Guide For Electrophysiology & Biophysics Laboratory Techniques
- П.Г. Костюк, В.Л. Зима, І.С. Магура, М.С. Мірошниченко, М.Ф. Шуба. Біофізика / За редакцією П.Г. Костюка. — Київ: «Обереги», 2001. — 1000 экз. — ISBN 966-513-021-8.
Ссылки
- Сайт об исследовании ионных каналов Архивная копия от 17 мая 2014 на Wayback Machine