3′-Нетранслируемая область
3′-Нетранслируемая область (3′-НТО, англ. 3′-untranslated region, 3′-UTR) — некодирующий участок мРНК, располагающийся на её 3′-конце после кодирующей области. Такое же название имеет участок ДНК, соответствующий 3′-UTR транскрипта[1]. 3′-UTR может принимать участие в регуляции эффективности трансляции, стабильности мРНК, содержать сигналы полиаденилирования[2] и сайты связывания микроРНК, а также выполнять ряд других регуляторных функций.

Структура
Длина и нуклеотидный состав
Длина 3′-UTR может составлять от 60 до 4000 нуклеотидов. Средняя длина 3′-UTR у человека составляет около 800 нуклеотидов, в то время как средняя длина 5′-UTR — 200 нуклеотидов[3]. Примечательно, что общая длина 3′-UTR у человека более чем в два раза превосходит таковую у остальных млекопитающих, что свидетельствует о большем, чем у других млекопитающих, числе регуляторных элементов у людей[4]. Состав оснований также различается в 3′- и 5′-UTR. Так, в 5′-UTR выше содержание G+C, чем в 3′-UTR. Особенно это различие заметно в мРНК теплокровных позвоночных, у которых содержание G+C в 5′-UTR составляет 60 %, а в 3′-UTR — 45 %[5][6].
Длина и вторичная структура 3′-UTR в значительной мере определяются её участием во взаимодействиях 5’-конца транскрипта с 3’-концом (см. ниже), и нередко длинные 3′-UTR оказывают значительное влияние на экспрессию генов. В 1996 году было показано, что увеличение 3′-UTR мРНК с 19 до 156 нуклеотидов уменьшало экспрессию в 45 раз, вне зависимости от ориентации, гена или последовательности вставленных нуклеотидов. Это свидетельствует о том, что длина 3′-UTR имеет важное значение в экспрессии мРНК. Другим фактором, определяющим важность длины 3′-UTR, кроме взаимодействия 3′- и 5′-UTR, является способность 3’-UTR к взаимодействию с микроРНК — особыми регуляторными молекулами РНК, подавляющими трансляцию (подробнее см. ниже). Эти взаимодействия осуществляются на специальных сайтах, которых больше в длинных 3′-UTR, поэтому длинная 3′-UTR может оказывать более сильный подавляющий эффект на трансляцию. Так, было проведено сравнение длины 3′-UTR и количества сайтов связывания с микроРНК на ней у генов рибосомных белков и генов, участвующих в нейрогенезе. Оказалось, что у генов рибосомных 3′-UTR короче и меньше специфических сайтов связывания с микроРНК, а у генов, вовлеченных в нейрогенез, наоборот, 3′-UTR длиннее и содержит много специфических сайтов связывания с микроРНК. Рассмотрим другой пример. Ген Hip2 использует альтернативные 3′-UTR для гибкого контроля экспрессии (подробнее об этом явлении см. ниже). Более длинная из возможных 3′-UTR этого гена содержит консервативные сайты связывания двух микроРНК, экспрессирующихся в активированных Т-клетках. При активации относительная экспрессия транскрипта с более длинной 3′-UTR снижалась, а общая экспрессия белка увеличивалась, так как экспрессировались мРНК с более короткими 3′-UTR, не содержащие сайтов связывания для подавляющих микроРНК. Показано также, что длина 3′-UTR зависит от наличия в ней таких регуляторных элементов, как AU-богатые элементы (ARE) (подробнее см. ниже)[4].
В общем, длинные 3′-UTR приурочены к относительно невысокому уровню экспрессии, как было показано в экспериментах, в которых проводилось сравнение экспрессии изоформ одного белка, чьи мРНК различались лишь по длине 3′-UTR. Ген SLC7A1 экспрессируется в две мРНК с различающимися 3′-UTR, причём более длинная из них содержит дополнительный сайт связывания микроРНК. Функциональный полиморфизм в этом гене связан с появлением дисфункции эндотелия и наследственной предрасположенностью к гипертонии. Интересно, что аллель, отвечающий за проявление этих расстройств, обычно имеет более длинную 3′-UTR, в связи с чем его уровень экспрессии ниже, чем у аллеля дикого типа, имеющего более короткую 3′-UTR[4].
Интроны
В отличие от 5′-UTR, 3′-UTR содержат относительно мало интронов (около 5 %). У некоторых генов млекопитающих, появившихся в результате обратной транскрипции со сплайсированного транскрипта, имеются интроны в 3′-UTR, которые снижают экспрессию этих генов, направляя их транскрипты на путь NMD (то есть на разрушение). Этот отрицательный эффект интронов в 3′-UTR на экспрессию генов, возможно, объясняет их малое распространение в этой области. Более того, установлено, что некоторые транскрипты способны связываться с микроРНК только при наличии интрона в 3′-UTR, что также подавляет экспрессию гена. Это показывает, что различное вырезание интронов в 3′-UTR даёт возможность для изоформо-специфичной регуляции, опосредованной микроРНК, которая может осуществляться тканеспецифично[7].
Вторичная структура
По-видимому, вторичная структура 3’-UTR имеет гораздо большее значение, чем считалось ранее. Важна не только длина 3’-UTR, но и её вторичная структура, и мутации, изменяющие её, могут нарушить экспрессию гена. В 2006 году было проведено исследование 83 вариантов 3′-UTR, приуроченных к различным заболеваниям, и была установлена связь между функциональностью этих вариантов и изменениями в предсказанной вторичной структуре[8].
Вторичную структуру 3′-UTR сложно предсказать, так как множество связывающихся с ней белковых факторов могут значительно влиять на её пространственную структуру. Эти факторы могут изменять её из-за разрушения укладки мРНК, а могут взаимодействовать с другими факторами, из-за чего мРНК может замкнуться в петлю. Наиболее распространённым примером элементов вторичной структуры, способных влиять на экспрессию, является шпилька, и в 3’-UTR именно со шпильками связываются РНК-связывающие белки. Транскрипт нейротрофического фактора мозга (англ. brain-derived neurotrophic factor, BDNF) содержит длинную шпильку, ответственную за стабильность мРНК в нейронах в ответ на кальциевые сигналы. Предполагается, что шпилька представляет собой удобную площадку для взаимодействия ряда РНК-связывающих белков, некодирующих РНК и сигналов полиаденилирования в ответ на Са2+. У транскрипта TNFα в 3′-UTR располагается элемент ARE, образующий шпильку, которая способна модулировать сродство этого участка к различными белкам (подробнее см. ниже). Эти примеры показывают, что модуляция вторичной структуры 3’-UTR при помощи белков или иных средств может изменять её специфичность к связыванию с различными транс'′-факторами, тем самым регулируя экспрессию генов на пост-транскрипционном уровне[9].
Альтернативные 3′-UTR
Альтернативное полиаденилирование (англ. alternative polyadenilation, APA) и альтернативный сплайсинг являются двумя механизмами, приводящими к появлению различных изоформ мРНК, различающихся своими 3′-UTR. APA может происходить из-за наличия разных сайтов полиаденилирования и различных терминальных экзонов; APA используют предположительно ~50 % человеческих генов. Этот механизм очень удобен для сложных организмов, так как он даёт возможность транскриптам экспрессироваться в один и тот же белок, но на различном уровне и в различной пространственной локализации из-за различий в регуляции, опосредованной 3′-UTR. Альтернативные 3′-UTR чрезвычайно важны для тканеспецифичной экспрессии генов, а также для различной экспрессии на разных этапах развития. Значительные изменения продуктов АРА характерны для ряда видов рака. АРА также играет важную роль в локализации изоформ белка. Белковый продукт гена HuR является ARE-связывающим белком, участвующим в стабилизации многих мРНК, содержащих ARE. Благодаря АРА образуется ряд вариантов белка HuR, различающихся по уровню экспрессии, и, хотя у подавляющего большинства транскриптов этого белка отсутствует ARE, у некоторых всё-таки есть функциональные ARE в 3′-UTR. Эти ARE способны связывать HuR, тем самым осуществляя регуляцию по типу положительной обратной связи. Таким образом, использование альтернативных 3′-UTR даёт возможность для ещё большего разнообразия белковых продуктов одного гена[10].
Функции
Взаимодействие с микроРНК
МикроРНК — короткие одноцепочечные некодирующие молекулы РНК эндогенного происхождения длиной около 20 нуклеотидов. Они взаимодействуют с мРНК-мишенями по принципу комплементарности и обычно блокируют трансляцию мишени или вызывают её разрушение. Как правило, сайты связывания микроРНК с мРНК локализованы в 3′-UTR последней, хотя некоторые из них находятся в 5′-UTR и даже кодирующей области. МикроРНК часто экспрессируются по-разному в зависимости от типа ткани и стадии развития, и генам, вовлечённым в общие для всех генов процессы, приходится селективно избегать последовательностей в транскриптах, частично комплементарных микроРНК, то есть избегать наличия сайтов связывания с микроРНК. Этот процесс селективного избегания оказывает огромное влияние на эволюцию 3′-UTR[11].

Стабилизация мРНК
Изменение стабильности транскрипта позволяет осуществлять быстрый контроль экспрессии без изменения темпов трансляции. Такой механизм имеет важное значение в таких жизненно важных процессах, как клеточный рост и дифференциация, а также адаптация к условиям окружающей среды. Наиболее хорошо изученными регуляторными элементами, регулирующими стабильность мРНК, являются AU-богатые элементы (англ. AU-rich elements, AREs), располагающиеся в 3′-UTR мРНК некоторых генов. Эти элементы имеют размер от 50 до 150 нуклеотидов и обычно содержат многочисленные копии пентануклеотида AUUUA[12].
Было установлено, что последовательности AREs различаются, и по количеству и расположению мотивов AUUUA выделяют 3 класса AREs:
- I класс: 1—3 разбросанно расположенных AUUUA внутри U-богатого участка;
- II класс: множество перекрывающихся мотивов AUUUA;
- III класс: AUUUA нет, но есть U-богатые участки[12].
AREs связываются с белками (англ. ARE-binding proteins, ARE-BPs), которые, как правило, способствуют разрушению мРНК в ответ на различные внутри- и внеклеточные сигналы, хотя некоторые из них регулируют трансляцию. AREs регулируют экспрессию генов, кодирующих цитокины, факторы роста, генов-супрессоров опухолей, протоонкогенов, а также генов, чьи белковые продукты участвуют в регуляции клеточного цикла, например, гены циклинов, ферментов, факторов транскрипции, рецепторов и мембранных белков. Такое разнообразие генов, чьи транскрипты содержат AREs, свидетельствует о важности стабильности транскрипта в регуляции гена[12]. Кроме изменения стабильности мРНК, ARE могут также активировать трансляцию, хотя этот механизм менее распространен и хуже понятен[13].
Другим элементом, регулирующим стабильность транскрипта, является недавно открытый GU-богатый элемент (GRE). Он взаимодействует с CUGBP1 — РНК-связывающим белком, способствующим распаду связанной с ним мРНК[13].
Участие в полиаденилировании
Полиаденилирование представляет собой процесс добавления серии аденозинов (то есть поли(А)-хвоста) к 3′-концу ещё незрелого РНК-транскрипта[13]. Установлено, что в 3′-UTR располагаются элементы, регулирующие этот процесс. Так, показано, что во всех полиаденилируемых мРНК на расстоянии 20—30 нуклеотидов от 3′-конца транскрипта, к которому присоединяется поли(А)-хвост, содержится последовательность AAUAAA — сигнал полиаденилирования (сигналами полиаденилирования могут также выступать близкие последовательности, такие как AU/GUAAA или UAUAAA). Впоследствии оказалось, что, хотя последовательность AAUAAA абсолютно необходима для полиаденилирования, существуют и другие элементы, без которых нормальное присоединение поли(А)-хвоста невозможно. В частности, была выявлена GU-богатая последовательность, располагающаяся сразу после AAUAAA в направлении к 3' концу (её также называют англ. downstream sequence element, DSE), а также особая последовательность, располагающаяся сразу перед AAUAAA (англ. upstream sequence element, USE). Эти элементы в значительной мере консервативны не только для млекопитающих, но и для всех эукариот. Для полиаденилирования важны также нуклеотиды, располагающиеся в месте разрезания 3′-конца транскрипта (к этому месту после разрыва будет присоединён поли(А)-хвост). Таким образом, 3′-UTR играет важнейшую роль в процессе полиаденилирования[14].
Участие в маскировании мРНК
3′-UTR играет существенную роль в процессе маскирования мРНК. Маскирование мРНК происходит, например, в процессе оогенеза и сперматогенеза, когда синтезированные в течение этих процессов мРНК не транслируются в белок, а хранятся в неактивном состоянии, иногда довольно долго. При оплодотворении и в ходе раннего эмбриогенеза происходит демаскирование материнских мРНК, и с них синтезируются необходимые белки. Маскирование и хранение мРНК происходит также в дифференцирующихся соматических клеток взрослого организма в течение длительного времени[15].
Впервые явление маскирования мРНК было изучено у двустворчатого моллюска Spisula solidissima в 1990 году. Оказалось, что в его ооцитах хранится большое количество маскированных мРНК, кодирующих малую субъединицу рибонуклеотидредуктазы и циклин А. Было показано, что, когда мРНК находится в маскированном состоянии, с участком в её 3′-UTR связан комплекс маскирующих белков. Выяснилось также, что маскированные мРНК имеют сильно укороченный поли(А)-хвост — с 200—250 адениловых остатков до 20—40. При демаскировании мРНК маскирующие белки фосфорилируются, в результате чего кэп освобождается от блокирующего белка и стимулируется полиаденилирование мРНК цитоплазматической поли(А)-полимеразой, восстанавливающей длинный поли(А)-хвост, необходимый для эффективной трансляции[16].
Вставка селеноцистеина
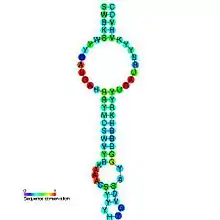
3′-UTR иногда участвует в процессе включения в полипептидную цепь редкой, но функционально важной аминокислоты — селеноцистеина. Особого кодона для селеноцистеина нет, и присоединение тРНКSec осуществляется к терминирующему кодону UGA, но только тогда, когда за ним находится особая последовательность вставки селеноцистеина — SECIS, формирующая характерный элемент вторичной структуры. SECIS может находиться на значительном удалении (до 200 нуклеотидов) от UGA и у архей и эукариот локализуется в 3′-UTR мРНК[17][18].
Участие в NMD
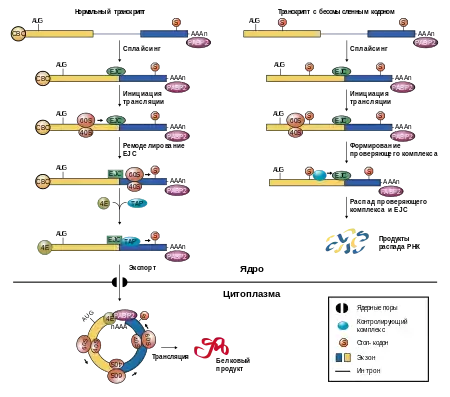
NMD (англ. nonsense-mediated decay) является действенным механизмом разрушения нефункциональных мутантных транскриптов. Обычно в этом механизме эффективность определяется расположением мутации относительно места соединения экзонов, однако 3′-UTR также может иметь некоторое значение. Механизм терминации трансляции на преждевременных стоп-кодонах зависит от расстояния между терминаторным кодоном и поли(А)-связывающим белком PABPC1. Показано, что увеличение расстояния между стоп-кодоном и поли(А)-хвостом приводит к запуску NMD, а изменения в пространственной структуре 3’-UTR могут модулировать NMD[8].
Взаимодействие 5′-UTR и 3′-UTR
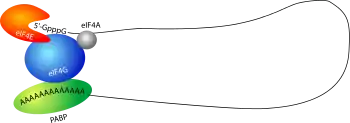
Известно, что мРНК способна замыкаться в кольцо (циркуляризация) за счёт взаимодействия специальных белков, связывающихся с поли(А)-хвостом, способствующих связыванию фактора eIF4F с кэпом. В результате мРНК приобретает замкнутую форму, стимулируется инициация трансляции и повышается эффективность трансляции. Однако в некоторых случаях связываться друг с другом могут 5′-UTR и 3′-UTR одной и той же мРНК. Так, у мРНК человеческого гена p53 имеются участки в 5′-UTR и 3′-UTR, комплементарные друг другу. Связываясь друг с другом и с трансляционным фактором RPL26, они тем самым способствуют повышению эффективность трансляции белка р53 в ответ на повреждения ДНК[8].
Анализ мРНК различных генов человека показал, что в 5′-UTR присутствует мотив, специфически взаимодействующий с 3′-концами микроРНК, при этом во многих таких мРНК на 5′-конце имеется сайт, комплементарный 3′-UTR. Дальнейшие исследования показали, что связывание 5′-UTR с микроРНК облегчает связь 5′-конца мРНК с 3′-концом, и мРНК, активность которых значительно определяется микроРНК, имеют предсказуемые сайты связывания на обеих UTR. Такие мРНК получили название miBridge. Далее было установлено, что утрата этих сайтов связывания уменьшала репрессию трансляции транскрипта, управляемую микроРНК. Так было выяснено, что сайты связывания UTR друг с другом необходимы для подавления трансляции мРНК. Это свидетельствует о том, что комплементарное взаимодействие 5′-UTR и 3′-UTR необходимо для точной регуляции экспрессии генов[9].
3′-UTR прокариот и вирусов
Бактерии
В мРНК бактерий также имеются 5′- и 3′-нетранслируемые области[19][20].
В отличие от эукариот, длинные 3′-UTR редки у бактерий и слабо изучены. Тем не менее, известно, что у некоторых бактерий, в частности, Salmonella enterica, имеются мРНК с длинными 3′-UTR, похожими на эукариотические (у S. enterica это мРНК hilD). Предполагается, что 3′-UTR hilD выполняют различные функции, в частности, влияют на обращение своих мРНК, поскольку делеция этих областей вызывала увеличение количества соответствующих мРНК[21].
Археи
Нетранслируемые области имеются и в мРНК многих архей. В частности, в 5′- и 3′-UTR мРНК метаногенной археи Methanococcus jannaschii (как и у других представителей порядков Methanopyrales и Methanococcales) локализован элемент SECIS, ответственный за вставку аминокислоты селеноцистеина в полипептидную цепь[22].
Установлено, что мРНК большинства галоархей, а также Pyrobaculum и Sulfolobus лишены выраженной 5′-UTR, но у мРНК архей-метаногенов имеются длинные 5′-UTR. В связи с этим предполагается, что механизм инициации трансляции метаногенных архей может быть отличен от такового у остальных представителей этого домена[23]. Тем не менее, в мРНК галоархей имеются 3′-UTR и их 3′-концы не подвергаются посттранскрипционной модификации. Удивительным является тот факт, что в тех транскриптах галоархей, у которых есть 5′-UTR, отсутствует последовательность Шайна — Дальгарно. Длина же 3′-UTR галоархей составляла от 20 до 80 нуклеотидов; никаких консервативных структурных мотивов и последовательностей, кроме пента-U-нуклеотида в области терминации трансляции, выявлено не было[24].
Вирусы
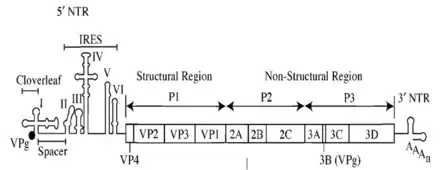
У многих вирусов инициация трансляции происходит по кэп-независимому механизму и осуществляется через элементы IRES, локализованные в 5′-UTR[25]. Тем не менее, у вирусов обнаружен и другой кэп-независимый механизм инициации трансляции, не связанный с IRES. Такой механизм имеется у многих вирусов растений. В этом случае имеется особый кэп-независимый трансляционный элемент (англ. cap-independent translation element (CITE)), расположенный в 3′-UTR. Нередко CITE связывает факторы трансляции, например, комплекс eIF4F, и затем комплементарно взаимодействует с 5′-концом, доставляя факторы инициации трансляции к месту её начала[26].
У вирусов, геном которых представлен одноцепочечной молекулой РНК положительной полярности, 3′-UTR не только оказывает влияние на трансляцию, но также задействована в репликации: именно с неё начинается репликация вирусного генома[27].
Вирус кори (род Morbillivirus семейства Paramyxoviridae) имеет геном, представленный одноцепочечной молекулой РНК отрицательной полярности. Для его генов М и F был установлен интересный механизм. мРНК этих генов имеют длинные UTR, на их долю приходится ~6,4 % всей мРНК. Хотя эти гены непосредственно не участвуют в репликации, 3′-UTR мРНК гена М увеличивает экспрессию белка М и тем самым запускает репликацию генома. В то же время 5′-UTR мРНК гена F снижает образование белка F и тем самым подавляет репликацию[28].
Методы изучения
При изучении структуры и функций 3′-UTR ученые используют несколько различных методов. Даже если для данной 3′-UTR показано наличие её в определённой ткани, для получения полного представления о её функциях необходимо проанализировать эффекты её различной локализации, выяснить срок функционирования, описать взаимодействия с транс-регуляторными белками, влияние на эффективность трансляции[29]. С помощью методов биоинформатики на основании анализа первичной структуры (то есть последовательности нуклеотидов) можно искать элементы ARE и сайты связывания с микроРНК в данной 3′-UTR. Экспериментальными методами устанавливаются последовательности, взаимодействующие с теми или иными транс-регуляторными белками, и в настоящий момент на основании данных секвенирования и экспериментальных данных возможно находить сайты взаимодействия с определёнными белками в данном транскрипте[30]. Искусственно индуцируя мутации в 3′-UTR, например, затрагивающие терминаторный кодон, сигнал полиаденилирования или вторичную структуру 3′-UTR, можно установить, как мутации в этих участках могут приводить к нарушениям трансляции и появлению болезней (подробнее о заболеваниях, ассоциированных с 3′-UTR, см. ниже)[31]. Итак, с помощью всех этих методов мы можем развивать наши представления о структуре и функциях цис-регуляторных элементов в 3′-UTR, а также взаимодействующих с 3′-UTR белков.
Клиническое значение
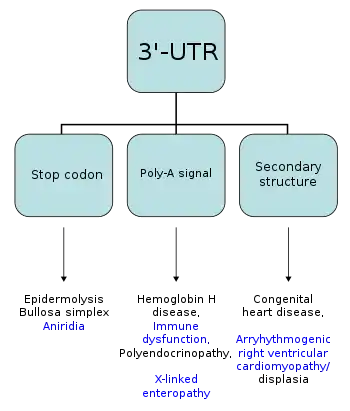
Мутации, затрагивающие 3′-UTR, имеют важное значение, поскольку одна такая мутация может сказаться на экспрессии многих генов. Хотя на уровне транскрипции мутации влияют на конкретный аллель и физически сцепленные гены, поскольку связывающиеся с 3′-UTR белки также принимают участие в процессинге и экспорте МРНК из ядра. Таким образом, мутация может влиять на несвязанные гены [32]. Так, мутации, произошедшие в ARE, приводят к сбою в работе ARE-связывающих белков, в результате чего могут развиться такие заболевания, как злокачественные перерождения кроветворных органов и лейкемия[33][34]. Повышенное содержание тринуклеотида CTG в 3′-UTR гена миотониновой протеинкиназы вызывает миотоническую дистрофию. Вставка ретротранспозона длиной 3 килобаз, состоящего из тандемных повторов, в 3′-UTR гена белка фукутина, связана с врожденной мышечной дистрофией типа Фукуяма[29]. Изменения в элементах, локализованных в 3′-UTR, связаны с развитием таких заболеваний человека, как острый миелоидный лейкоз, альфа-талассемия, нейробластома, кератинопатия, аниридия, синдром IPEX, врождённые пороки сердца[31]. Связь некоторых из этих заболеваний с конкретными элементами 3′-UTR представлена на схеме ниже.
Примечания
- Barrett et. al., 2013, p. 9.
- Molecular biology glossary: 3' Untranslated Region (3' UTR).
- Mignone, Flavio; Graziano Pesole. mRNA Untranslated Regions (UTRs) (неопр.). — 2011. — 15 August. — doi:10.1002/9780470015902.a0005009.pub2.
- Barrett et. al., 2013, p. 31.
- Pesole G, Liuni S, Grillo G, Saccone C. Structural and compositional features of untranslated regions of eukaryotic mRNAs. (англ.) // Gene. — Elsevier, 1997. — Vol. 205, no. 1—2. — P. 95—102.
- Здесь и далее в разделах «Структура» и «Функции» приводится информация по эукариотическим клеточным 5′-UTR. Данные по 5′-UTR бактерий, архей и вирусов рассматриваются в соответствующем разделе.
- Barrett et. al., 2013, p. 21—22.
- Barrett et. al., 2013, p. 32.
- Barrett et. al., 2013, p. 32—33.
- Barrett et. al., 2013, p. 33.
- Barrett et. al., 2013, p. 25—27.
- Barrett et. al., 2013, p. 28.
- Barrett et. al., 2013, p. 29.
- Nick J. Proudfoot. Ending the message: poly(A) signals then and now // Genes & Dev.. — 2011. — Т. 25. — С. 1770—1782. — doi:10.1101/gad.17268411.
- Спирин, 2011, с. 416.
- Спирин, 2011, с. 418.
- Коничев, Севастьянова, 2012, с. 328.
- Berry, M. J.; Banu, L.; Harney, J. W.; Larsen, P. R. Functional Characterization of the Eukaryotic SECIS Elements which Direct Selenocysteine Insertion at UGA Codons (англ.) // The EMBO Journal : journal. — 1993. — Vol. 12, no. 8. — P. 3315—3322. — PMID 8344267.
- Льюин Б. Гены. — БИНОМ, 2012. — С. 144. — 896 с. — ISBN 978-5-94774-793-5.
- Н. В. Равин, С. В. Шестаков. Геном прокариот // Вавиловский журнал генетики и селекции. — 2013. — Т. 17, № 4/2. — С. 972—984.
- Javier López-Garrido, Elena Puerta-Fernández, Josep Casadesús. A eukaryotic-like 3′ untranslated region in Salmonella enterica hilD mRNA // Nucl. Acids Res.. — 2014. — ISSN 1362-4962. — doi:10.1093/nar/gku222.
- R. Wilting, S. Schorling, B. C. Persson, A. Bock. Selenoprotein Synthesis in Archaea: Identification of an mRNA Element of Methanococcus jannaschii Probably Directing Selenocysteine Insertion // J. Mol. Biol.. — 1997. — Т. 266. — С. 637—641.
- Jian Zhang. Gene expression in Archaea: Studies of transcriptional promoters, messenger RNA processing, and five prime untranslated regions in Methanocaldococcus jannashchii. — 2009. Архивировано 31 мая 2014 года.
- Brenneis M., Hering O., Lange C., Soppa J. Experimental characterization of Cis-acting elements important for translation and transcription in halophilic archaea. // PLoS Genet.. — 2007. — Т. 3, № 12. — doi:10.1371/journal.pgen.0030229.
- Thompson, Sunnie R. Tricks an IRES uses to enslave ribosomes (англ.) // Trends in Microbiology : journal. — Cell Press, 2012. — Vol. 20, no. 11. — P. 558—566. — doi:10.1016/j.tim.2012.08.002. — PMID 22944245.
- Qiuling Fan, Krzysztof Treder, W Allen Miller. Untranslated regions of diverse plant viral RNAs vary greatly in translation enhancement efficiency // BMC Biotechnology. — 2012. — Т. 12, № 22. — doi:10.1186/1472-6750-12-22.
- Dreher T. W. FUNCTIONS OF THE 3′-UNTRANSLATED REGIONS OF POSITIVE STRAND RNA VIRAL GENOMES // Annu Rev Phytopathol.. — 1999. — Т. 37. — С. 151—174.
- Makoto Takeda, Shinji Ohno, Fumio Seki, Yuichiro Nakatsu, Maino Tahara, Yusuke Yanagi. Long Untranslated Regions of the Measles Virus M and F Genes Control Virus Replication and Cytopathogenicity // J. Virol.. — 2005. — Т. 79, № 22. — С. 14346—14354. — doi:10.1128/JVI.79.22.14346-14354.2005.
- Conne, Béatrice; Stutz, André; Vassalli, Jean-Dominique. The 3' untranslated region of messenger RNA: A molecular 'hotspot' for pathology? (англ.) // Nature Medicine : journal. — 2000. — 1 June (vol. 6, no. 6). — P. 637—641. — doi:10.1038/76211.
- Zhao, W.; Blagev, D.; Pollack, J. L.; Erle, D. J. Toward a Systematic Understanding of mRNA 3' Untranslated Regions (англ.) // Proceedings of the American Thoracic Society : journal. — 2011. — 4 May (vol. 8, no. 2). — P. 163—166. — doi:10.1513/pats.201007-054MS.
- Sangeeta Chatterjee, Jayanta K. Pal. Role of 5- and 3-untranslated regions of mRNAs in human diseases // Biol. Cell. — 2009. — С. 251—262. — doi:10.1042/BC20080104. (недоступная ссылка)
- Chatterjee, Sangeeta; Pal, Jayanta K. Role of 5′- and 3′-untranslated regions of mRNAs in human diseases (англ.) // Biology of the Cell : journal. — 2009. — 1 May (vol. 101, no. 5). — P. 251—262. — doi:10.1042/BC20080104.
- Baou, M.; Norton, J. D.; Murphy, J. J. AU-rich RNA binding proteins in hematopoiesis and leukemogenesis (англ.) // Blood. — American Society of Hematology, 2011. — 13 September (vol. 118, no. 22). — P. 5732—5740. — doi:10.1182/blood-2011-07-347237.
- Khabar, Khalid S. A. Post-transcriptional control during chronic inflammation and cancer: a focus on AU-rich elements (англ.) // Cellular and Molecular Life Sciences : journal. — 2010. — 22 May (vol. 67, no. 17). — P. 2937—2955. — doi:10.1007/s00018-010-0383-x.
Литература
- Спирин А. С. Молекулярная биология. Рибосомы и биосинтез белка. — М.: Издательский центр «Академия», 2011. — 496 с. — ISBN 978-5-7695-6668-4.
- Коничев А. С., Севастьянова Г. А. Молекулярная биология. — Издательский центр «Академия», 2012. — 400 с. — ISBN 978-5-7695-9147-1.
- Lucy W. Barrett, Sue Fletcher, Steve D. Wilton. Untranslated Gene Regions and Other Non-coding Elements. — SpringerBriefs in Biochemistry and Molecular Biology, 2013. — 57 p. — ISBN 978-3-0348-0679-4.