Энтоз
Энто́з (англ. entosis) — вид программируемой клеточной гибели, при котором одна эпителиальная клетка поглощается другой эпителиальной клеткой и впоследствии умирает в вакуоли или лизосоме поглотившей клетки. Энтоз часто наблюдается в опухолях, потому что он запускается при утрате контактов клетки с внеклеточным матриксом, что наиболее часто наблюдается у раковых клеток[1]. Показано также, что энтоз играет важную роль в эмбриональном развитии млекопитающих.
К другим видам клеточной гибели, которые сопровождаются интернализацией клетки внутри другой клетки, относят фагоптоз (например, поглощение нейронов клетками глии), суицидальный эмпериполез (например, T-лимфоциты внутри гепатоцитов), эмперитоз (натуральные киллеры внутри опухолевых клеток), каннибализм (например, лимфоциты в клетках метастазов меланомы), гомотипический клеточный каннибализм (клетки аденокарциномы поджелудочной железы)[2].
Необычные структуры, представляющие собой клетку внутри другой клетки, описал Карл Йозеф Эберт в 1864 году. Он нашёл эпителиальные клетки, содержащие внутри лимфоциты[3]. Энтоз впервые был охарактеризован Майклом Оверхольтцером с соавторами в 2007 году и описан в качестве формы «клеточного каннибализма» у лимфобластов, которые были выделены у пациентов с болезнью Хантингтона[4]. В 2009 году Комитет номенклатуры видов клеточной гибели (англ. Nomenclature Committee on Cell Death) включил энтоз в свой перечень[5]. Предлагается рассматривать его как IV тип клеточной гибели[6].
Распространение
Процесс поглощения одной клетки другой можно наблюдать при споруляции у бактерий и хищных амёб, которые таким образом спасаются от голодания. Поглощение одной клетки другой клеткой нередко встречается и у настоящих многоклеточных животных (Metazoa). Например, оно часто сопровождает гибель клеток при развитии нематоды Caenorhabditis elegans, его можно наблюдать у питающих клеток дрозофилы. У млекопитающих описаны разные варианты структур, представляющих собой одну клетку внутри другой, причём обычно эти описания были выполнены патологами. Например, были обнаружены тимоциты внутри питающих клеток тимуса, иммунные клетки внутри мегакариоцитов. Особенно часто подобные структуры встречаются в опухолях. Таким образом, клеточный каннибализм — древний процесс, консервативный на протяжении эволюции[2].
Морфология
На основании морфологических черт выделяют пять стадий энтоза. На первой стадии поглощённая клетка имеет округлую форму и сохраняет естественный размер. Её плазматическая мембрана близко контактирует с мембраной энтотической вакуоли. На второй стадии поглощённая клетка начинает сжиматься и образует короткие выросты, направленные к мембране энтотической вакуоли. Ядро остаётся округлым, хроматин диффузный, ядрышки хорошо различимы. На третьей стадии размер поглощённой клетки продолжает уменьшаться, сама клетка и её ядро приобретают неправильную форму. В цитоплазме накапливаются вакуоли, хроматин конденсируется, ядрышки становятся видны менее отчётливо. На четвёртой стадии поглощённая клетка и её ядро всё больше деформируются, продолжается интенсивное формирование цитоплазматических вакуолей, ядрышки исчезают. На последней, пятой стадии внутри клетки-хозяина остаются лишь остатки поглощённой клетки. Наиболее часто в культуре клеток выявляются случаи энтоза на второй стадии[5].
Механизм
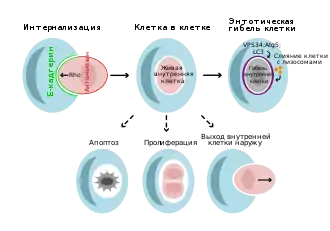
Описано 2 типа энтоза — гомо- и гетеротипический, при которых взаимодействуют клетки, соответственно, одного либо разных типов[7][8]. Некоторые исследователи считают энтозом только те случаи, когда взаимодействуют клетки одного типа[9].
Энтоз запускается при откреплении клеток от внеклеточного матрикса, как при другой форме программируемой клеточной гибели — аноикисе, хотя механизмы этих видов программируемой смерти клеток существенно различаются. По этой причине энтоз в основном наблюдается у раковых клеток, так как они, как правило, не прикреплены ни к внеклеточному матриксу, ни друг к другу. Однако энтоз был обнаружен и у полностью прикреплённых эпителиальных клеток. Было показано, что ему могут способствовать митоз и нехватка глюкозы[10][2].
В отличие от фагоцитоза, в энтозе принимают участие эпителиальные адгезионные контакты, в состав которых входит рецептор межклеточной адгезии E-кадгерин, а также белок, который связывает адгезионные контакты с цитоскелетом, — α-катенин. Е-кадгерин и α-катетин необходимы и достаточны для энтоза в клетках рака молочной железы. Клетки, поглощаемые путём энтоза, в отличие от фагоцитируемых клеток, принимают активное участие в поглощении себя при помощи ГТФазы RhoA и эффекторных киназ ROCKI и ROCKII. Сверхэкспрессии RhoA или ROCKI/II достаточно для запуска поглощения эпителиальных клеток, экспрессирующих кадгерин, поэтому энтоз более напоминает инвазивную деятельность клеток и образование клетки-в-клетке, чем простое поглощение. В самом деле, клетка, интернализующаяся по пути энтоза, демонстрирует блеббинг (англ. blebbing), свойственный инвазивным клеткам, передвигающимся амёбоидным способом. Лёгкие и тяжёлые цепи актина и миозина накапливаются в интернализующихся клетках в клеточном кортексе, противолежащем кадгериновому межклеточному контакту, но не в поглощающих клетках, как это происходит при фагоцитозе. Кроме того, при фагоцитозе поглощаются только мёртвые или умирающие клетки, а при энтозе — живые[11]. Наряду с актином и миозином в кортексе поглощаемой клетки локализуются RhoA, ROCKI и ROCKII, а также RhoA-зависимый актинполимеризующий формин mDia1. Образование зоны актин-миозинового сокращения, которая поляризуется так, чтобы обеспечивать проникновение одной клетки в другую, обеспечивается при помощи белков: активирующего белка Rho-ГТФаз (GAP), который привлекается к кадгериновым межклеточным контактам и подавляет RhoA, а также фактора обмена гуаниновых нуклеотидов Rho (GEF), который привлекается к дистальному концу кортекса поглощаемой клетки и активирует RhoA[12].
Клетки, поглощённые путём энтоза, как правило, умирают, хотя некоторые делятся в вакуолях внутри клеток-хозяев или даже покидают клетку-хозяина без повреждений и способными к размножению. Тот факт, что поглощённая путём энтоза клетка может покинуть клетку-хозяина, свидетельствует о том, что в поглощаемых клетках до поглощения не запускается программа смерти. Действительно, клетки, подвергающиеся энтозу, не экспонируют на своей мембране фосфатидилсерин, функционирующий как сигнал «съешь меня» и необходимый для фагоцитоза[12]. Поглощаемая клетка находится внутри энтотической вакуоли клетки-хозяина. В конце концов энтотические вакуоли сливаются с лизосомами, и клетка, находящаяся внутри вакуоли, разрушается[13].
Гибель клетки при энтозе происходит в отсутствие активации каспазы-3, свойственной апоптозу, и погибающие клетки не демонстрируют признаков апоптоза, таких как фрагментация ДНК. Гибель таких клеток не предотвращается при введении ингибитора апоптоза Bcl-2. Гибель поглощённых энтозом клеток некоторыми свойствами напоминает аутофагию, так как они накапливают аутофагосомы. Впрочем, в этом случае аутофагия нужна для выживания клетки, так как при блокировке аутофагии клетка погибает апоптозом. Поскольку поглощённые клетки изолированы от питательных веществ и факторов роста, присутствующих в среде, скорее всего, голодание индуцирует аутофагию. Поскольку аутофагия направлена на выживание поглощённой клетки, кажется странным тот факт, что для гибели поглощённой клетки нужны белки аутофагии. В этом случае белки аутофагии не участвуют в формировании аутофагосом, а направляют липидирование лёгкой цепи 3 (LC3) ассоциированного с микротрубочками белка 1А и его прикрепление к одномембранным вакуолям. Липидирование LC3 на энтотических вакуолях способствует слиянию с лизосомами, так что гибель клетки при энтозе происходит из-за зависимого от аутофагии лизосомного разрушения живых поглощённых клеток[12].
Образовавшиеся после разрушения питательные вещества переносятся в цитозоль, а вакуоли подвергаются процессингу, механизм которого ещё до конца не понятен. Показано, что энтотические вакуоли делятся на более мелкие вакуоли, при этом содержимое энтотических вакуолей распределяется по лизосомной сети. Деление вакуолей регулируется серин/треонин-киназой mTORC1, которая локализуется в мембране вакуоли, окружающей поглощённую клетку[13].
Механизм энтоза, спровоцированного митозом, несколько отличается от описанного выше. При митозе энтоз происходит, когда делящаяся эпителиальная клетка внедряется в соседнюю с формированием структуры «клетка внутри клетки». Впоследствии внутренняя клетка погибает и разрушается. Процесс может начаться на любой стадии митоза. При митозе энтоз может протекать в самых разных конфигурациях: две дочерние клетки интернализуются внутри одной или разных клеток, иногда даже образуются структуры типа матрёшки: одна из дочерних клеток интернализуется внутри другой, а она, в свою очередь, внедряется в соседнюю клетку. При митозе энтоз запускается в отсутствие белка Cdc42, при ингибировании Rap1, остановке митоза на стадии прометафазы (например, после обработки нокодазолом) и у некоторых раковых клеток. Ключевые белки, вовлечённые в митотический энтоз, — Rho, ROCK, миозин, актин, α-катенин, то есть в отличие от энтоза, вызванного утратой связи с матриксом, в митотическом энтозе не задействованы кадгерины[2].
Энтоз может запускаться в условиях нехватки глюкозы. Так, у прикреплённых клеток рака груди линии MCF7 в отсутствие глюкозы в течение 72 часов более 30 % клеток интернализовались в своих соседях. Поглощённая клетка служит дополнительным источником питательных веществ для клетки-хозяина, способствуя, таким образом, её выживанию и даже пролиферации. Нехватка глюкозы, кроме того, значительно ускоряет энтоз. Если в обычных условиях последний длится около суток, то при недостатке глюкозы около 60 % клеток оказываются мертвы через 5 часов после поглощения[2]. Стоит отметить, что в условиях недостатка глюкозы клетки погибают также по пути апоптоза и некроза[14]. Было показано, что при недостатке глюкозы поглощающими клетками («победителями») становятся клетки, у которых уровень экспрессии АМФ-активируемой протеинкиназы (AMPK) ниже. AMPK выступает в роли сенсора энергетического состояния клетки. Таким образом, можно заключить, что энтоз представляет собой форму комплексного ответа популяции клеток на глюкозное голодание[15].
Биологические функции
У мышей при имплантации зародыша эпителиальные клетки матки должны быть удалены, чтобы зародыш мог прикрепиться к её стенке. Показано, что эпителиальные клетки подвергаются энтозу в трофэктодерме бластоцисты, чтобы позволить произойти имплантации. У животных с подавленной работой киназ ROCK имплантация не происходила. Иммуноокрашивание показало, что на границе поглощаемой и поглощающей клеток находится β-катенин. Гибель интернализованных эпителиальных клеток происходила в отсутствие каспазы 3, поэтому смерть происходила не по программе апоптоза. Таким образом, имеются первые доказательства того, что энтоз задействован в эмбриональном развитии млекопитающих[12][11]. В 2017 году было показано, что в условиях спячки у дальневосточной черепахи (Pelodiscus sinensis) клетки Сертоли поглощают сперматозоиды по пути энтоза[16].
Было показано, что энтоз увеличивает геномную нестабильность поглощающей клетки (клетки-хозяина), поскольку нарушает её цитокинез и, следовательно, приводит к анеуплоидии дочерних клеток. Многие энтотические клетки рака груди имеют несколько ядер, поэтому, возможно, частая встречаемость многоядерных клеток в линиях раковых клеток связана с высокой частотой энтоза. Клетка-хозяин часто не может нормально поделиться после энтоза из-за нарушений в формировании сократительного кольца[6].
Клиническое значение
Клеточные структуры, напоминающие энтоз, описаны для многих видов рака, таких как рак молочной железы, толстой кишки, печени, поджелудочной железы, почек[17] и других органов. Показано, что энтоз может стимулироваться онкогенами, например, Kras[8]. При выращивании в культуре клинических образцов рака молочной железы показано, что на границе поглощающей и поглощаемой клеток находится белок β-катенин, а в кортексе поглощаемой клетки — лёгкие цепи миозина, фосфорилированные ROCK I/II. Иногда в опухолях обнаруживаются сложные структуры из трёх клеток, расположенных одна в другой. Энтоз может противостоять росту опухолевых клеток. Подавление энтоза путём нокдауна белков аутофагии усиливает злокачественный рост опухолевых клеток. Поскольку энтоз запускается при откреплении клетки от межклеточного матрикса, что также запускает процесс гибели по типу аноикиса, который подавлен при метастазировании, можно заключить, что энтоз выступает в качестве механизма, замедляющего рост численности опухолевых клеток. Однако было показано, что энтоз может вызывать анеуплоидию, которая способствует развитию опухоли[17]. При энтозе деление поглощающих клеток нарушается из-за блокировки образования борозды, что часто приводит к нарушению цитокинеза. Было также показано, что поглощающие клетки забирают питательные вещества от поглощённых клеток, подвергающихся энтозу, что способствует росту опухолевых клеток в условиях голодания[12].
Противоположные действия энтоза на рост клеток могут способствовать соревнованию двух соседних клеток, при котором поглощаемая клетка погибает и «проигрывает», а поглощающая клетка получает питательные вещества и «выигрывает». В основе такого соревнования лежит разница в активности пути Rho и актомиозинового комплекса соседних клеток[12]. В соревновании нормальной клетки и клетки, претерпевшей злокачественное перерождение, всегда «выигрывает» последняя, подтверждая тем самым, что активация онкогенов связана с способностью к «выигрышу». Кроме того, показано, что к «выигрышу» более способны клетки, которые легче нормальных подвергаются механической деформации, что также характерно для опухолевых клеток[8]. Механическая деформация контролируется RhoA и актомиозиновым комплексом. Поглощение клетки регулируется Kras и Rac, которые влияют на миозин[18].
Примечания
- Salvesen G. S. Dying from within: granzyme B converts entosis to emperitosis. (англ.) // Cell death and differentiation. — 2014. — Vol. 21, no. 1. — P. 3—4. — doi:10.1038/cdd.2013.157. — PMID 24317271.
- Durgan J., Florey O. Cancer cell cannibalism: Multiple triggers emerge for entosis. (англ.) // Biochimica Et Biophysica Acta. — 2018. — June (vol. 1865, no. 6). — P. 831—841. — doi:10.1016/j.bbamcr.2018.03.004. — PMID 29548938.
- Walton E. L. Can cannibalizing cancer cells challenge classic cell death classification? (англ.) // Biomedical Journal. — 2017. — June (vol. 40, no. 3). — P. 129—132. — doi:10.1016/j.bj.2017.06.001. — PMID 28651733.
- Overholtzer M., Mailleux A. A., Mouneimne G., Normand G., Schnitt S. J., King R. W., Cibas E. S., Brugge J. S. A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion. (англ.) // Cell. — 2007. — Vol. 131, no. 5. — P. 966—979. — doi:10.1016/j.cell.2007.10.040. — PMID 18045538.
- Garanina A. S., Kisurina-Evgenieva O. P., Erokhina M. V., Smirnova E. A., Factor V. M., Onishchenko G. E. Consecutive entosis stages in human substrate-dependent cultured cells. (англ.) // Scientific Reports. — 2017. — 2 October (vol. 7, no. 1). — P. 12555—12555. — doi:10.1038/s41598-017-12867-6. — PMID 28970591.
- Martins I., Raza S. Q., Voisin L., Dakhli H., Law F., De Jong D., Allouch A., Thoreau M., Brenner C., Deutsch E., Perfettini J. L. Entosis: The emerging face of non-cell-autonomous type IV programmed death. (англ.) // Biomedical Journal. — 2017. — June (vol. 40, no. 3). — P. 133—140. — doi:10.1016/j.bj.2017.05.001. — PMID 28651734.
- Cano C. E., Sandí M. J., Hamidi T., Calvo E. L., Turrini O., Bartholin L., Loncle C., Secq V., Garcia S., Lomberk G., Kroemer G., Urrutia R., Iovanna J. L. Homotypic cell cannibalism, a cell-death process regulated by the nuclear protein 1, opposes to metastasis in pancreatic cancer. (англ.) // EMBO molecular medicine. — 2012. — Vol. 4, no. 9. — P. 964—979. — doi:10.1002/emmm.201201255. — PMID 22821859.
- Kroemer G., Perfettini J. L. Entosis, a key player in cancer cell competition. (англ.) // Cell research. — 2014. — Vol. 24, no. 11. — P. 1280—1281. — doi:10.1038/cr.2014.133. — PMID 25342563.
- Gupta N., Jadhav K., Shah V. Emperipolesis, entosis and cell cannibalism: Demystifying the cloud. (англ.) // Journal Of Oral And Maxillofacial Pathology : JOMFP. — 2017. — January (vol. 21, no. 1). — P. 92—98. — doi:10.4103/0973-029X.203763. — PMID 28479694.
- Hamann J. C., Surcel A., Chen R., Teragawa C., Albeck J. G., Robinson D. N., Overholtzer M. Entosis Is Induced by Glucose Starvation. (англ.) // Cell Reports. — 2017. — 5 July (vol. 20, no. 1). — P. 201—210. — doi:10.1016/j.celrep.2017.06.037. — PMID 28683313.
- Li Y., Sun X., Dey S. K. Entosis allows timely elimination of the luminal epithelial barrier for embryo implantation. (англ.) // Cell reports. — 2015. — Vol. 11, no. 3. — P. 358—365. — doi:10.1016/j.celrep.2015.03.035. — PMID 25865893.
- Krishna S., Overholtzer M. Mechanisms and consequences of entosis. (англ.) // Cellular and molecular life sciences : CMLS. — 2016. — Vol. 73, no. 11—12. — P. 2379—2386. — doi:10.1007/s00018-016-2207-0. — PMID 27048820.
- Krajcovic M., Krishna S., Akkari L., Joyce J. A., Overholtzer M. mTOR regulates phagosome and entotic vacuole fission. (англ.) // Molecular biology of the cell. — 2013. — Vol. 24, no. 23. — P. 3736—3745. — doi:10.1091/mbc.E13-07-0408. — PMID 24088573.
- Püschel F., Muñoz-Pinedo C. In the Hunger Games, the Winner Takes Everything. (англ.) // Trends In Biochemical Sciences. — 2017. — October (vol. 42, no. 10). — P. 763—764. — doi:10.1016/j.tibs.2017.08.004. — PMID 28869131.
- Hamann J. C., Overholtzer M. Entosis enables a population response to starvation. (англ.) // Oncotarget. — 2017. — 29 August (vol. 8, no. 35). — P. 57934—57935. — PMID 28938526.
- Ahmed N., Yang P., Huang Y., Chen H., Liu T., Wang L., Nabi F., Liu Y., Chen Q. Entosis Acts as a Novel Way within Sertoli Cells to Eliminate Spermatozoa in Seminiferous Tubule. (англ.) // Frontiers In Physiology. — 2017. — Vol. 8. — P. 361—361. — doi:10.3389/fphys.2017.00361. — PMID 28611685.
- Kong Y., Liang Y., Wang J. Foci of Entotic Nuclei in Different Grades of Noninherited Renal Cell Cancers. (англ.) // IUBMB life. — 2015. — Vol. 67, no. 2. — P. 139—144. — doi:10.1002/iub.1354. — PMID 25855323.
- Sun Q., Luo T., Ren Y., Florey O., Shirasawa S., Sasazuki T., Robinson D. N., Overholtzer M. Competition between human cells by entosis. (англ.) // Cell research. — 2014. — Vol. 24, no. 11. — P. 1299—1310. — doi:10.1038/cr.2014.138. — PMID 25342560.
Типы клеточной гибели | |
|---|---|
| Непрограммируемые | |
| Программируемые | |
