Фосфоенолпируваткарбоксилаза
Фосфоенолпируваткарбоксилаза (ФЕП-карбоксилаза) представляет собой фермент из семейства карбоксилаз, который встречается у растений и некоторых бактерий. Он катализирует присоединение бикарбоната (НСО3−) к фосфоенолпирувату (ФЕП) с образованием четырёх углеродного соединения оксалоацетата и неорганического фосфата[1]:
- ФЕП + НСО3− → оксалоацетат + Фн
| Фосфоенолпируваткарбоксилаза | |
|---|---|
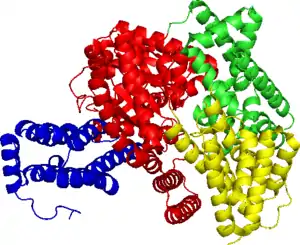 Мономер ФЕП-карбоксилазы из Flaveria trinervia. | |
| Идентификаторы | |
| Шифр КФ | 4.1.1.31 |
| Номер CAS | 9067-77-0 |
| Базы ферментов | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Gene Ontology | AmiGO • EGO |
| Поиск | |
| PMC | статьи |
| PubMed | статьи |
| NCBI | NCBI proteins |
| CAS | 9067-77-0 |
Это первая реакция фиксации углерода у СAM (англ. crassulacean acid metabolism) и С4-растений, а также одна из анаплеротических реакций цикла трикарбоновых кислот у бактерий и растений. Структура фермента, а также его двухступенчатый каталитический механизм достаточно хорошо изучены. Активность ФЕП-карбоксилазы находится под жёстким контролем и регулируется как путём фосфорилирования, так и аллостерически.
Структура
ФЕП-карбоксилаза присутствует в растениях и некоторых видах бактерий, но отсутствует у грибов или животных (включая человека)[2]. Нуклеотидная последовательность гена этого фермента отличается между организмами, но всегда сохраняются активный центр фермента и аллостерические сайты связывания, необходимые для его регуляции. Консервативной остаётся и его третичная структура[3].
Кристаллическая структура ФЕП-карбоксилазы была определена для нескольких организмов, в том числе для Zea maysа (кукурузы) и Escherichia coli[3]. Фермент существует в виде димера димеров: две идентичные субъединицы связываются, образуя димер через солевые мостики между аргинином (R438 — точное положение может меняться в зависимости от происхождения гена) и глутаминовой кислотой (E433)[4]. Этот димер в свою очередь связывается с другим димером и вместе они образуют комплекс из четырёх субъединиц. Каждая субъединица состоит в основном из альфа-спиралей (65 %)[1], имеет массу 106 кДа[5] и состоит из приблизительно 966 аминокислот[6].
Активный центр фермента охарактеризован не полностью. Он включает в себя консервативные остатки аспарагиновой (D564) и глутаминовой кислот (E566), которые через свои карбоксильные группы нековалентно связывают двухвалентный катион[1]. В зависимости от организма это может быть ион магния, марганца или кобальта[1][2], его роль заключается в координации молекулы фосфоенолпирувата (ФЕП) и промежуточных продуктов реакции. Остаток гистидина (H138) в активном центре, как считается, служит, для переноса протона при катализе[1][4].
Механизм катализа
Механизм катализа ФЕП-карбоксилазы довольно хорошо изучен. Реакция образования оксалоацетата является высокоэкзотермической и потому необратима; изменение энергии Гиббса для этого процесса (△G°’) составляет −30 кДж/моль[1]. Субстрат и кофактор связываются в следующем порядке: ион металла (Cо2+, Mg2+ или Mn2+), ФЕП и гидрокарбонат (НСО3−)[1][2]. Реакция протекает в два основных этапа, как описано ниже и показано на схеме:
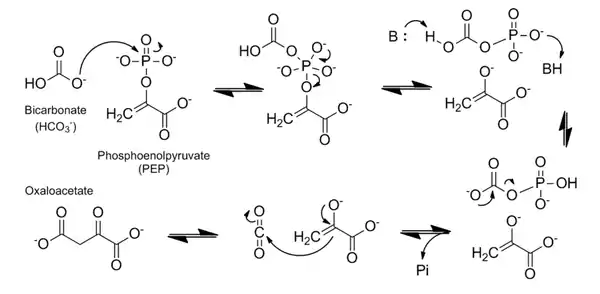
1. Гидрокарбонат выступает как нуклеофил и атакует фосфатную группу ФЕП. Это приводит к расщеплению ФЕП на карбоксифосфат (активированную форму CO2) и очень реакционноспособную енольную форму пирувата.
2. На карбоксифосфат переносится протон. В этом процессе участвует остаток гистидина (H138), который сначала отщепляет протон от карбоксильной группы, а затем, как кислота, переносит его на фосфат[1]. После этого карбоксифосфат разлагается на углекислый газ и неорганический фосфат с выделением энергии, что делает реакцию необратимой. Наконец, углекислый газ подвергается атаке енолата в результате чего и образуется оксалоацетат[1][2][7].
Двухвалентный катион координирует енолат и углекислый газ в ходе реакции; молекула СО2 теряется только в 3 % случаев[2]. Активный центр фермента является гидрофобным и не пропускает воду, поскольку карбоксифосфат довольно легко гидролизуется[1].
Биологическая функция
ФЕП-карбоксилаза выполняет три главные функции:
- Первичная фиксация углекислоты в форме бикарбоната в клетках мезофилла листа при С4-фотосинтез,
- Первичная фиксация углекислоты при CAM-фотосинтезе
- И поддержание уровня интермедиатов в цикле трикарбоновых кислот.
Основной механизм ассимиляции углекислоты растениями происходит через фермент рибулозо-1,5-дифосфаткарбоксилазу-оксигеназу (Рубиско), которая присоединяет СО2 к рибулозо-1,5-дифосфату (пяти углеродный сахар) с образованием двух молекул 3-фосфоглицерата. Однако при высоких температурах и низкой концентрации СО2 Рубиско вместо углекислого газа присоединяет кислород, что приводит к образованию метаболически инертного продукта гликолата, который перерабатываться в процессе фотодыхания. Чтобы предотвратить это бесполезный процесс, растения могут увеличивать локальную концентрацию CО2 с помощью С4-фотосинтеза[3][8]. ФЕП-карбоксилаза играет ключевую роль в фиксации СО2 в виде бикарбонат-аниона, соединяя его с ФЕП для создания оксалоацетата в ткани мезофилла. Затем оксалоацетат превращается обратно в пируват (через малат), чтобы высвободить СО2 в более глубоком слое обкладки проводящего пучка, где углекислый газ фиксируется Рубиско в цикле Кальвина. Пируват преобразуется обратно в ФЕП в клетках мезофилла, и цикл начинается снова. Таким образом происходит активное концентрирование СО2[2][9][10].
Вторая важная и очень похожая функция ФЕП-карбоксилазы — участие в CAM-фотосинтезе. Этот метаболический путь распространен у растений, живущих в засушливых местообитаниях. Растения не могут позволить открытые устьица в течение дня, чтобы поглощать CО2, так как при этом теряется слишком много воды в процессе транспирации. Вместо этого устьица открываются ночью, когда испарение воды минимально, CО2 Связывается путём фиксации ФЕП-карбоксилазой в форме оксалоацетата. Оксалоацетат затем превращается в малат с помощью фермента малатдегидрогеназы и депонируется в вакуоли, а затем используется в течение дня, когда световые реакции генерируют достаточно энергии (главным образом в виде АТФ) и восстановительных эквивалентов (НАДФН) для работы цикла Кальвина[2][3][10].
Третья функция ФЕП-карбоксилазы не связана с фотосинтезом. Аналогично пируваткарбоксилазе, ФЕП-карбоксилаза пополняет пул оксалоацетата в цикле трикарбоновых кислот. Образовавшийся в ходе гликолиза ФЕП преобразуется в пируват, который превращается в ацетил-КоА и поступает в ЦТК, где взаимодействует с оксалоацетатом, образуя цитрат. Чтобы увеличить поток вещества через цикл часть ФЕП преобразуется в оксалоацетат с помощью ФЕП-карбоксилазы, восполняя оксалоацетат, который откачивается из цикла для синтеза биомолекул клетки. ЦТК представляет собой центральный путь обмена веществ, поэтому увеличение потока проходящего через него вещества важно для биосинтеза многих молекул, например аминокислот[11].
Регуляция
.png.webp)
ФЕП-карбоксилаза регулируется двумя путями: через фосфорилирование и аллостерически. На рисунке сбоку показана схема механизма регуляции.
Фосфорилирование киназой фосфоенолпируваткарбоксилазы активирует фермент, а фосфатаза ФЕП-карбоксилазы снижает его активность. Как киназа так и фосфатаза регулируются на уровне транскрипции. Существует также мнение, что малат обеспечивает обратную связь в этом процессе, снижая уровень экспресси киназы и повышая экспрессию фосфатазы[12]. Оксалоацетат у CAM и С4-организмов превращается в малат, высокие концентрации которого активируют экспрессию фосфатазы, которая дефосфорилирует и деактивирует ФЕП-карбоксилазу, что ведёт к снижению накопления оксалоацетата, а следовательно и малата[1][12].
Основными аллостерическими ингибиторами ФЕП-карбоксилазы являются карбоновые кислоты такие как малат и аспартат[5][12]. Поскольку малат образуется на следующем шаге САМ и С4 циклов, сразу после того как ФЕП-карбоксилаза катализирует конденсацию СО2 и ФЕП в оксалоацетат, то образуется обратная связь. И аспартат, и оксалоацетат легко превращаются друг в друга по механизму трансаминирования; таким образом, высокие концентрации аспартата путём обратной связи ингибируют ФЕП-карбоксилазу.
Основные аллостерические активаторы ФЕП-карбоксилазы — это ацетил-КоА (только у бактерий)[13], фруктозо-1,6-дифосфат[1][13] и триозофосфаты (только у растений)[14]. Эти молекулы являются индикаторами активного гликолиза и сигнализируют о необходимости производства оксалоацетата, чтобы усилить поток вещества через цикл трикарбоновых кислот. Кроме того, увеличение гликолиза означает усиленное снабжение ФЕП и, следовательно, больше акцептора для фиксации СО2 и транспорта его в цикл Калвина. Примечательно также, что отрицательные эффектор аспартат конкурирует с положительным эффектором ацетил-КоА, что предполагает наличие у них общего сайта связывания[15].
Исследования показали, что энергетические эквиваленты, такие как АМФ, АДФ и АТФ, не оказывает существенного влияния на ФЕП-карбоксилазу[16].
Величина аллостерического влияния этих разных молекул на активность ФЕП-карбоксилазы зависит от конкретного организма[17].
Примечания
- Kai, Yasushi; Matsumura, Hiroyoshi; Izui, Katsura. Phosphoenolpyruvate carboxylase: three-dimensional structure and molecular mechanisms (англ.) // Archives of Biochemistry and Biophysics : journal. — Elsevier, 2003. — Vol. 414, no. 2. — P. 170—179. — ISSN 0003-9861. — doi:10.1016/S0003-9861(03)00170-X. — PMID 12781768.
- Chollet, Raymond; Vidal, Jean; O'Leary, Marion H. PHOSPHOENOLPYRUVATE CARBOXYLASE: A Ubiquitous, Highly Regulated Enzyme in Plants (англ.) // Annual Review of Plant Biology : journal. — 1996. — Vol. 47, no. 1. — P. 273—298. — ISSN 1040-2519. — doi:10.1146/annurev.arplant.47.1.273.
- Paulus, Judith Katharina; Schlieper, Daniel; Groth, Georg. Greater efficiency of photosynthetic carbon fixation due to single amino-acid substitution (англ.) // Nature Communications : journal. — Nature Publishing Group, 2013. — Vol. 4, no. 2. — P. 1518. — ISSN 2041-1723. — doi:10.1038/ncomms2504. — PMID 23443546.
- Kai, Y.; Matsumura, H.; Inoue, T.; Terada, K.; Nagara, Y.; Yoshinaga, T.; Kihara, A.; Tsumura, K.; Izui, K. Three-dimensional structure of phosphoenolpyruvate carboxylase: A proposed mechanism for allosteric inhibition (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — Vol. 96, no. 3. — P. 823—828. — ISSN 0027-8424. — doi:10.1073/pnas.96.3.823.
- Gonzalez, Daniel H.; Iglesias, Alberto A.; Andreo, Carlos S. Active-site-directed inhibition of phosphoenolpyruvate carboxylase from maize leaves by bromopyruvate (англ.) // Archives of Biochemistry and Biophysics : journal. — Elsevier, 1986. — Vol. 245, no. 1. — P. 179—186. — ISSN 0003-9861. — doi:10.1016/0003-9861(86)90203-1. — PMID 3947097.
- RCSB PDB — 3ZGE: Greater efficiency of photosynthetic carbon fixation due to single amino acid substitution Structure Summary Page
- Fujita, Nobuyuki; Izui, Katsura; Nishino, Tokuzo; Katsuki, Hirohiko. Reaction mechanism of phosphoenolpyruvate carboxylase. Bicarbonate-dependent dephosphorylation of phosphoenol-.alpha.-ketobutyrate (англ.) // Biochemistry : journal. — 1984. — Vol. 23, no. 8. — P. 1774—1779. — ISSN 0006-2960. — doi:10.1021/bi00303a029. — PMID 6326809.
- Leegood, Richard C. A welcome diversion from photorespiration (англ.) // Nature Biotechnology. — Nature Publishing Group, 2007. — Vol. 25, no. 5. — P. 539—540. — ISSN 1087-0156. — doi:10.1038/nbt0507-539. — PMID 17483837.
- Hatch, Marshall D. C(4) photosynthesis: discovery and resolution (неопр.) // Photosynthesis Research. — 2002. — Т. 73, № 1/3. — С. 251—256. — ISSN 0166-8595. — doi:10.1023/A:1020471718805. — PMID 16245128.
- Keeley, Jon E.; Rundel, Philip W. Evolution of CAM and C4Carbon‐Concentrating Mechanisms (англ.) // International Journal of Plant Sciences : journal. — 2003. — Vol. 164, no. S3. — P. S55—S77. — ISSN 1058-5893. — doi:10.1086/374192.
- Cousins, A. B.; Baroli, I.; Badger, M. R.; Ivakov, A.; Lea, P. J.; Leegood, R. C.; von Caemmerer, S. The Role of Phosphoenolpyruvate Carboxylase during C4 Photosynthetic Isotope Exchange and Stomatal Conductance (англ.) // Plant Physiology : journal. — 2007. — Vol. 145, no. 3. — P. 1006—1017. — ISSN 0032-0889. — doi:10.1104/pp.107.103390. — PMID 17827274.
- Nimmo, Hugh G. The regulation of phosphoenolpyruvate carboxylase in CAM plants (англ.) // Trends : journal. — 2000. — Vol. 5, no. 2. — P. 75—80. — ISSN 1360-1385. — doi:10.1016/S1360-1385(99)01543-5. — PMID 10664617.
- Morikawa M., Izui K., Taguchi M., Katsuki H. Regulation of Escherichia coli phosphoenolpyruvate carboxylase by multiple effectors in vivo. Estimation of the activities in the cells grown on various compounds (англ.) // Journal of Biochemistry : journal. — 1980. — February (vol. 87, no. 2). — P. 441—449. — PMID 6987214.
- José A. Monreal, Fionn McLoughlin, Cristina Echevarría, Sofía García-Mauriño and Christa Testerink. Phosphoenolpyruvate Carboxylase from C4 Leaves Is Selectively Targeted for Inhibition by Anionic Phospholipids (англ.) // Plants Physiology : journal. — February 2010. — Vol. 152, no. 2. — P. 634—638. — doi:10.1104/pp.109.150326. — PMID 20007442.
- Smith, Thomas E. Escherichia coli phosphoenolpyruvate carboxylase: Competitive regulation by acetyl-coenzyme A and aspartate (англ.) // Archives of Biochemistry and Biophysics : journal. — Elsevier, 1970. — Vol. 137, no. 2. — P. 512—522. — ISSN 0003-9861. — doi:10.1016/0003-9861(70)90469-8. — PMID 4909168.
- Coombs, J.; Maw, Susan L.; Baldry, C. W. Metabolic regulation in C4 photosynthesis: PEP-carboxylase and energy charge (англ.) // Planta : journal. — 1974. — Vol. 117, no. 4. — P. 279—292. — ISSN 0032-0935. — doi:10.1007/BF00388023. — PMID 24458459.
- Schuller, K. A.; Plaxton, W. C.; Turpin, D. H. Regulation of Phosphoenolpyruvate Carboxylase from the Green Alga Selenastrum minutum: Properties Associated with Replenishment of Tricarboxylic Acid Cycle Intermediates during Ammonium Assimilation (англ.) // Plant Physiology : journal. — 1990. — Vol. 93, no. 4. — P. 1303—1311. — ISSN 0032-0889. — doi:10.1104/pp.93.4.1303. — PMID 16667617.