Раса
Ра́са — система популяций человека, характеризующаяся сходством по комплексу физических или социальных признаков в соответствии с воспринятием их обществом[1]. Изначально термин имел языковые и национальные коннотации, а к XVII веку стал использоваться по отношению к признакам, имеющим внешнее фенотипическое проявление и сформировавшихся в определённом географическом регионе[2]. Такие признаки могли появиться как результат адаптации к различным условиям среды, происходившей в течение многих поколений.

В результате генетических исследований конца XX — начала XXI веков многие генетики пришли к выводу о невозможности точного выделения рас, так как различия и сходство рас по внешним признакам не всегда совпадают с генетическими различиями и сходством. В связи с этим и другими соображениями многие учёные считают термин «раса» в отношении человека не имеющим генетической основы[3].
С точки зрения современной западной науки расы являются социальными конструктами и идентичностями, которые присваиваются людям в зависимости от правил, принятых в обществе[4]. Будучи частично основанными на физическом сходстве людей внутри групп, расы тем не менее не несут физического или биологического смысла[1][5][6].
Разделение людей на расы менялось на протяжении человеческой истории, оно часто включало в себя народные таксономии, определяющие присущие людям черты характера по внешним признакам[7]. Современная наука считает такой биологический эссенциализм устаревшим[8] и в целом скептична к расовым объяснениям физических и поведенческих отличий между группам людей[9][10][11][12][13].
Несмотря на широкий научный консенсус о некорректности эссенциалистской и типологической концепций расы[14][15][16][17][18][19], учёные в разных странах продолжают разными способами пытаться изучать расы[20]. После Второй мировой войны термин «раса» был дискредетирован из-за связи расовой теории с преступлениями нацизма. Взамен для исследования групп людей используются другие термины, такие так популяция, народ, этническая группа, сообщество[21][22].
Разные школы антропологов выделяли от трёх до семи основных рас и десятки малых антропологических типов, среди основных — европеоидная, монголоидная, негроидная и американоидная.
Этимология
Слово «раса» в русском языке известно с середины XIX века, являясь заимствованием из фр. race или нем. Rasse, которые в свою очередь восходят к исп. raza или итал. razza. Дальнейшая этимология не вполне ясна: существуют версии о происхождении слова от лат. generatio («рождение, способность к размножению»), лат. ratio («род», «порода», «разновидность») или араб. ra’s («голова», «происхождение», «начало»)[23][24].
Расоведение
Расове́дение — раздел науки антропологии, изучающий человеческие расы[25].
Расоведение изучает классификацию рас, историю их формирования и такие факторы их возникновения, как селективные процессы, изоляция, смешение и миграции, влияние климатических условий и вообще географической среды на расовые признаки.
Изучение рас с уклоном в «научный расизм», утверждающий о якобы существующем превосходстве одних рас над другими, получило особенное распространение в национал-социалистической Германии и др. западноевропейских странах, а также ранее в США (Ку-клукс-клан), где служило обоснованием институционализированного расизма, шовинизма и антисемитизма.
Велик вклад в расоведение советских учёных (таких, как В. В. Бунак, В. П. Алексеев и др.).
Иногда расоведение отождествляют с этнической антропологией. Однако последняя относится, строго говоря, только к изучению расового состава отдельных этносов, то есть племён, народов, наций, и происхождению этих общностей.
В той части расовых исследований, которые направлены на изучение этногенеза, антропология ведёт исследования совместно с языкознанием, историей, археологией. При изучении движущих сил расообразования антропология вступает в тесный контакт с генетикой, физиологией, зоогеографией, климатологией, общей теорией видообразования. Изучение рас в антропологии имеет значение для решения многих проблем. Оно важно для использования антропологического материала как исторического источника, освещения проблем систематики, главным образом малых систематических единиц, познания закономерностей популяционной генетики, уточнения некоторых вопросов медицинской географии.
Концепция расы в современной науке
Научная польза понятия «раса» подвергалась сомнению ещё в 1930-е годы[3]. В середине XX века возникла концепция нереальности рас как определённых физическими признаками внутривидовых подразделений человечества. Одним из первых её сформулировал бельгийский учёный Ж. Йерно, назвавший расу не фактом, а понятием.
В 1964 году антропологи из разных стран, в том числе СССР, разработали для ЮНЕСКО документ, в котором утверждалось, что так как географические вариации признаков, которые используются в классификациях, сложны и не имеют резких разрывов, человечество не может разделяться на строго разграниченные категории с четкими границами[3].
Считается доказанным, что каждый «расовый признак» определяется несколькими генами, ареал каждого из которых не совпадает. Популяции отличаются не генотипами, а частотой тех или иных аллелей, которые не влияют на основные функции человека[3].
В современной западной антропологии был сформирован вывод об отсутствии рас как определённых физическими признаками внутривидовых подразделений и существовании только «клинальной изменчивости». Естественнонаучное содержание понятия «раса» связывается с признанным устаревшим типологическим подходом, а внимание концентрируется на изучении биологической вариативности и её причин[3].
В советской антропологии близкие по сути выводы формулировались ещё в 1930-е годы и поддерживаются в настоящее время в российской науке. Так, В. В. Бунак отмечал условность классификаций и отсутствие «чистых рас», а Г. И. Петров называл расы «исторически слагающимися категориями». Антрополог А. А. Зубов, хотя и считал неверным отказ от понятия «раса», отмечал, что все человеческие расы составляют вместе единый подвид вида Homo sapiens[3].
Этнолог и археолог В. А. Шнирельман, отмечает, что отказ западными специалистами от понятия «раса» достаточно хорошо обоснован научно, хотя и был также мотивирован политическими соображениями, а также что рассуждающий о расах «учёный не может оставаться в рамках чистой биологии». При этом он отмечает, что западные специалисты, отказываясь от понятия «раса», имеют в виду понимание расы, свойственное расистам или обывателям[3].
Однако вне зависимости от признания расы биологической реальностью или социальной конструкцией, все ученые соглашаются с тем, что человечество представляет собой единый биологический вид, а границы между популяциями не являются жёсткими и связаны с географическими, социальными и культурными, а не биологическими факторами[3].
Деление на расы
Концепции
Существует множество мнений о том, сколько рас может быть выделено внутри вида Homo sapiens. Существующие точки зрения варьируют от гипотезы двух основных расовых стволов до гипотезы 15 самостоятельных рас. Между этими крайними точками зрения лежит широкий спектр гипотез, постулирующий от 3 до 5 расовых стволов[26].
Согласно Советскому энциклопедическому словарю, существует около 30 человеческих рас (расово-антропологических типов), объединённых в три группы рас, которые именуются «большими расами»: негроидная, европеоидная и монголоидная. Сами расы (малые расы) делятся на субрасы, и нет единого мнения относительно принадлежности тех или иных субрас к тем или иным расам (малым расам). Кроме того, разные антропологические школы используют разные названия для одних и тех же рас.
Типологическая концепция расы
Типологическая концепция расы исторически появляется первой. Согласно типологическому подходу, описав черты конкретного человека, можно чётко отнести его к той или иной расе: выделяются расовые типы, и каждый индивид оценивается по степени приближения к тому или иному «чистому» типу. Например, ширина губ и носа больше определённой величины в сочетании с низким головным указателем, большим выступанием лица вперёд, курчавоволосостью и кожей темнее некого типового стандарта расценивается как свидетельство принадлежности к негроидной расе. По такой схеме можно даже определять расовую принадлежность конкретного человека в процентах. Сложность типологической концепции заключается в выделении «чистых» типов, отчётливо отличающихся один от другого. В зависимости от количества таких типов и признаков, определённых как расовые, будет меняться и расовое определение человека. Более того, последовательное строгое применение типологического принципа приводит к тому, что родные братья могут быть отнесены к разным расам[27].
Как отмечал советский антрополог В. П. Алексеев, типологическая концепция расы «всё больше приобретает характер анахронизма и отходит в историю антропологической науки»[28].
Ряд гипотез в рамках типологической концепции (например, существование экваториальной расы, объединяющей негроидную и австралоидную) были опровергнуты современными генетическими исследованиями.
Популяционная концепция расы
В современном российском расоведении господствует популяционная концепция расы. Согласно ей, расы — это не дискретные общности, а группы популяций, между которыми есть плавные переходы[29]. При этом подходе расовый тип не может определяться у индивидуума[30].
В США отход от типологической концепции расы к популяционно-генетической датируется 1950 г. В СССР основы популяционной концепции расы были сформулированы ещё в 1938 г. В. В. Бунаком. В дальнейшем концепция была развита В. П. Алексеевым[31].
Историческая концепция расы

Согласно концепции, сформулированной В. В. Бунаком в 1938 г. расы не являются стабильными, а представляют собой меняющиеся во времени категории[32]. Историческая концепция не противоречит популяционной, а лишь дополняет её, поскольку объясняет, почему между расами не существует жёстких границ[33].
Примеры классификаций
А. А. Зубов, опираясь на данные одонтологических признаков, разделял человечество на западный и восточный надрасовые стволы. При этом к западному стволу он отнёс европеоидную и негроидную расы, а к восточному — монголоидную, американоидную и австралоидную.
В. В. Бунак выделял 4 расовых ствола на основе представлений о древности формирования: тропический, южный, западный и восточный. Внутри тропического ствола выделялись африканская и океанийская расовые ветви, внутри южного — континентальная и древнеиндонезнийская, внутри западного — кушитская, средиземная, средиземноморская и европейская, внутри восточного — уральская, сибирская, южноазиатская и американская[25].
Часто выделяемые расы
Европеоидная раса
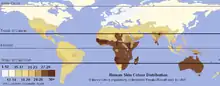
Ареал европеоидной расы включает раннюю область расселения — Европу, Ближний Восток, Северную Африку, а также Среднюю и Центральную Азию, Индостан, и области, в которых они расселились в сравнительно недавнее время — Северную и Южную Америку, Австралию и Южную Африку. Характерные признаки европеоидов включают ортогнатный профиль лица (выступающий вперёд в горизонтальной плоскости). Волосы прямые или волнистые, как правило мягкие (в частности, у северных групп). Глаза имеют широкий разрез, хотя глазная щель бывает небольшой, нос обычно средне или сильно выступающий с высоким переносьем, губы тонкие или утолщенные, сильный или средний рост волос на лице и теле. Широкие кисти рук и стопы. Цвет кожи, волос и глаз разнообразный: от очень светлых оттенков у северных групп до очень тёмных у южных. На границах ареала образует плавные переходы к уральской, южносибирской, монголоидной, негроидной, эфиопской и дравидийской расам.
Древнее человечество, представленное ранними тропическими популяциями Homo (Homo habilis, Homo rudolfensis, Homo ergaster и др.), было, вероятно, тёмнопигментированным. Биологический смысл светлой кожи — улучшение синтеза витамина D, участвующего в процессах роста и усвоения кальция костями, что предотвращает развитие у детей рахита в условиях плохой освещённости. Этот признак возникал многократно у различных групп, начиная с неандертальцев, и является приспособительным к условиям арктического климата, до конца последнего оледенения доминировавшего на большей части территории Европы вплоть до средиземноморского побережья. Светлая окраска радужной оболочки глаза и волос отчасти связана с пигментацией кожи, отчасти — является случайно возникшим нейтральным признаком. Появление на севере Европы аномально светлопигментированых популяций («голубоглазых блондинов») — следствие неоднократного проявления «эффекта основателя» и «эффекта бутылочного горлышка» в условиях крайней изоляции. Светлая пигментация является рецессивным признаком — даже при гетерозиготном наличии аллеля синтеза меланина она легко переходит в тёмную[34].
Данные генетики показали, что охотники-собиратели, жившие 8500 лет назад на территории современных Испании, Люксембурга и Венгрии, обладали тёмной кожей, у них не фиксируются гены SLC24A5 и SLC45A2, отвечающие за депигментацию. У охотников-собирателей севера Европы, живших 7700 лет назад, фиксируются эти гены, а также HERC2/OCA2, связанный с голубыми глазами и, вероятно, светлой кожей и волосами. Но окончательное распространение гена светлой кожи SLC24A5 по Центральной и Южной Европе связано с миграцией неолитических земледельцев с Ближнего Востока[35].
Негроидная раса

Согласно новейшим генетическим исследованиям, негроиды представляют собой нескольких рано обособившихся человеческих линий, причём бушмены и пигмеи в отношении генетического родства противостоят не только всем остальным африканским линиям, но и всем прочим группам людей вообще[36]. Среди негроидов можно выделить как минимум три резко обособленные друг от друга расы — негрскую, центральноафриканскую (пигмейскую) и южноафриканскую (бушменскую, капоидную).
Негрская раса распространена на большей территории Африки южнее Сахары. Характерные признаки: очень тёмная кожа, волосы курчавые, нос широкий, с уплощённым переносьем, большое межглазное пространство, губы толстые, долихокефалия, лицо низкое, прогнатное[37].
Негрилльская (или пигмейская, или центральноафриканская) раса распространена в экваториальных дождевых лесах Центральной Африки. Характерные признаки: очень низкий рост, сильный рост бороды, усов и волос на теле, широкий и короткий нос с плоским переносьем и часто выпуклой спинкой, сравнительно тонкие губы[38].

Южноафриканская (бушменская, капоидная) раса распространена в засушливых областях Южной Африки. Характерные признаки: низкий рост, плоское лицо, маленькая нижняя челюсть, благодаря чему лицо приобретает подтреугольную форму, нос узкий, переносье плоское, развит эпикантус, сравнительно светлая желтовато-бурая кожа, спирально-завитые волосы, стеатопигия у женщин, слабый рост бороды и усов. Многие признаки напоминают монголоидные.
Эфиопская раса
Раса с переходными чертами, имеющая признаки негроидной расы и европеоидные черты.
Монголоидная раса
Распространена в основном в Азии. Иногда объединяется с американоидной расой в большую азиатско-американскую расу. Характерные черты: цвет кожи варьирует от светлых оттенков до тёмно-смуглых, уплощённое лицо с выдающимися скулами, чаще высокое, высокие орбиты, ортогнатизм и мезогнатизм, рост бороды, усов и волос на теле слабый, разрез глаз узкий, часто встречается эпикантус, небольшие ресницы; степень выступания носа варьирует, но чаще небольшая, переносье обычно вогнутое, толщина губ от малой до утолщённой в Юго-Восточной Азии, ширина рта небольшая. На границах ареала имеет плавные переходы в европеоидную, дравидийскую, веддоидную, меланезийскую, полинезийскую, уральскую и южносибирскую расы.
Американоидная раса
Американоидная раса — раса, распространена в Северной и Южной Америке. Эпикантус сравнительно редок у взрослых, хотя довольно част у детей. Характерные черты: высокое крупное лицо с широкой нижней челюстью, мезогнатизм, средний или большой нос, иногда «орлиной» формы, с высоким переносьем, широкий рот, глазная щель шире, чем у азиатских монголоидов, уплощённость лица меньше, чем у монголоидов, но больше, чем у других рас, рост бороды и усов слабый. Часто объединяется с собственно монголоидной расой в азиатско-американскую. Заселение Америки происходило в основном через район Берингова пролива. Многие ископаемые останки и признаки отдельных современных популяций могут свидетельствовать о древнейшем проникновении в Америку людей из Полинезии и Меланезии.
Веддо-австралоидная раса

К австралоидной подгруппе этой большой расы относят коренное население Австралии. Характерные признаки: массивность долихокранного черепа при небольшой массивности остального скелета, сильные надбровные дуги, крупные прогнатные челюсти, большие зубы, короткая шея, тёмная пигментация кожи, широкий нос с вогнутым переносьем, рост бороды и усов сильный. Наиболее близки к австралоидной расе веддоидная и меланезийская расы, с которыми иногда объединяется в австрало-веддоидную расу[39].
Веддоидная подгруппа распространена в Центральной и Южной Индии, на Шри-Ланке, в Юго-Восточной Азии и Индонезии. Характерные признаки: грацильное телосложение, укороченная шея, тёмная кожа, менее широкий нос, широкие губы, рост бороды и усов от среднего до сильного. Может быть описана как грацильный вариант австралоидной расы, с которой иногда объединяется в австрало-веддоидную расу[40]. От австралоидной расы отличается также меньшим ростом бороды и усов, меньшим прогнатизмом, менее широким носом, меньшим ростом. Имеются многочисленные переходы к монголоидной, дравидийской и европеоидной расам[40].
Вопрос формирования расовых признаков
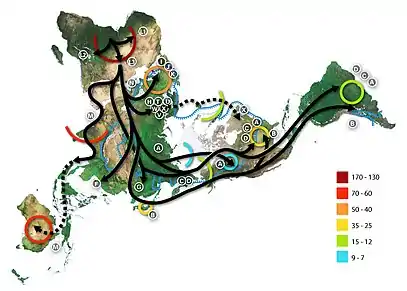
Вероятнее всего, расы в современном смысле слова возникли лишь после окончания последнего оледенения и начала неолитической революции благодаря появлению первых земледельческих культур, которые сумели за небольшое в историческом масштабе время резко увеличить свою численность и за счёт этого обеспечить своему комплексу расовых признаков доминирование на большой территории, сформировав современные «большие расы». Многие исследователи говорят об отсутствии рас в верхнем палеолите, или «верхнепалеолитическом полиморфизме» человечества. По мнению антрополога Станислава Дробышевского, дело не в том, что расовые черты верхнепалеолитических людей были ещё «недостаточно дифференцированы» или «не до конца сформировались», а в том, что ни одна из групп их не могла добиться долговременного преимущества перед другими — то есть, имело место не большее единообразие «недифференцированного» человечества, а, напротив, его большая мозаичность (полиморфность), из которой впоследствии выкристаллизовались современные расовые типы. До этого же момента малочисленные популяции палеолитических охотников-собирателей, жившие зачастую в условиях частичной или полной изоляции друг от друга, за счёт генетико-автоматических процессов накапливали такое количество локальных особенностей, что среди них невозможно выделить чётко очерченные расовые группы, обладающие устойчивым комплексом признаков[41][42][43][44].
В настоящее время отрицается приспособительный характер многих расовых признаков — более вероятно, что популяциям, являвшимися их носителям, просто «повезло» в эволюционном отношении, что позволило случайному комплексу характерных для них внешних признаков закрепиться и получить широкое распространение[45]. Вероятно, особую роль в этом процессе имело появление первых земледельческих культур, которые сумели резко увеличить свою численность, оттеснив группы, бывшие носителями иных расовых типов, на окраины ареала, и тем самым по сути сформировав так называемые «большие расы». За пределами же ареалов древних земледельцев такой нивелировки расовых признаков на основе численного преобладания носителей определённого типа не произошло, следствием чего стало сохранение большого разнообразия расовых признаков у американских индейцев, австралийских аборигенов, меланезийцев, койсаноидных жителей Южной Африки и других групп. Эти группы ни в коем случае не являются в эволюционном отношении «застойными» или более архаичными («протоморфными») по сравнению с «большими расами». Напротив — в больших группах, живущих в антропогенных ландшафтах, изменчивость признаков резко падает, появляется тенденция к их консервации, нарушаемая лишь метисацией на границах ареалов. Биологическая эволюция здесь в значительной степени замещается социальным и техническим развитием, хотя и не прекращается вообще. В то же время малочисленные, изолированные друг от друга популяции, испытывающие сильное воздействие отбора, отличаются повышенной пластичностью, быстро накапливая как приспособительные, так и случайные, нейтральные в эволюционном отношении, но заметно сказывающиеся на внешнем облике, признаки[41][42][43][46].
Так, «робустность» (массивность сложения) аборигенов Австралии является сравнительно недавним эволюционным приобретением, являющимся следствием биологической адаптации к тяжёлым условиям жизни, а вовсе не следствием их «протоморфности» (архаичности). Причём, судя по данным археологии, в сравнительно недавнее (историческое) время тенденцию к увеличению массивности у предков аборигенов сменила обратная — в сторону грацильности, вероятно, вследствие социального прогресса или изменения условий среды на более мягкие. В то же время австралийцы европейского происхождения не обнаруживают никаких признаков биологического приспособления к среде обитания (в том числе и в отдалённой перспективе), поскольку окружили себя «второй природой» — высокоразвитой техносферой, позволяющей существовать в условиях Австралии человеку, биологически сравнительно плохо к ним адаптированному. С эволюционной точки зрения белые австралийцы являются, пожалуй, более «протоморфными» и «архаичными», чем аборигены, которые сравнительно недавно (в эволюционном масштабе) приобрели целый ряд специализированных (то есть, прогрессивных с эволюционной точки зрения) черт.
Впрочем, роль техники всё же не стоит абсолютизировать — так, есть наблюдения, позволяющие судить о воздействии естественного отбора на популяцию современных людей, в совсем недавнее время (вторая половина XX века) участвовавших в освоении Крайнего Севера. На протяжении жизни одного поколения практически все неприспособленные к таким условиям поселенцы вернулись на Большую землю — остались лишь те, кто имел адаптивный в таких условиях тип, то есть, по особенностям телосложения и обмена веществ был приспособлен к экстремальным холодам (причём показательно, что по этим особенностям они напоминали местное аборигенное население). Если бы эта популяция людей продолжила размножаться эндогамно и подвергалась бы при этом жёсткому естественному отбору, как это обычно было при миграциях в доисторическое и раннеисторическое время, в ней всего за несколько поколений закрепился бы устойчивый набор признаков, соответствующих адаптации к экстремальному холоду[47].
Древние и реликтовые расы
Современная популяционная генетика допускает, что ныне существующие расы не исчерпывают всего исторического морфологического разнообразия людей современного типа, и что в древности существовали расы, которые либо исчезли бесследно, либо признаки которых были позднее размыты в результате ассимиляции носителями других рас[48]. В частности, уралист В. В. Напольских выдвинул гипотезу о существовании в прошлом палеоуральской расы, признаки которой в настоящее время размыты между урало-сибирскими европеоидами и западными монголоидами, однако не являются свойственными ни для европеоидов в целом, ни для монголоидов в целом[49].
Верхнепалеолитические типы
Антрополог С. В. Дробышевский указывает, что морфологическое разнообразие людей в верхнем палеолите было, возможно, даже более выраженным, чем в настоящее время, и что черепа людей того времени не могут быть чётко диагностированы с применением современной расовой классификации, географическая или временная принадлежность часто также явно не выражена.
В частности на основании европейских находок он описывает следующие выделяемые разными авторами доисторические расы или морфологические типы, часть из которых выделена на основе одного черепа:
- гримальдийская
- кроманьонская
- Барма-Гранде
- шанселядская
- оберкассельская
- брюннская
- брюнн-пржедмостская
- ориньякская
- солютрейская
На Ближнем Востоке того же времени отмечается преемственность пре-натуфийцев и натуфийцев. Они имеют протоевропеоидные черты, иногда с негроидной примесью. При этом натуфийцы имеют отличия от североафриканских групп Афалу и Тафоральт. Среди восточноафриканских находок выделялись «бушменский тип», «эфиопский тип», «негроидный тип», более массивный, чем современный. Верхнепалеолитические черепа из Китая, Юго-Восточной Азии и Индонезии часто не обнаруживают монголоидности, но сближаются с восточными экваториалами и чаще определяются как «прото-австралоиды» или «австрало-меланезийский тип». Многие находки региона никак не могут быть описаны с использованием современной расовой классификации и сочетают в себе черты как южных монголоидов, так и австралоидов, айнов, носителей культуры ёмон (или дзёмон), архаичных индейцев и других групп[50].
Метисация

В выделении рас как популяций, обладающих отличными от других признаками, важную роль играла географическая изоляция. Такая изоляция ещё несколько тысяч лет назад была обусловлена в основном огромными расстояниями и малой численностью людей. В результате увеличения численности или миграций популяции контактировали и происходило смешение рас или метисация. В результате появились смешанные антропологические типы или малые расы (южносибирская, полинезийская и другие)[51][52].
Все известные расы человечества могут давать плодовитое совместное потомство, даже наиболее изолированным популяциям, вроде американских индейцев или австралийских аборигенов, «не хватило» как минимум нескольких миллионов лет изоляции до возникновения биологической несовместимости с другими группами[53].
В результате смешанных браков (и метисации вообще) появляются индивиды со смешанными расовыми признаками, а в зонах контакта между ареалами различных рас — целые контактные расы, обладающие таковыми на популяционном уровне. Для них в некоторых культурах принята специальная терминология. Так, мулаты — результат совмещения негроидной и европеоидной расы, метисы — европеоидной и американоидной/монголоидной, а самбо — негроидной и американоидной. В настоящее время значительная часть населения Земли является в той или иной степени метисной, например, большая часть жителей Центральной и Южной Америки. С другой стороны, сравнительная устойчивость комплекса расовых признаков подобных крупных групп метисного происхождения позволяет рассматривать их как самостоятельные малые расы, находящиеся в процессе формирования.
Было проведено множество исследований, которые показали отсутствие вредных для потомства с медицинской точки зрения последствий межрасовой метисации. Это является следствием сравнительно недавнего расхождения рас и постоянных контактов между ними на протяжении истории[54].
Расизм
Идеи о превосходстве одних рас (или расовых типов) над другими получили развитие в XIX — середине XX века и были признаны ложными с научной точки зрения[55]. Отрицательную роль сыграло их распространение в истории человечества: от геноцида коренного населения Нового Света и до Холокоста времён Второй мировой войны.
См. также
Примечания
- Barnshaw, John. Race // Encyclopedia of Race, Ethnicity, and Society, Volume 1. — SAGE Publications, 2008. — P. 1091–3. — ISBN 978-1-45-226586-5.
- Рогинский Я. Я., Левин М. Г. Антропология: Учебник для студентов ун-тов. — З-е изд., перераб.и доп. — М.: Высшая школа, 1978. — С. 341.
- Шнирельман В. А. «Порог толерантности»: Идеология и практика нового расизма. — М.: Новое литературное обозрение, 2011. — Т. 1. Глава 5. — С. 92—104.
- Gannon, Megan Race Is a Social Construct, Scientists Argue (англ.). Scientific American (5 February 2016). Дата обращения: 8 сентября 2020.
- Smedley, Audrey; Takezawa, Yasuko I. & Wade, Peter, Race: Human, Encyclopædia Britannica, Encyclopædia Britannica Inc., <http://www.britannica.com/topic/race-human>. Проверено 22 августа 2017.
- Yudell, M.; Roberts, D.; DeSalle, R.; Tishkoff, S. (2016-02-05). “Taking race out of human genetics”. Science [англ.]. 351 (6273): 564—565. Bibcode:2016Sci...351..564Y. DOI:10.1126/science.aac4951. ISSN 0036-8075. PMID 26912690.
- См.:
- Montagu, Ashley (1962). “The Concept of Race”. American Ethnography Quasimonthly. Дата обращения 26 January 2009.
- Bamshad, Michael; Olson, Steve E (10 November 2003). “Does Race Exist?” (PDF). Scientific American. 289 (6): 78—85. Bibcode:2003SciAm.289f..78B. DOI:10.1038/scientificamerican1203-78. PMID 14631734. Архивировано из оригинала (PDF) 14 June 2007. Используется устаревший параметр
|url-status=(справка)
- Sober, Elliott. Philosophy of biology. — 2nd. — Boulder, CO : Westview Press, 2000. — P. 148–151. — ISBN 978-0813391267.
- Lee et al., 2008: «We caution against making the naive leap to a genetic explanation for group differences in complex traits, especially for human behavioral traits such as IQ scores»
- American Anthropological Association Statement on 'Race'. AAAnet.org. American Anthropological Association (AAA) (17 May 1998). Дата обращения: 18 апреля 2009.: "For example, 'Evidence from the analysis of genetics (e.g., DNA) indicates that most physical variation, about 94 %, lies within so-called racial groups. Conventional geographic «racial» groupings differ from one another only in about 6 % of their genes. This means that there is greater variation within 'racial' groups than between them.Шаблон:'"
- Keita, S O Y; Kittles, R A; Royal, C D M; Bonney, G E; Furbert-Harris, P; Dunston, G M; Rotimi, C N (2004). “Conceptualizing human variation”. Nature Genetics. 36 (11s): S17—S20. DOI:10.1038/ng1455. PMID 15507998.
Modern human biological variation is not structured into phylogenetic subspecies ('races'), nor are the taxa of the standard anthropological 'racial' classifications breeding populations. The 'racial taxa' do not meet the phylogenetic criteria. 'Race' denotes socially constructed units as a function of the incorrect usage of the term.
- Harrison, Guy. Race and Reality. — Amherst : Prometheus Books, 2010. — «Race is a poor empirical description of the patterns of difference that we encounter within our species. The billions of humans alive today simply do not fit into neat and tidy biological boxes called races. Science has proven this conclusively. The concept of race (...) is not scientific and goes against what is known about our ever-changing and complex biological diversity.».
- Roberts, Dorothy. Fatal Invention. — London, New York : The New Press, 2011. — «The genetic differences that exist among populations are characterized by gradual changes across geographic regions, not sharp, categorical distinctions. Groups of people across the globe have varying frequencies of polymorphic genes, which are genes with any of several differing nucleotide sequences. There is no such thing as a set of genes that belongs exclusively to one group and not to another. The clinal, gradually changing nature of geographic genetic difference is complicated further by the migration and mixing that human groups have engaged in since prehistory. Human beings do not fit the zoological definition of race. A mountain of evidence assembled by historians, anthropologists, and biologists proves that race is not and cannot be a natural division of human beings.».
- Race Is Real, but not in the way Many People Think, Agustín Fuentes, Psychology Today.com, 9 April 2012
- The Royal Institution - panel discussion - What Science Tells us about Race and Racism.
- “Genetic variation, classification and 'race'”. Nature. Дата обращения 18 November 2014.
Ancestry, then, is a more subtle and complex description of an individual's genetic makeup than is race. This is in part a consequence of the continual mixing and migration of human populations throughout history. Because of this complex and interwoven history, many loci must be examined to derive even an approximate portrayal of individual ancestry.
- Michael White. Why Your Race Isn't Genetic. Pacific Standard. — «[O]ngoing contacts, plus the fact that we were a small, genetically homogeneous species to begin with, has resulted in relatively close genetic relationships, despite our worldwide presence. The DNA differences between humans increase with geographical distance, but boundaries between populations are, as geneticists Kenneth Weiss and Jeffrey Long put it, "multilayered, porous, ephemeral, and difficult to identify." Pure, geographically separated ancestral populations are an abstraction: "There is no reason to think that there ever were isolated, homogeneous parental populations at any time in our human past."». Дата обращения: 13 декабря 2014.
- The Genetic Ancestry of African Americans, Latinos, and European Americans across the United States. The American Journal of Human Genetics. — «The relationship between self-reported identity and genetic African ancestry, as well as the low numbers of self-reported African Americans with minor levels of African ancestry, provide insight into the complexity of genetic and social consequences of racial categorization, assortative mating, and the impact of notions of "race" on patterns of mating and self-identity in the US. Our results provide empirical support that, over recent centuries, many individuals with partial African and Native American ancestry have "passed" into the white community, with multiple lines of evidence establishing African and Native American ancestry in self-reported European Americans.». Дата обращения: 22 декабря 2014.
- Carl Zimmer. White? Black? A Murky Distinction Grows Still Murkier. «On average, the scientists found, people who identified as African-American had genes that were only 73.2 percent African. European genes accounted for 24 percent of their DNA, while .8 percent came from Native Americans. Latinos, on the other hand, had genes that were on average 65.1 percent European, 18 percent Native American, and 6.2 percent African. The researchers found that European-Americans had genomes that were on average 98.6 percent European, .19 percent African, and .18 Native American. These broad estimates masked wide variation among individuals.».
- Lieberman, L.; Kaszycka, K. A.; Martinez Fuentes, A. J.; Yablonsky, L.; Kirk, R. C.; Strkalj, G.; Wang, Q.; Sun, L. (December 2004). “The race concept in six regions: variation without consensus”. Coll Antropol. 28 (2): 907—21. PMID 15666627.
- Race2. Oxford Dictionaries. Oxford University Press. — «1. Each of the major division of humankind, having distinct physical characteristics [example elided]. 1.1. mass noun The fact or condition of belonging to a racial division or group; the qualities or characteristics associated with this. 1.2. A group of people sharing the same culture, history, language, etc.; an ethnic group [example elided].». Дата обращения: 5 октября 2012. Provides 8 definitions, from biological to literary; only the most pertinent have been quoted.
- Keita, S O Y; Kittles, R A; Royal, C D M; Bonney, G E; Furbert-Harris, P; Dunston, G M; Rotimi, C N (2004). “Conceptualizing human variation”. Nature Genetics. 36 (11s): S17—S20. DOI:10.1038/ng1455. PMID 15507998.
Many terms requiring definition for use describe demographic population groups better than the term 'race' because they invite examination of the criteria for classification.
- Раса / М. Фасмер. Этимологический словарь русского языка
- Раса / П. Я. Черных. Историко-этимологический словарь современного русского языка
- Глоссарий Архивировано 22 апреля 2015 года. // Богатенков Д. В., Дробышевский С. В. Антропология
- Алексеев В. П. Избранное. Т.2. Антропогеография. — М.: Наука, 2007. — С. 23. ISBN 978-5-02-035544-6
- Типологическая концепция // Богатенков Д. В., Дробышевский С. В. Антропология
- Становление человечества // Алексеев В. П. Избранное. Т.1. Антропогенез — М.: Наука, 2007. — С. 255. ISBN 978-5-02-035543-9
- Бунак В. В. Род Homo, его возникновение и последующая эволюция / Отв. редактор д.и.н. А. А. Зубов. — М.: Наука, 1980. — 329 с.
- Виктор Шнирельман «Цепной пес расы»: диванная расология как защитница «белого человека»
- Становление человечества // Алексеев В. П. Избранное. Т.1. Антропогенез — М.: Наука, 2007. — С. 255—258. ISBN 978-5-02-035543-9
- Дробышевский С. В. Введение. О сложности изучения расовой изменчивости // Антропогенез.ру
- «Бунак всегда прав»: о главном советском антропологе
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. А что у европеоидов?
- Ученые рассказали о недавней смене европейцами цвета кожи
- Александр Григорьевич Козинцев, д.и.н., главный научный сотрудник Музея антропологии и этнографии РАН им. Петра Великого (Кунсткамеры). Расовая классификация в свете новых генетических данных.
- Негрская раса / Пестряков А. П. // Нанонаука — Николай Кавасила. — М. : Большая российская энциклопедия, 2013. — С. 268—269. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 22). — ISBN 978-5-85270-358-3. Архивированная копия. Дата обращения: 2 июня 2019. Архивировано 30 декабря 2017 года. (Дата обращения: 30 декабря 2017)
- Центральноафриканская раса / Пестряков А. П. // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов. — М. : Большая российская энциклопедия, 2004—2017. Архивированная копия. Дата обращения: 2 июня 2019. Архивировано 30 декабря 2017 года. (Дата обращения: 30 декабря 2017)
- Алексеева, Богатенков, Дробышевский, 2004, с. 337.
- Алексеева, Богатенков, Дробышевский, 2004, с. 355.
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. Индейцы протоморфны ?
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. А что у европеоидов ?
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. О негрской и меланезийской расах.
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. О «верхнепалеолитическом полиморфизме».
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. Кау Свэмп: эректусами тут и не пахнет.
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. Социальные факторы антропогенеза.
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. У европейцев архаики будет побольше, чем у андаманцев….
- Дробышевский С. В.Формирование Homo sapiens. Когда возникли расы? // Антропогенез.ру
- Напольских В. В. Введение в историческую уралистику. — Ижевск: УИИЯЛ УрО РАН, 1997. — 268 с. — ISBN 5-7691-0671-9.
- Дробышевский С. В. О верхнепалеолитических расах // Антропогенез.ру
- Нестурх М. Ф. Человеческие расы. 1954. Происхождение человеческих рас. 4. Смешение (метисация)
- Алексеева Т. И. (ред.), Богатенков Д. В., Дробышевский С. В. Антропология: Учебно-методический комплекс . — 2004. 5.2.7, 5.2.8
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. Светлана Александровна Боринская, д. б. н., ведущий научный сотрудник лаборатории анализа генома Института общей генетики им. Н. И. Вавилова РАН. Для генетической несовместимости с европейцами индейцам не хватило нескольких миллионов лет.
- Станислав Владимирович Дробышевский, научный редактор АНТРОПОГЕНЕЗ.РУ, к.б.н., доцент кафедры антропологии биологического факультета МГУ им. Ломоносова. Светлана Александровна Боринская, д. б. н., ведущий научный сотрудник лаборатории анализа генома Института общей генетики им. Н. И. Вавилова РАН. О различиях между расами… и о телевизионных мошенниках
- Токарев С. А. История зарубежной этнографии. 1978
Литература
- Азимов А. Расы и народы = Races and People. — М.: Центрполиграф, 2005. — 206 с. — ISBN 5-9524-0399-9.
- Алексеев В. П. Новые споры о старых проблемах. — М.: Знание, 1991. — 64 с. — ISBN 5-07-001788-8.
- Алексеева Т. И. (ред.), Богатенков Д. В., Дробышевский С. В. Антропология: Учебно-методический комплекс. — М.: Институт дистантного образования Российского университета дружбы народов, 2004. — 459 с. Архивировано 10 сентября 2016 года. (Дата обращения: 8 мая 2018)
- Арутюнов С. А., Чебоксаров Н. Н. Раса, популяция и этнос / Доклад/ Советская социол. ассоц. Советский оргкомитет по подготовке VII Междунар. социол. конгресса. МСК VII. Варна. 1970; 165. — 1970. — 14 с.
- Бунак В. В. Род Homo, его возникновение и последующая эволюция. — М.: Наука, 1980.
- Кун К. С. Расы Европы = The Races of Europe. — М.: АСТ, Астрель, 2011. — 816 с.
- Нестурх М.Ф. Человеческие расы. — М.: Просвещение, 1965. — 110 с.
- Проблема расы в российской физической антропологии. — М.: Институт этнологии и антропологии РАН, 2002. — 96 с. — ISBN 5-201-00838-0.
- Хрисанфова Е. Н., Перевозчиков И. В. § 4. Географическая локализация и краткое описание основных антропологических типов // Антропология. Учебник.. — 2-е изд.. — М.: Изд-во МГУ, 1999. — 400 с.
- Раса / Чебоксаров Н. Н. // Проба — Ременсы. — М. : Советская энциклопедия, 1975. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 21).
- Раса // Еврейская энциклопедия Брокгауза и Ефрона. — СПб., 1908—1913.
- Проблема расы в современной науке // Шнирельман В. А. «Порог толерантности»: Идеология и практика нового расизма./ В 2-х томах. — М.: Новое литературное обозрение, 2011. — Т. 1 — 552 с.: ил. / Т.2 — 856 с.: ил. — ISBN 978-5-86793-874-1
- Вергелес М. О. Понятие «расы» в американской физической антропологии // Вестник Московского университета. Серия 23. Антропология. № 3, 2015.
- Тетушкин Е. Я. Существуют ли расы человека? // Человек. 2015. Т. № 1. С. 5-31.
- Lee, Sandra SJ; Mountain, Joanna; Koenig, Barbara; Altman, Russ (2008). “The ethics of characterizing difference: guiding principles on using racial categories in human genetics”. Genome Biol. 9 (7): 404. DOI:10.1186/gb-2008-9-7-404. PMC 2530857. PMID 18638359.
Ссылки
- Боринская С. А. О митохондриальной Еве и генетическом разнообразии современного человечества // Antropogenez.ru
- Богатенков Д. В., Дробышевский С. В. Антропология.
- Дробышевский С. В. Возникновение современных рас // Antropogenez.ru
- Антропологический справочник, содержащий краткие статьи об основных единицах расовой систематики (антропологических таксонах).
- Научно-просветительский форум «Учёные против мифов»
- Соколов А. Б. Расы — разные виды. Мифы об эволюции человека. ScienceVideoLab.
| В библиографических каталогах |
|---|
