Капикуа
Капикуа (англ. Capicua transcriptional repressor) — белок человека, способный подавлять экспрессию ряда генов (транскрипционный репрессор). Капикуа кодируется геном CIC, расположенным на длинном плече 19 хромосомы.
Структура
Белок капикуа эволюционно консервативен — отмечается схожесть его структуры у червя Caenorhabditis elegans и у человека. В организме выявляются две изоформы белка, короткая (CIC-S) и длинная (CIC-L), отличающиеся формой амино-конца. Белок содержит два эволюционно-консервативных домена: HMG-box (high-mobility group box) и домен C1 (C1 domain) – совместно эти домены распознают определенные октамерные последовательности нуклеотидов ДНК.[1] Также капикуа содержит сигнал ядерной локализации, позволяющий ему перемещаться в ядро клетки.

История
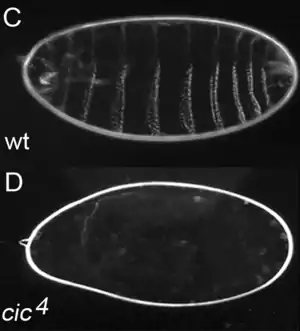
Ген CIC был впервые идентифицирован в 2000 году в организме мухи-дрозофилы.[1] Было показано, что он кодирует транскрипционный репрессор, участвующий в регулировке процессов эмбриогенеза. Ученые установили, что мутация, при которой у мушки не развиваются брюшные сегменты, но присутствуют сегменты головы и хвоста, поражает ген CIC, отсюда происходит и название гена — «capicua» по-каталански означает «голова-и-хвост».
Патологии
В 2017 году было показано, что мутации гена CIC могут вызывать расстройство под названием «аутосомно-доминантная умственная отсталость 45» (англ. mental retardation, autosomal dominant-45, MRD-45).[3]
Опухоли

При многих типах рака CIC является супрессором опухоли.[1] И напротив, в опухолях некоторых типов обнаруживаются мутации гена CIC. По состоянию на 2020 год, опухолевые мутации CIC наиболее часто отмечались при олигодендроглиоме.[1] Геномная транслокация, приводящая к образованию гибридного белка, состоящего частично из CIC, частично из DUX4, способна вызывать агрессивную разновидность «Юинг-подобной» саркомы.[5]
В то время как в своей нормальной форме капикуа подавляет экспрессию генов, образуемые с его участием гибридные белки, связанные с развитием опухолей, предположительно теряют эту функцию. Так, например, обстоит дело в случае с химерным белком CIC-DUX4, который является уже не репрессором, а активатором генов.[6]
Взаимодействия
- FOLR1 — капикуа оказывает влияние на экспрессию фолатного рецептора-альфа, а мутации CIC предположительно приводят к церебральной фолатной недостаточности[7]
- ATXN1 — капикуа образует комплекс с белком атаксин-1 («комплекс ATXN1-CIC»), важный для правильного формирования структур мозга.[3]
- DUX4 – отмечается образование химерных белков CIC-DUX4 в опухолях.[3]
- FOXO4 – отмечается образование химерных белков CIC-FOXO4 в опухолях.[6]
- NUTM1 – отмечается образование химерных белков CIC-NUTM1 в опухолях.[6]
- LEUTX – отмечается образование химерных белков CIC-LEUTX в опухолях.[6]
Литература
- «Регулировка и функции Капикуа у млекопитающих» (англ.) — обзор в открытом доступе по свободной лицензии, журнал Experimental & Molecular Medicine, 2020 год.[1]
- Making heads or tails – the emergence of capicua (CIC) as an important multifunctional tumour suppressor - обзор роли CIC в развитии раковых заболеваний, в открытом доступе, 2020 год.[6]
Ссылки
- De novo мутации гена CIC, вызывающие потерю функции гена, способствуют развитию церебральной фолатной недостаточности через снижение экспрессии FOLR1 — русский перевод исследования, осуществленного в 2020 году.[7]
- Mental retardation, autosomal-dominant, MRD-45 - "Умственная отсталость, аутосомно-доминантная, тип 45" — запись в генетическом каталоге OMIM.
Примечания
- Lee Y (2020). “Regulation and function of capicua in mammals”. Exp Mol Med. 52 (4): 531—537. DOI:10.1038/s12276-020-0411-3. PMC 7210929. PMID 32238859.
- Forés M, Simón-Carrasco L, Ajuria L, Samper N, González-Crespo S, Drosten M, Barbacid M, Jiménez G (March 2017). “A new mode of DNA binding distinguishes Capicua from other HMG-box factors and explains its mutation patterns in cancer”. PLoS Genetics. 13 (3): e1006622. DOI:10.1371/journal.pgen.1006622. PMC 5344332. PMID 28278156.
- Lu HC, Tan Q, Rousseaux MW, Wang W, Kim JY, Richman R, Wan YW, Yeh SY, Patel JM, Liu X, Lin T, Lee Y, Fryer JD, Han J, Chahrour M, Finnell RH, Lei Y, Zurita-Jimenez ME, Ahimaz P, Anyane-Yeboa K, Van Maldergem L, Lehalle D, Jean-Marcais N, Mosca-Boidron AL, Thevenon J, Cousin MA, Bro DE, Lanpher BC, Klee EW, Alexander N, Bainbridge MN, Orr HT, Sillitoe RV, Ljungberg MC, Liu Z, Schaaf CP, Zoghbi HY (April 2017). “Disruption of the ATXN1-CIC complex causes a spectrum of neurobehavioral phenotypes in mice and humans”. Nature Genetics. 49 (4): 527—536. DOI:10.1038/ng.3808. PMC 5374026. PMID 28288114.
- Nakai S, Yamada S, Outani H, Nakai T, Yasuda N, Mae H, Imura Y, Wakamatsu T, Tamiya H, Tanaka T, Hamada K, Tani A, Myoui A, Araki N, Ueda T, Yoshikawa H, Takenaka S, Naka N (November 2019). “Establishment of a novel human CIC-DUX4 sarcoma cell line, Kitra-SRS, with autocrine IGF-1R activation and metastatic potential to the lungs”. Scientific Reports. 9 (1): 15812. DOI:10.1038/s41598-019-52143-3. PMC 6825133. PMID 31676869.
- Antonescu CR, Owosho AA, Zhang L, Chen S, Deniz K, Huryn JM, Kao YC, Huang SC, Singer S, Tap W, Schaefer IM, Fletcher CD (July 2017). “Sarcomas With CIC-rearrangements Are a Distinct Pathologic Entity With Aggressive Outcome: A Clinicopathologic and Molecular Study of 115 Cases”. The American Journal of Surgical Pathology. 41 (7): 941—949. DOI:10.1097/PAS.0000000000000846. PMC 5468475. PMID 28346326.
- Wong D, Yip S (April 2020). “Making heads or tails - the emergence of capicua (CIC) as an important multifunctional tumour suppressor”. The Journal of Pathology. 250 (5): 532—540. DOI:10.1002/path.5400. PMID 32073140.
- Cao X, Wolf A, Kim SE, Cabrera RM, Wlodarczyk BJ, Zhu H; et al. (2020). “CIC de novo loss of function variants contribute to cerebral folate deficiency by downregulating FOLR1 expression”. J Med Genet. DOI:10.1136/jmedgenet-2020-106987. PMC 7895856. PMID 32820034.
