Зрение млекопитающих
Зре́ние млекопита́ющих — процесс восприятия млекопитающими видимого электромагнитного излучения, его анализа и формирования субъективных ощущений, на основании которых складывается представление животного о пространственной структуре внешнего мира. Отвечает за данный процесс у млекопитающих зрительная сенсорная система, основы которой сложились ещё на раннем этапе эволюции хордовых. Её периферическую часть образуют органы зрения (глаза), промежуточную (обеспечивающую передачу нервных импульсов) — зрительные нервы, а центральную — зрительные центры в коре головного мозга[1][2].
Распознавание визуальных стимулов у млекопитающих является результатом совместной работы органов зрения и головного мозга. При этом значительная часть зрительной информации обрабатывается уже на уровне рецепторов, что позволяет многократно сократить объём такой информации, поступающей к мозгу. Устранение избыточности количества информации неизбежно: если объём информации, поступающей на рецепторы зрительной системы, измеряется миллионами бит в секунду (у человека — порядка 1⋅107 бит/с), то возможности нервной системы по её обработке ограничены десятками бит в секунду[3].
Органы зрения у млекопитающих развиты, как правило, достаточно хорошо, хотя в их жизни они имеют меньшее значение, чем у птиц: обычно млекопитающие обращают мало внимания на неподвижные предметы, так что к стоящему без движения человеку даже столь осторожные звери, как лисица или заяц, могут подойти вплотную. Размеры глаз у млекопитающих относительно невелики; так, у человека масса глаз составляет 1 % от массы головы, в то время как у скворца достигает 15 %. Более крупные глаза имеют ночные звери (например, долгопят) и животные, обитающие в открытых ландшафтах. У лесных зверей зрение не столь острое, а у роющих подземных видов (кроты, гоферы, слепушонки, цокоры, златокроты) глаза в большей или меньшей мере редуцированы, в некоторых случаях (сумчатые кроты, слепыш, слепой крот) даже затянуты кожистой перепонкой[4][5].
Строение глаза
Как и у других позвоночных, глаз млекопитающего развивается из переднего мозгового пузыря и имеет округлую форму (глазное яблоко). Снаружи глазное яблоко защищено белковой фиброзной оболочкой, передняя часть которой прозрачна (роговица), а остальная — нет (склера). Следующий слой — сосудистая оболочка, спереди переходящая в радужную оболочку с отверстием в центре — зрачком[6]. Большая часть глазного яблока занята стекловидным телом, заполненным водянистой жидкостью. Поддержание формы глазного яблока обеспечивается за счёт жёсткой склеры и внутриглазного давления, создаваемого этой жидкостью. Эта водянистая жидкость регулярно обновляется: она выделяется в заднюю камеру глаза эпителиальными клетками цилиарного тела, откуда попадает в переднюю камеру через зрачок и далее попадает в венозную систему[7].
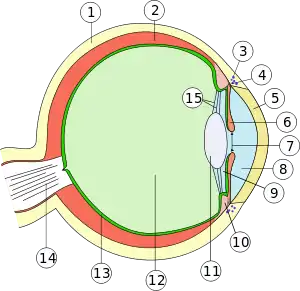
- 1 — склера,
- 2 — сосудистая оболочка,
- 3 — канал Шлемма,
- 4 — корень радужной оболочки,
- 5 — роговица,
- 6 — радужная оболочка,
- 7 — зрачок,
- 8 — передняя камера,
- 9 — задняя камера,
- 10 — цилиарное тело,
- 11 — хрусталик,
- 12 — стекловидное тело,
- 13 — сетчатка,
- 14 — зрительный нерв,
- 15 — цинновы связки.
Через зрачок отражённый от объектов свет проникает внутрь глаза. Количество пропускаемого света определяется диаметром зрачка, просвет которого автоматически регулируется мышцами радужной оболочки. Хрусталик, удерживаемый на месте цилиарным пояском, фокусирует прошедшие через зрачок лучи света на сетчатке — внутреннем слое оболочки глаза, содержащем фоторецепторы — светочувствительные нервные клетки[8]. Сетчатка состоит из нескольких слоёв (изнутри наружу): пигментный эпителий, фоторецепторы, горизонтальные клетки Кахаля, биполярные клетки, амакриновые клетки и ганглионарные клетки[7]. Подробнее о строении сетчатки см. ниже.
Окружающие хрусталик мышцы обеспечивают аккомодацию глаза. У млекопитающих для достижения высокой резкости изображения хрусталик при наблюдении близких объектов принимает выпуклую форму, при наблюдении удалённых — почти плоскую[8]. У пресмыкающихся и птиц аккомодация, в отличие от млекопитающих, включает не только изменение формы хрусталика, но и изменение расстояния между хрусталиком и сетчаткой. В целом способность глаза млекопитающего к аккомодации значительно уступает таковой у птиц: у человека она в детстве не превышает 13,5 дптр и заметно снижается с возрастом, а у птиц (особенно ныряющих) она может достигать 40—50 дптр. У мелких грызунов (полёвки, мыши) из-за незначительности обзора способность к аккомодации практически утрачена[4][9].
Роль защитных образований для глаз играют веки, снабжённые ресницами. У внутреннего угла глаза размещается гардерова железа, выделяющая жировой секрет (её нет у приматов), а в наружном углу — слёзная железа, выделения которой (слёзная жидкость) омывают глаз. Слёзная жидкость улучшает оптические свойства роговицы, сглаживая шероховатости её поверхности, а также защищает её от пересыхания и других неблагоприятных воздействий[7]. Эти железы наряду с веками и глазными мышцами относят к вспомогательному аппарату глаза[10][11].
Фоторецепторы
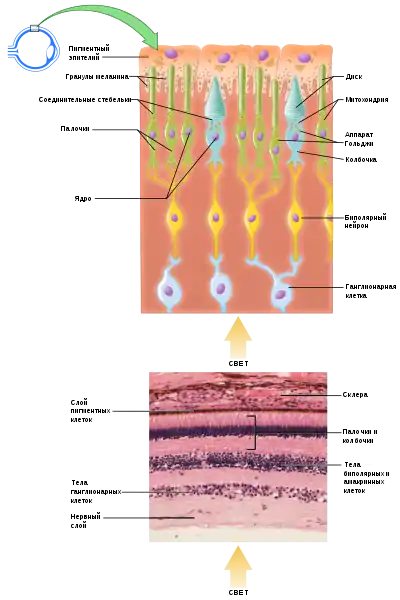
Среди фоторецепторов выделяют две основные разновидности — палочки и колбочки, причём палочки преобладают; так, у человека сетчатка содержит около 123 млн палочек и 7 млн колбочек[12]. Палочки отвечают за восприятие только интенсивности света и обеспечивают ночное зрение, а при дневном зрении ведущую роль играют колбочки, позволяя животным не только воспринимать свет, но и различать цвета[13]. Зрительные пигменты находятся в мембранных дисках колбочек и палочек[7].
Фоторецепторы содержат светочувствительные пигменты — опсины; это — трансмембранные белки, относящиеся к семейству GPCR[14], 7 α-спиралей опсина пронизывают мембрану[13]. С молекулой опсина связана молекула светоабсорбирующей молекулы — ретиналя (производное витамина А). Ретиналь и опсин в совокупности образуют зрительный пигмент палочек — родопсин. Ретиналь имеет угловой цис- и линейный транс-изомеры, причём при возбуждении светом цис-изомер переходит в транс-изомер. Такое изменение конфигурации ретиналя дестабилизирует и активирует связанный с ним опсин. После передачи возбуждения специальные ферменты возвращают ретиналь в исходное цис-состояние[13].
Возбуждение от активированного опсина передаётся на G-белок трансдуцин, который активирует фермент фосфодиэстеразу. Этот фермент отрывает от натриевого канала мембраны палочки цГМФ, гидролизуя его до ГМФ. В результате этого натриевые каналы палочки закрываются, и клетка гиперполяризуется (таким образом, рецепторный потенциал палочки запускается не деполяризацией, а гиперполяризацией). После этого в её синаптическом окончании, образующим синапс с расположенным после нейроном, не выделяется нейромедиатор глутамат (в темноте он, напротив, выделяется). В зависимости от типа глутаматного рецептора некоторые из граничащих с палочками нейронов в ответ на выделение или невыделение глутамата гиперполяризуются, другие — деполяризуются. Обычно с палочками контактируют биполярные клетки (одна — с несколькими палочками), но вместо них здесь могут находиться горизонтальные или амакриновые клетки. От них возбуждение передаётся ганглионарным клеткам, которые сообщают его зрительному нерву[15].
Колбочки используют такой же механизм передачи сигнала, как и палочки, но с некоторыми различиями. Существует три типа колбочек, содержащих соответственно три типа зрительных пигментов — фотопсинов, или йодопсинов: красных, зелёных и синих. Они образуются в результате связывания ретиналя с тремя различными типами опсинов. Хотя эти опсины несильно отличаются друг от друга, они реагируют на свет c разными длинами волн, при этом их спектры поглощения частично перекрываются. Перекрывание спектров обеспечивает ощущение других цветов; например, при возбуждении красных и зелёных колбочек глаз видит жёлтый или оранжевый цвет — в зависимости от того, какого типа колбочки более стимулированы[16]. В сетчатке имеются 3 типа ганглионарных клеток: М-клетки (α, или Y) — быстропроводящие, чувствительные к свету и особенно чувствительные к движению; P-клетки (β, или Х), которые обеспечивают высокое пространственное разрешение, стабильно реагируют на постоянный цвет и поэтому делают возможным анализ образов и цвета; W-клетки (или γ), которые регулируют диаметр зрачка и рефлекс быстрого скачкообразного движения глаз[17].
В отличие от пресмыкающихся и птиц, у млекопитающих колбочки не имеют светофильтров в виде цветных жировых капель[4]. Фоторецепторов нет на оптическом диске, и этот участок называется слепым пятном поля зрения[18].
Наружная светочувствительная часть палочек и колбочек регулярно обновляется: старые мембранные диски на их поверхности сбрасываются и заменяются новыми дисками из внутренней части, а отброшенные диски поглощаются фагоцитами[7].
Впрочем, у млекопитающих цветовое зрение развито слабее, чем у птиц с их четырёхкомпонентным зрением: у подавляющего большинства млекопитающих зрение — двухкомпонентное, а трёхкомпонентное цветовое зрение имеется только у высших приматов (узконосые и частично широконосые обезьяны)[19]. Так, европейская рыжая полёвка различает лишь красный и жёлтый цвета, а у опоссума, лесного хоря и некоторых других видов цветное зрение вообще не обнаружено[20]. В то же время некоторые сумчатые, рукокрылые и грызуны способны видеть в ультрафиолетовом диапазоне[21].
Установлено, что за цветное зрение у позвоночных отвечают 4 семейства генов, кодирующих опсины колбочек: SWS1, SWS2, Rh2, LWS. Все четыре семейства генов были выявлены у современных птиц, рыб и пресмыкающихся, у современных земноводных — лишь 3. У млекопитающих ситуация значительно сложнее. У современных однопроходных выявлены гены семейств SWS2 и LWS, а также нефункциональный ген из SWS1; у современных сумчатых имеются гены из SWS1 и LWS, а также, возможно, из Rh2. Современные плацентарные имеют гены опсинов только из семейств SWS1 и LWS[22].
В 1990-х гг. у млекопитающих был открыт третий тип фоторецепторов — светочувствительные ганглионарные клетки, содержащие меланопсин, обладающий очень слабой чувствительностью к свету. В восприятии зрительных образов эти рецепторы практически не задействованы, но они участвуют в управлении циркадными ритмами и в регуляции размера зрачка[14].

Часть света, достигшего сетчатки, проходит через неё и поглощается пигментным эпителием сетчатки. У многих млекопитающих (особенно у ночных) эта оболочка образует, однако, блестящий слой — тапетум (или «зеркальце»), образованный эластичными волокнами или эндотелиальными клетками[23]. Он отбрасывает лучи света обратно на сетчатку, снижая его потери[24]. Наличие тапетума обусловливает кажущее свечение глаз млекопитающих в почти полной темноте. Такое «свечение» глаз характерно для многих млекопитающих, особенно хищных, в том числе и некоторых приматов, но у человека встречается как атавизм[23].
Зрительные пути и обработка сигнала
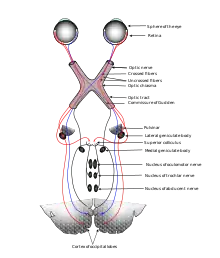
Итак, как отмечалось выше, аксоны ганглионарных клеток образуют зрительный нерв, который передаёт зрительную информацию от глаз в головной мозг. Каждый зрительный нерв располагается сзади от глазного яблока; его длина невелика, причём разные волокна зрительного нерва несут информацию от разных участков сетчатки. Существенно, что зрительные нервы от правого и левого глаз перекрещиваются, образуя частичный перекрёст зрительных нервов — зрительную хиазму, располагающуюся примерно в центре основания коры головного мозга. При этом нервные волокна, идущие от тех участков сетчатки, которые примыкают к носу, ведут в контралатеральное (противоположное) полушарие конечного мозга, а нервные волокна, отходящие от височных отделов сетчатки, ведут в ипсилатеральное полушарие; благодаря этому зрительная информация от каждого глаза поступает в оба полушария[25][26].
Помимо зрительного нерва, в промежуточную часть зрительной сенсорной системы входят подкорковые ганглии мозга и латеральные коленчатые тела. К числу подкорковых ганглиев мозга относят: предкрышечное поле среднего мозга, регулирующее диаметр зрачка; верхние бугры четверохолмия, участвующие в глазодвигательной функции; супрахиазматическое ядро гипоталамуса, выступающее в роли генератора циркадных ритмов. Латеральные коленчатые тела, лежащие в таламусе, являются важнейшими среди подкорковых зрительных центров и вносят существенный вклад в обработку зрительной информации. Большинство аксонов ганглионарных клеток приходят именно в латеральные коленчатые тела, и лишь меньшая часть этих аксонов проецируются на подкорковые ганглии мозга[27][28].
Из латеральных коленчатых тел сигнал поступает в центральную часть зрительной сенсорной системы — зрительную кору[28]. Зрительная кора подразделяется на первичную зрительную кору, расположенную в затылочной доле коры больших полушарий и иначе называемую стриарной корой, и экстрастриарную зрительную кору, состоящую из нескольких участков (зон), некоторые из которых располагаются также в височной и теменной долях. Первичная зрительная кора каждого полушария получает информацию от ипсилатерального наружного коленчатого тела, после чего информация передаётся по нескольким путям в различные зоны экстрастриарной зрительной коры. В результате зрительная информация по точкам проецируется на зрительную кору, где и происходит обработка характеристик изображения (цвета, формы, движения, глубины и др.), причём для целостного восприятия эти свойства должны быть интегрированы[26][27].
У многих млекопитающих хорошо развито бинокулярное зрение, основанное на формировании двух изображений, полученных каждым глазом, и их последующем сопоставлении. В ходе обмена информацией между обоими зрительными центрами два полученных изображения сливаются в одну трёхмерную картину[26].
Примечания
- Константинов, Наумов, Шаталова, 2012, с. 35, 336.
- Гистология, цитология и эмбриология, 2004, с. 340—341.
- Воротников С. А. . Информационные устройства робототехнических систем. — М.: Изд-во МГТУ им. Н. Э. Баумана, 2005. — 384 с. — ISBN 5-7038-2207-6. — С. 19—22.
- Дзержинский, Васильев, Малахов, 2014, с. 391.
- Константинов, Наумов, Шаталова, 2012, с. 336.
- Гистология, цитология и эмбриология, 2004, с. 341—344.
- Зильбернагль, Деспопулос, 2013, с. 356.
- Джадд Д., Вышецки Г. . Цвет в науке и технике. — М.: Мир, 1978. — 592 с. — С. 16—18.
- Константинов, Шаталова, 2004, с. 209, 273, 391.
- Гистология, цитология и эмбриология, 2004, с. 360—362.
- Payne A. P. The harderian gland: a tercentennial review // Journal of Anatomy. — 1994. — Vol. 185 (Pt 1). — P. 1—49. — PMID 7559104.
- Ткаченко, 2009, с. 389.
- Campbell, 2011, p. 1097.
- Terakita A. The opsins // Genome Biology. — 2005. — Vol. 6, № 3. — P. 213. — doi:10.1186/gb-2005-6-3-213. — PMID 15774036.
- Campbell, 2011, p. 1096—1099.
- Campbell, 2011, p. 1099, 1100.
- Зильбернагль, Деспопулос, 2013, с. 370.
- Зильбернагль, Деспопулос, 2013, с. 360.
- Bowmaker J. K. Evolution of colour vision in vertebrates // Eye (London, England). — 1998. — Vol. 12 (Pt 3b). — P. 541—547. — doi:10.1038/eye.1998.143. — PMID 9775215.
- Константинов, Шаталова, 2004, с. 391.
- Vaughan, Ryan, Czaplewski, 2011, p. 23.
- Jacobs G. H. Evolution of colour vision in mammals // Phil. Trans. R. Soc. B. — 2009. — Vol. 364, № 1531. — P. 2957—2967. — doi:10.1098/rstb.2009.0039.
- Тапетум — статья из Биологического энциклопедического словаря
- Locket N. A. The choroidal tapetum lucidum of Latimeria chalumnae // Proceedings of the Royal Society of London. Series B. — 1974. — Vol. 186, № 1084. — P. 281—290. — doi:10.1098/rspb.1974.0049. — PMID 4153107.
- Хомская Е. Д. . Нейропсихология. 4-е изд. — СПб.: Питер, 2011. — 496 с. — ISBN 978-5-459-00730-5. — С. 150.
- Campbell, 2011, p. 1099.
- Зильбернагль, Деспопулос, 2013, с. 370—371.
- Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 79, 116.
Литература
На русском языке
- Гистология, цитология и эмбриология. 6-е изд / Под ред. Ю. И. Афанасьева, С. Л. Кузнецова, H. А. Юриной. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. . Зоология позвоночных. 2-е изд. — М.: Издат. центр «Академия», 2014. — 464 с. — ISBN 978-5-4468-0459-7.
- Зильбернагль С., Деспопулос А. . Наглядная физиология. — М.: БИНОМ. Лаборатория знаний, 2013. — 408 с. — ISBN 978-5-94774-385-2.
- Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5.
- Константинов В. М., Шаталова С. П. . Зоология позвоночных. — М.: Гуманитарный издательский центр ВЛАДОС, 2004. — 527 с. — ISBN 5-691-01293-2.
- Лысов В. Ф., Ипполитова Т. В., Максимов В. И., Шевелёв Н. С. . Физиология и этология животных. 2-е изд. — М.: КолосС, 2012. — 605 с. — ISBN 978-5-9532-0826-0.
- Ткаченко Б. И., Брин В. Б., Захаров Ю. М., Недоспасов В. О., Пятин В. Ф. . Физиология человека. Compendium / Под ред. Б. И. Ткаченко. — М.: ГЭОТАР-Медиа, 2009. — 496 с. — ISBN 978-5-9704-0964-0.
На английском языке
- Campbell N. A., Reece J. B., Urry L. A. e. a. . Biology. 9th ed. — Benjamin Cummings, 2011. — 1263 p. — ISBN 978-0-321-55823-7.
- Vaughan T. A., Ryan J. M., Czaplewski N. J. . Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
Ссылки
- Чугунов, Антон. Зрительный родопсин — рецептор, реагирующий на свет. // Сайт Biomolecula.ru (10 марта 2007). Дата обращения: 18 марта 2018.
- Чугунов, Антон. Откуда пошло зрение. // Сайт Biomolecula.ru (8 ноября 2007). Дата обращения: 18 марта 2018.
- Лебедев, Виктор. Женский взгляд. // Сайт Biomolecula.ru (8 марта 2015). Дата обращения: 18 марта 2018.
- Чугунов, Антон. Оптогенетика + голография = прозрение?. // Сайт Biomolecula.ru (1 марта 2013). Дата обращения: 18 марта 2018.
- Бозрова, Светлана; Посух, Ольга. Когда ослепшие прозреют?. // Сайт Biomolecula.ru (11 апреля 2017). Дата обращения: 18 марта 2018.