Горизонтальные клетки
Горизонтальные клетки — слой ассоциативных нейронов сетчатки.
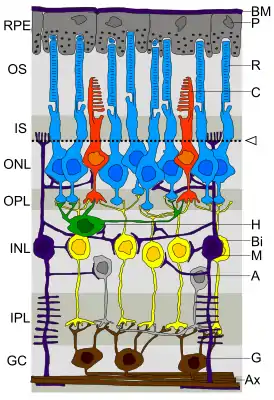
OS — наружный сегмент фоторецепторов
IS — внутренний сегмент фоторецепторов
ONL — внешний ядерный слой
OPL — внешний сплетениевидный слой
INL — внутренний ядерный слой
IPL — внутренний сплетениевидный слой
GC — ганглионарный слой
BM — мембрана Бруха
P — пигментные эпителиоциты
R — палочки
C — колбочки
Стрелка и пунктирная линия — внешняя пограничная мембрана
H — горизонтальные клетки
B — биполярные клетки
M — Клетки Мюллера
A — амакриновые клетки
G — ганглионарные клетки
AX — аксоны
Горизонтальные клетки располагаются в сетчатке сразу за фоторецепторами и отдают им большое количество дендритов, которые, переплетаясь, образуют сплошное густое кружево. Их аксоны могут тянуться на значительные расстояния и имеют сильно разветвленные концевые образования. Горизонтальные клетки соединены между собой электрическими синапсами и образуют функциональный синцитий. Возбуждение одной горизонтальной клетки распространяется на другие. В центральной ямке (лат. fovea) сетчатки приматов одна горизонтальная клетка соединяется примерно с 6-ю колбочками, а на периферии — с 30-40. Дендритное поле горизонтальной клетки на периферии сетчатки достигает диаметра 250 мкм. В то же время, размер рецептивных полей горизонтальных клеток может изменяться в пределах 300мкм ÷ 5мм[1], что значительно превышает протяженность дендритных разветвлений. За счет этого горизонтальные клетки могут формировать периферию рецептивных полей биполярных нейронов[2][3][4][5][6][7][8]. При освещении фоторецепторов горизонтальные клетки отвечают гиперполяризацией.
Фоторецепторы вместе с горизонтальными клетками и биполярами образуют так называемую триаду[4]. Триада состоит из пресинаптической мембраны фоторецептора, в которую заходит дендрит биполяра, а по бокам от него заходят отростки горизонтальных клеток. Значение и принципы функционирования триады пока остаются неизвестными. За счет таких связей между фоторецепторами, горизонтальными клетками и биполярами, центр рецептивного поля биполяров формируется прямым путём при передаче сигнала от фоторецепторов, периферия формируется за счет непрямого пути через горизонтальные клетки за счёт возбуждающих или тормозных синапсов.
Примечания
- Измайлов И. А., Соколов Е. Н., Чернорызов А. М. Психофизиология цветового зрения. — М.: Изд-во Московского университета, 1989. — 206 с.
- Школьник-Яррос Е. Г. , Калинина А. В. Нейроны сетчатки. — М.: Наука, 1986. — 208 с.
- Масланд Р. Функциональная организация сетчатки// В мире науки. — 1987, № 4 — с. 58-68.
- Бызов А. Л. Полищук Н. А. О механизме обратной связи от горизонтальных клеток к фоторецепторам: химическая или электрическая гипотеза?// Сенсорные системы. — 1987. Т1, № 4. — с.344-352.
- Бызов А. Л., Голубцов К. В. Модель нейрона-регулятора эффективности синаптической передачи// Биофизика. — 1978. — т.23 № 1. — с. 119—125.
- Шмидт Р. Тевс Г. Физиология человека. т.1. -М.: Мир, 1996. — 328 с.
- Dacey D. Lee B. Stafford D. Horizontal cells of the primate retina: cone specifity without spectral opponency // Science. — 1996. Vol. 271, № 5429. р. 656—659.
- Глезер В. Д. Зрение и мышление. — Л.: Наука, 1985, — 248 с.
| Фиброзная оболочка (внешняя) | |||||||
|---|---|---|---|---|---|---|---|
| Сосудистая оболочка (средняя) | |||||||
| Сетчатка (внутренняя оболочка) |
| ||||||
| Передний сегмент | |||||||
| Задний сегмент | |||||||
| Глазные мускулы | |||||||
| Зрачковые мышцы | |||||||
| Нервная система и другое |
| ||||||