Цианидиум
Циани́диум (лат. Cyanidium) — род примитивных ацидотермофильных красных водорослей. Описан один общепризнанный вид — цианидиум тепличный (Cyanidium caldarium).
| Цианидиум | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Biliphyta Отдел: Подотдел: Cyanidiophytina H.S.Yoon et al., 2006 Класс: Цианидиофициевые (Cyanidiophyceae Merola et al., 1981) Порядок: Цианидиевые Семейство: Цианидиевые Род: Цианидиум |
||||||||||
| Международное научное название | ||||||||||
| Cyanidium Geitler, 1933 | ||||||||||
| Единственный вид | ||||||||||
|
|
||||||||||
| ||||||||||
Один из самых примитивных эукариотических организмов. Одноклеточная водоросль, внешними морфологическими признаки сходная с зелёной водорослью хлореллой. Клетки обладают примитивным эукариотическим строением, а хлоропласты содержат пигменты, характерные для синезелёных водорослей, что позволяет предположить, что цианидиум представляет собой переходную стадию эволюции между цианобактериями и эукариотическими водорослями.
Широко распространённый в горячих кислотных источниках вид, всегда встречающийся вместе с видами близкого рода Galdieria, а нередко являющийся единственным фотосинтезирующим организмом в среде (фотосинтезирующие термофильные бактерии, например, Chloroflexus, при pH 2—3, оптимальном для цианидиума, существовать не могут).
Описание
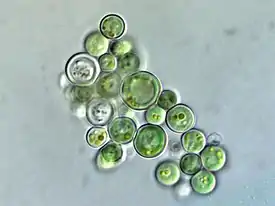
Обычно одноклеточная водоросль, редко несколько клеток собраны в скопления в общем гомогенном слизистом матриксе. Клетки шаровидные, (1,5)2—5(6) мкм в диаметре, голубовато-зелёные[1].
В клетке имеется единственный двумембранный хлоропласт чашевидной или шаровидной формы, лишённый пиреноида. Тилакоиды в хлоропласте одиночные, параллельно и концентрически расположенные (не в гранах), с фикобилисомами на внешней стороне. Основные пигменты тилакоида — хлорофилл a, фикоцианин-C, аллофикоцианин, каротиноиды. При делении хлоропласта образуется кольцо деления — кольцо электрон-плотного материала.
Митохондрия в клетке единственная, вогнутой формы. Вакуоли отсутствуют. Диктиосомы имеются.
Половой процесс не известен. Размножение осуществляется с помощью эндоспор, в клетке образуются 4 споры тетраэдрической формы, 2—3 мкм в диаметре[1].
Внутриклеточный уровень pH близок к 6,8—7. Протоны, пассивно проникающие в клетку из среды, выталкиваются обратно под действием АТФ-азы в клеточной мембране для обеспечения нормального функционирования внутриклеточных процессов[2].
Водоросль обладает цветочным запахом благодаря выделению летучих нор-каротиноидов[3], из клеток выделены 6-метилгептен-5-он-2, геранилацетон, 1,2-дигидро-1,1,6-триметилнафталин, β-ионон. Запасающие вещества в клетке — фитогликоген, флоридозиды и изофлоридозиды.
Число хромосом — n = 5[4].
Распространение и экология
Широко распространённый вид, известный из кислотных термических источников по всему миру, не обнаруженный в Африке и Антарктиде. Обитает в кислотных термальных источниках с естественными значениями pH от 0,05 до 5,0[1] и температурой от 13 до 57 °C. Образует слизистые налёты на камнях. При большей температуре увеличивается доля ненасыщенных жирных кислот в клетке. Оптимальное значение pH — 2—3; температурный оптимум — 45 °C. Во всех источниках, где обнаружен цианидиум, также обнаруживаются те или иные виды Galdieria, причём в различных источниках преобладают разные роды[5].
Способы, которыми водоросль, встречающаяся на небольших изолированных участках с экстремальными условиями и не образующая подвижных структур, либо структур, способных длительное время сохранять жизнеспособность при иных условиях, распространилась столь широко, не известны. Томас Брок (англ. Thomas D. Brock) в 1967 году предполагал, что цианидиум и другие современные экстремофилы встречались на Земле до образования твёрдой земной коры, что объясняет их наличие в отдалённых друг от друга местах, а также отсутствие на Гавайских островах, образовавшихся сравнительно недавно. Однако наличие цианидиума в Исландии и на Азорских островах, о чём писал Брок в 1978 году, эта гипотеза не объясняет. Цианидиум мог распространяться по близко расположенным термическим источникам при непосредственном наземном перетекании жидкости из одного источника в другой, однако даже при наличии постоянного перетока между источниками при различии их физических и химических характеристик микроорганизмы, как правило, новый источник не заселяют (что было показано при изучении нескольких травертин в Йеллоустоне). Источники могут быть связаны и подземными токами, однако широкого распространения водоросли это никак не объясняет, температуры под землёй часто оказываются выше переносимых организмами, и даже близкие гидротермальные источники могут не обладать подземной связью[5]. По мнению Топлина и соавторов (2008), распространение происходит с помощью птиц[6].
В качестве источников азота цианидиум может использовать как ионы аммония, так и нитрат-ионы благодаря синтезу термостойкой нитратредуктазы, а также глутаминсинтетазы.
История изучения и номенклатура
Из-за своего необычных строения, внешнего вида и экологических предпочтений цианидиум в разное время относили к совершенно разным группам организмов — к цианобактериям, к зелёным водорослям, к криптомонадам, иногда считали промежуточным звеном между красными и зелёными водорослями[7].
В настоящее время цианидиум считают представителем наиболее примитивных красных водорослей. Вместе с родами Galdieria и Cyanidioschyzon составляет класс Цианидиофициевые водоросли (Cyanidiophyceae), иногда принимаемый в качестве отдельного отдела Cyanidiophyta в составе подцарства Rhodoplantae, включающего также Красные водоросли и Глаукофитовые водоросли. Исследования последовательностей, кодирующих ген psbA (отвечающий за синтез белковой субъединицы D1 фотосистемы II) и 16S рРНК хлоропласта, а также генов малой и большой субъединиц Рубиско показали, что хлоропласт цианидиума сходен с хлоропластами других красных водорослей[8]. Предполагается, что цианидиевые отделились от других красных водорослей около 1,3 млн лет назад[5].
Научное название рода происходит от др.-греч. κυανός — «тёмно-синий»[9].
Первые сведения
Первое описание цианидиума было выполнено в 1839 году итальянским естествоиспытателем Джузеппе Менегини (итал. Giuseppe Meneghini; 1811—1889) на основании образцов, собранных в сернистых водах источников в Асколи-Пичено Антонио Орсини (итал. Antonio Orsini; 1788—1870)[7].
С принятым в настоящее время эпитетом водоросль была впервые описана одной из первых американских альгологов, Жозефиной Тилден (англ. Josephine Tilden; 1869—1957), в 1898 году. Тилден собранный 7 июля 1896 года в одном из источников Национального парка Йеллоустон образец водоросли описала в качестве экологической формы Protococcus botryoides Kütz., 1846 (Protosiphon botryoides (Kütz.) G.A.Klebs, 1896) Protococcus botryoides var. caldarius. Однако другой образец, собранный в тот же день и почти в том же месте, а также ещё три образца — собранный ей же 27 июня 1896 года и собранные известным геологом Уолтером Харви Уидом (англ. Walter Harvey Weed, 1862—1944) в 1897 году она отнесла к виду Chroococcus varius A.Braun. Хроококк — цианобактерия, представленная в виде отдельных сферических или полусферических клеток, собранных в колонии по менее 50 клеток, не образует эндоспор[7].
Ошибочность определения образцов как Chroococcus была показана в 1901 году Уильямом Альбертом Сетчеллом (англ. William Albert Setchell), отнёсшим все образцы к роду цианобактерий Pleurocapsa. В 1910 году Тилден согласилась как с определением Сетчелла, так и с отнесением образцов к этому роду[7][10].
В 1925 году австрийский ботаник и цитолог Лотар Гейтлер выделил новый род Chroococcopsis, в который включил в том числе и водоросль, описанную Тилден. В 1933 году он определил образцы, привезённые с немецкой экспедиции в Сундаланд (1928—1929), как этот вид и после их тщательного изучения опубликовал описание отдельного рода Cyanidium. Он неуверенно относил этот род к синезелёным водорослям, однако предполагал, что в клетках могут содержаться хлоропласт и ядро (качество имеющихся у него образцов не позволяло подтвердить это). В 1935 году он в соавторстве с Францем Руттнером описал семейство Cyanidiaceae, поместив его в порядок Хамесифоновые. Также Гейтлер и Руттнер предположили, что образцы, собранные в японских горячих источниках Гансом Молишем в 1924 году, могут относиться к этому виду[7].
В 1936 году Джозеф Коупленд (англ. Joseph Copeland), не имевший доступа к работам Гейтлера, выделил Pleurocapsa caldaria из Йеллоустона в род Pluto. Он назвал род по имени бога подземного царства из-за обитания водоросли в смертоносных местах — горячих кислотных источниках и грязевых вулканах[7].
В 1939 году Ондесс Ламар Инман (англ. Ondess Lamar Inman) обнаружил цианидиум в гейзерах Сономы. Его образцы были определены в 1940 году Фрэнсисом Эллиоттом Друэтом (англ. Francis Elliott Drouet) в род зелёных водорослей Хлорелла. В 1943 году Друэт, однако, изменил свою точку зрения относительно природы этих организмов и перенёс их в род цианобактерий Dermocarpa — водорослей с полярными клетками, хотя клетки цианидиума лишены полярности, на что было указано Коуплендом[7].
Обнаружение эукариотической природы и цитохимия
В 1944 году Кэнъитиро Негоро (яп. 根来健一郎) подтвердил предположение Гейтлера и Руттнера, обнаружив цианидиум в Японии. Негоро впервые смог выделить чистую культуру цианидиума, а также указал на наличие некоторого отлично окрашенного отделения в клетках. В 1947 году Хироюки Хиросэ (яп. 広瀬弘幸) назвал его пластидой, чётко увидел структуру хлоропласта[7].
Три года спустя Хиросэ обнаружил в клетках водоросли хлорофилл a, β-каротин и ксантофилл. Несмотря на то, что хлорофилла b клетки цианидиума не содержат, он предположил, что водоросль может быть отнесена к роду Хлорелла[7].
В 1952 году Мэри Белл Эллен (англ. Mary Belle Allen) указала на близость неопределённой водоросли, обнаруженной в гейзерах Сономы, виду Chlorella variegata Beij., однако вместо характерных для зелёных водорослей пигментов обнаружила в клетках фикоцианин и хлорофилл a. В 1954 году она отождествила эту водоросль с водорослью Тилден, как и Хиросэ, относя её к хлорелле. Пьер Буррелли (фр. Pierre Bourrelly) предположил, что хлорелла, описываемая Эллен, является либо жёлто-зелёной водорослью, либо криптомонадой. Гордон Эллиотт Фогг (англ. Gordon Elliott Fogg) в 1956 году склонялся к отнесению этого организма к коккоидным формам криптомонад[7].
Хиросэ продолжил изучение водоросли, в 1958 году методом окрашивания по Фёльгену установил наличие в её клетках оформленного ядра, а также указал на близость запасающих веществ багрянковому крахмалу и гликогену. Он пришёл к выводу, что цианидиум является представителем красных водорослей, посчитав необходимым переименовать этот род в Rhodococcus. Годом позже Гейтлер согласился с выводами японского исследователя, однако указал, что смена родового названия ничем не оправдана[7].
Эллен в 1959 году отказалась от родства водоросли хлорелле, не найдя отличия между одним из её пигментов и фикоцианином-C синезелёных водорослей, однако продолжала относить её к аномально окрашенным зелёным водорослям[7].
Электронная микроскопия и проблема с номенклатурой
В 1960-х годах проводились электронномикроскопические исследования клеток цианидиума, в частности, было показано наличие настоящего комплекса Гольджи. В 1970 году Паоло Де Лука (итал. Paolo De Luca) и Роберто Таддеи (итал. Roberto Taddei), заметив, что в описаниях цианидиума из разных мест мира присутствуют существенные расхождения (в частности, во многих работах указывалось на наличие в клетках вакуолей и на способность одной клетки образовывать до 32 эндоспор, часто приводились различные данные о размерах клеток), инициировали исследование популяций цианидиума на Флегрейских полях. Они пришли к выводу, что они представлены двумя различными филогенетическими линиями, названными Cyanidium caldarium forma A и Cyanidium caldarium forma B. В дальнейшем был установлен ряд различий между ними: forma A не содержит линоленовой кислоты, способна к ассимиляционному восстановлению нитратов, не усваивает глюкозу при отсутствии света и является облигатным автотрофом; forma B содержит линоленовую кислоту, не усваивает нитраты, но поглощает глюкозу как на свету, так и в темноте и является факультативным гетеротрофом. В 1976 году Де Лука и Таддеи пришли к выводу, что эти две формы — два различных вида организмов, часто встречающиеся совместно, данные о них в литературе смешаны[7][11].
Почти во всех кислотных источниках, исследованных в 1974—1981 годах в Европе, Азии и Северной Америке (в частности, в Йеллоустоне и Сономе), обнаруживались представители двух видов. В ряде источников был обнаружен третий вид, получивший название Cyanidioschyzon merolae De Luca, Taddei & Varano, 1978. Однако опровергнуть однородность типового образца Protococcus botryoides f. caldarius, а также доказать существование различий между образцами, отнесёнными к различным видам самой Тилден, оказалось невозможным ввиду их плохой сохранности. Образцы, которые можно с уверенностью отнести к forma B, были описаны всего годом позднее, чем таксон Тилден, как Pleurococcus sulphurarius Galdieri. Сетчелл в 1901 году и Гейтлер в 1933 году описывали водоросль, идентичную forma A. Таким образом, наиболее рациональным представляется сохранение названия, основанного на эпитете, используемом Тилден, Сетчеллом и Гейтлером, за forma A и принятие названия, основанного на базиониме Галдьери, за forma B — Galdieria sulphuraria (Galdieri) Merola[7].
Штамм, выделенный Эллен (известный в литературе под названием «штамм Аллен» — Allen strain), на основании которого проводились многочисленные исследования цианидиума, в действительности оказалась представителем Galdieria[11].
Исследования цианидиума в СССР и России
М. В. Гусев в 1961 году включил цианидиум в монографию синезелёных водорослей, однако указывал на его эукариотическую природу[12].
Представители цианидиевых в России обнаруживаются в горячих источниках на Дальнем Востоке. С 1979 года Л. М. Герасименко и Б. В. Громовым был выделен ряд чистых и смешанных культур с Камчатки, проводились исследования по изучению жизненного цикла водоросли, морфологии клетки, строения ДНК, пигментации, влиянию кислотности на скорость роста, фотосинтетическую активность, пигментацию.
В 1991 году О. Ю. Сенцова изучила разнообразие видов класса на Дальнем Востоке России, описав три новых вида рода Galdieria. Штамм Аллен был определён ей в новый вид Galdieria partita[11]. В 2003 году И. Н. Стадничук, И. О. Селях и О. В. Муравенко опубликовали статью, в которой впервые были приведены хромосомные числа цианидиума и нескольких видов Galdieria, установлены кариологические различия между родами[4].
Синонимы
- Рода
- Pluto J.J.Copel., 1936, nom. illeg.
- Rhodococcus Hirose, 1958, nom. illeg.
- Вида
- Chlorella caldaria (Tilden) Hirose, 1950
- Chroococcopsis caldaria (Tilden) Geitler, 1925
- Coccochloris orsiniana Menegh., 1839
- Dermocarpa caldaria (Tilden) F.E.Drouet, 1943
- Palmella orsiniana (Menegh.) Kütz., 1846
- Pleurocapsa caldaria (Tilden) Setch., 1901
- Pluto caldarius (Tilden) J.J.Copel., 1936
- Protococcus botryoides f. caldarius Tilden, 1898basionym
- Protococcus vulcanicus Ces., 1892, nom. inval.
- Rhodococcus caldarius (Tilden) Hirose, 1958
Цианидиум в пещерах
Различные авторы сообщали о неоднократных находках водорослей, практически неотличимых от цианидиума, в слабокислых, нейтральных и слабощелочных средах, в частности, в известняковых пещерах, где они нередко образуют моновидовые биоплёнки. Впервые такие водоросли обнаружил в 1936 году немецкий гидробиолог Герхард Гельмут Швабе (нем. Gerhard Helmut Schwabe) в пяти пещерах в Чили, описав их под названием Cyanidium chilense G.H.Schwabe. В 1942 году из-за очень большого сходства с водорослью кислотных источников он стал считать её экологической разновидностью Cyanidium caldarium var. chilense, а также выделил немного отличающуюся от неё форму Cyanidium caldarium f. rumpens[13].
В 1964 году Имре Фридман обнаружил водоросль, определённую как цианидиум, в пещерах Израиля[14]. Генрих Скуя в 1970 году сообщал о находке цианидиума в пещере в Италии, в 1983 году Ж.-К. Леклер, А. Кутэ и П. Дюпюи обнаружили и предварительно определили как цианидиум водоросль в пещере во Франции[13].
Швабе описывал одноклеточные водоросли, образующие синезелёные до чёрно-зелёных при большей толщине плёнки зернистой или порошкообразной массы на камнях. Клетки сферические, 3—6 мкм в диаметре, окружены прочной гомогенной клеточной стенкой. Размножаются образованием четырёх, реже двух (по Скуе) или восьми (f. rumpens у Швабе) спор 1,8—3,0 мкм в диаметре[13].
Наиболее толстые плёнки водоросли, согласно Швабе, а также Леклеру и соавторам, образуются на границе фотической зоны, на расстоянии до 40 м от входа в пещеру. Температура в пещерах в течение года изменяется незначительно, в пещере, исследованной Леклером и соавторами, она изменялась в пределах от 4 до 10 °C. Водоросль, описанная Швабе как f. rumpens, напротив, встречается в трещинах на камнях, подверженных довольно резким суточным перепадам температуры и прямому солнечному свету в дневное время. Водоросль никогда не встречается на постоянно смачиваемых участках стен пещер, влажность воздуха в местах произрастания пещерного цианидиума, однако, должна оставаться высокой и постоянной (в пещере, описанной у Леклера — между 89 и 95 %). Уровень pH в пещерах, описанных Швабе и другими авторами, всегда превышал 5,0, в известняковых пещерах он всегда выше 7,0[13].
На основании филогенетических данных были описаны три различных организма, предварительно названные Cyanidium sp. Monte Rotaro, Cyanidium sp. Sybil cave и Cyanidium sp. Atacama. Было показано, что они образуют оформленную монофилетичную группу, действительно близкую обитателю кислотных источников. На основе анализа 16S рРНК было получено следующее филогенетическое древо[15]:
Cyanidioschyzon merolae Strain 10D AB002583
Cyanidioschyzon sp. Clone SK114 AY882672
Galdieria maxima Strain IPPAS P507 AY391361
Cyanidium sp. Atacama FJ390400
Cyanidium sp. Monte Rotaro AY391359
Cyanidium sp. Sybil cave AY391360
Cyanidium caldarium AF170718
Cyanidium caldarium X52985
Cyanidium caldarium Strain YSK-1 EU586032
Cyanophora paradoxa X81840
В настоящее время пещеры, исследованные Швабе, не определены и применить к одному из этих таксонов название, использованное немецким исследователем, невозможно. Франклин Отт (англ. Franklyn Ott) в 1993 году предложил считать их самостоятельными видами рода Цианидиум, в который он также включил всех представителей Galdieria. Систематическое положение не ацидофильных цианидиофициевых остаётся неопределённым.
Примечания
- R. G. Sheath & M. L. Vis. Red Algae // Freshwater Algae of North America: Ecology and Classification / edited by J. D. Wehr, R. G. Sheath & J. P. Kociolek. — 2nd edition. — 2015. — P. 248. — 1045 p. — ISBN 978-0-12-385876-4.
- I. Enami, H. Adachi & J.R. Shen. Mechanisms of Acido-Tolerance and Characteristics of Photosystems in an Acidophilic and Thermophilic Red Alga, Cyanidium caldarium // Red Algae in the Genomic Age / edited by J. Seckbach & D. J. Chapman. — Dordrecht, New York, 2010. — P. 375—389. — 498 p. — (Cellular Origin, Life in Extreme Habitats and Astrobiology, V. 13). — ISBN 978-90-481-3795-4.
- Jüttner, F. Nor-Carotenoids as the Major Volatile Excretion Products of Cyanidium (англ.) // Zeitschrift fur Naturforschung : journal. — 1979. — Vol. 34, no. 3—4. — P. 186—191.
- Стадничук, И. Н., Селях, И. О., Муравенко, О. В. Хромосомные числа видов микроводорослей класса Cyanidiophyceae (Rhodophyta) // Ботанический журнал. — 2003. — Т. 88, № 4. — С. 175—176.
- V. Reeb & D. Bhattacharya. The Thermo-Acidophilic Cyanidiophyceae (Cyanidiales) // Red Algae in the Genomic Age / edited by J. Seckbach & D. J. Chapman. — Dordrecht, New York, 2010. — P. 411—427. — 498 p. — (Cellular Origin, Life in Extreme Habitats and Astrobiology, V. 13). — ISBN 978-90-481-3795-4.
- Toplin, J.A., Norris, T.B., Lehr, C.R., McDermott, T.R. and Castenholz, R.W. Biogeographic and phylogenetic diversity of thermoacidophilic Cyanidiales in Yellowstone National Park, Japan, and New Zealand (англ.) // Applied and Environmental Microbiology : journal. — American Society for Microbiology, 2008. — Vol. 74, no. 9. — P. 2822—2833.
- F. D. Ott & J. Seckbach. A review on the taxonomic position of the algal genus Cyanidium Geitler 1933 and its ecological cohorts Galdieria Merola in Merola et al. 1981 and Cyanidioschyzon De Luca, Taddei and Varano 1978 // Evolutionary Pathways and Enigmatic Algae: Cyanidium caldarium (Rhodophyta) and Related Cells. — Dordrecht, 1993. — P. 113—132. — 344 p. — (Developments in Hydrobiology; 91). — ISBN 978-94-010-4381-6.
- D. J. Chapman. Enigmatic unicellular Protista: are they really enigmatic? The Algae case // Enigmatic microorganisms and life in extreme habitats / edited by J. Seckbach. — 1999. — P. 102—104. — (Cellular origin and life in extreme habitats; V. 1). — ISBN 978-1-4020-1863-3.
- Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. Ботаника: в 4 т. Т. 2. — М.: Издат. центр «Академия», 2006. — С. 180—181. — 320 с. — ISBN 978-5-7695-2750-1.
- J. Tilden. Minnesota Algae. — Minneapolis, 1910. — P. 47. — 328 p.
- Сенцова, О. Ю. О разнообразии одноклеточных ацидотермофильных водорослей рода Galdieria (Rhodophyta, Cyanidiophyceae) // Ботанический журнал. — 1991. — Т. 76, № 1. — С. 69—79.
- Гусев, М. В. Синезелёные водоросли // Микробиология. — 1961. — Т. 30, № 6. — С. 1108—1128.
- L. Hoffmann. Cyanidium-like algae from caves // Evolutionary Pathways and Enigmatic Algae: Cyanidium caldarium (Rhodophyta) and Related Cells. — Dordrecht, 1993. — P. 175—182. — 344 p. — (Developments in Hydrobiology; 91). — ISBN 978-94-010-4381-6.
- Friedmann, I. Progress in the biological exploration of caves and subterranean waters in Israel (англ.) // International Journal of Speleology : journal. — 1964. — Vol. 1. — P. 29—33.
- A. Azúa-Bustos & R. Vicuña. Chilean Cave Cyanidium // Red Algae in the Genomic Age / edited by J. Seckbach & D. J. Chapman. — Dordrecht, New York, 2010. — P. 429—439. — 498 p. — (Cellular Origin, Life in Extreme Habitats and Astrobiology, V. 13). — ISBN 978-90-481-3795-4.
Литература
- Evolutionary Pathways and Enigmatic Algae: Cyanidium caldarium (Rhodophyta) and Related Cells / edited by J. Seckbach. — Dordrecht, 1993. — 344 p. — (Developments in Hydrobiology; 91). — ISBN 978-94-010-4381-6.
- Виноградова, К. Л. Красные водоросли // Определитель пресноводных водорослей СССР. 13. Зеленые, красные и бурые водоросли / Отв. ред. М. М. Голлербах. — Л.: Наука, 1980. — С. 178. — 248 с.
- P. Albertano, C. Ciniglia, G. Pinto & A. Pollio. The taxonomic position of Cyanidium, Cyanidioschyzon and Galdieria: an update (англ.) // Hydrobiologia : journal. — 2000. — Vol. 433. — P. 137—143.