Тёмная акула
Тёмная акула[1][2], или белопёрая акула[2], или сумеречная акула[2][3], или тёмная акула-собака[2] (лат. Carcharhinus obscurus) — хищная рыба семейства серых акул отряда Кархаринообразных. Эти акулы обитают в тропических водах и многих умеренных областях всех океанов. Они встречаются на глубине до 400 м. Совершают сезонные миграции, летом перемещаясь ближе к полюсам, а зимой к экватору. У них стройное, обтекаемое тело с широким и округлым рылом. Второй спинной плавник намного меньше первого. Грудные плавники крупные, серповидной формы. Между спинными плавниками имеется гребень. Окраска дорсальной поверхности тела от бронзового до голубовато-серого цвета. Максимальная зарегистрированная длина 420 см, а масса 347 кг.
| Тёмная акула | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Группа: Класс: Подкласс: Инфракласс: Надотряд: Клада: Galeomorphi Отряд: Семейство: Подсемейство: Серые, или пилозубые акулы Триба: Carcharhinini Род: Вид: Тёмная акула |
||||||||||
| Международное научное название | ||||||||||
| Carcharhinus obscurus Lesueur, 1818 | ||||||||||
| Синонимы | ||||||||||
|
Carcharhinus iranzae Fourmanoir, 1961
|
||||||||||
| Ареал | ||||||||||
 |
||||||||||
| Охранный статус | ||||||||||
| ||||||||||

Рацион тёмных акул очень разнообразен. Они охотятся на разнообразных костистых и хрящевых рыб, а также морских беспозвоночных. На морских млекопитающих нападают довольно редко. Подобно прочим представителям рода серых акул, тёмные акулы являются живородящими. В помёте от 3 до 16 новорождённых. Беременность длится до 24 месяцев. Цикл репродукции трёхгодичный. Тёмные акулы очень медленно растут и поздно достигают половой зрелости. Эти акулы потенциально опасны для человека. Они являются объектом коммерческого промысла. Медленный цикл воспроизводства делает их очень чувствительными к негативному воздействию, обусловленному человеческими факторами[4][5].
Таксономия и филогенез
Французский натуралист Шарль Александр Лесюёр опубликовал первое научное описание тёмной акулы в 1818 году в номере журнала Академии естественных наук Филадельфии. Он отнёс её к роду Squalus и дал видовой эпитет лат. obscurus, что означает «тёмная» или «тусклая», ссылаясь на окраску рыбы[6]. Последующие авторы отнесли этот вид к роду Carcharhinus. Голотип назначен не был, хотя известно, что Лесюёр исследовал особь, пойманную, вероятно, в североамериканских водах[7].
Многие ранние источники использовали в качестве научного названия тёмной акулы Carcharias (позже Carcharhinus) lamiella, данное этому виду в 1882 году Дэвидом Старром Джордананом и Чарльзом Генри Гилбертом. Позднее выяснилось, что описанные учёными челюсти принадлежали не тёмной, а узкозубой акуле (Сarcharhinus brachyurus). Таким образом, Сarcharhinus lamiella считается синонимом не Сarcharhinus obscurus, а Сarcharhinus brachyurus[8][9].
В палеонтологической литературе часто упоминаются окаменелые зубы тёмных акул, хотя видовая идентификация зубов Carcharhinus довольно проблематична[10]. Эти находки, относящиеся к эпохе миоцена (25—5,3 млн лет назад), были обнаружены в формациях Кендис и Гранд Бэй в Карриаку, Гренадины[11], Мохра (Египет)[12], Полк (округ Флорида) и, вероятно, Серро ла Круз на севере Венесуэлы[13]. Кроме того, в формации Йорктаун и в Пунго ривер, Северная Каролина, а также в Чесапикском заливе, часто находят зубы, датируемые поздним миоценом и ранним плиоценом (11,6—3,6 млн лет назад), которые лишь немногим отличаются от зубов современных тёмных акул и ошибочно принимаются за зубы длиннокрылых акул[10]. Зубы тёмных акул были извлечены из окаменелых останков двух усатых китов, из которых один сохранился в формации Гуз Крик Лаймстоун, Северная Каролина, и датировался эпохой раннего плиоцена (около 3,5 млн лет назад), а другой был найден в грязевых отложениях эпохи плейстоцена-голоцена (примерно 12 000 лет назад)[14]
В 1982 году новозеландский учёный Джек Гаррик опубликовал результаты филогенетического анализа Carcharhinus на основе морфологии и поместил тёмную и галапагосскую акул (Carcharhinus galapagensis) в центре группы «obscurus». В состав группы вошли крупные акулы с треугольными зубами и гребнем между спинными плавниками, такие как Carcharhinus altimus, Carcharhinus perezi, серо-голубая и длиннокрылая акула[15]. Эта интерпретация была широко поддержана результатами фенетических исследований[16] и анализа аллозимной последовательности. Последний позволил установить взаимоотношения внутри ветви Carcharhinus, для членов которой характерно наличие межплавникового дорсального гребня. Было обнаружено, что галапагосские, серо-голубые и длиннокрылые акулы составляют производную кладу[17].
| |||||||||||||||||||||||||||||||||||||||
| Филогенетические взаимоотношения тёмной акулы, установленные на основании аллозимной последовательности[17] |
Описание
Один из крупнейших членов рода серых акул, тёмная акула обычно достигает длины 3,2 м при массе 160—180 кг. Максимальная зафиксированная длина и масса составляют 4,2 м и 347 кг соответственно[4][18]. Самки вырастают крупнее самцов[19]. У этой акулы стройное, обтекаемое тело с широким округлым рылом. Перед ноздрями имеются едва развитые кожные складки. Круглые глаза среднего размера оснащены мигательной мембраной. Рот очень короткий, по углам расположены тонкие борозды. Во рту по 13—15 (обычно 14) зубных рядов с каждой стороны обеих челюстей. Верхние зубы широкие, треугольные, стоят слегка под наклоном, края сильно зазубрены; нижние зубы ́уже и стоят в вертикальном положении, зазубрины по краям мельче. У тёмной акулы имеется пять пар довольно длинных жаберных щелей[18]. Большие грудные плавники составляют около 1/5 длины тела акулы, имеют серповидную форму, кончики заострены. Первый спинной плавник умеренного размера и нечёткой серповидной формы, у него заострённая вершина и сильно вогнутый задний край. Основание плавника лежит над свободными задними кончиками грудных плавников. Второй спинной плавник значительно меньше первого и расположен напротив анального плавника. Между первым и вторым спинными плавниками имеется гребень. Хвостовой плавник крупный и высокой, с хорошо развитой нижней лопастью и вентральной вырезкой под кончиком верхней лопасти[20]. Плакоидная чешуя имеет форму ромба, чешуйки близко посажены, на каждой имеется по пять горизонтальных зубцов[18]. Окраска от бронзового до голубовато-серого цвета, брюхо белое, белый цвет распространяется по бокам, где могут быть видны слабые светлые полосы. Плавники, особенно нижняя часть грудных и нижняя лопасть хвостового плавника, по краям темнее. Эта характерная окраска более ярко выражена у молодых акул[21].
Ареал

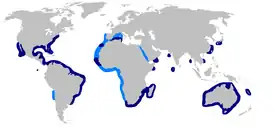
Тёмная акула встречается по всему миру в тропических и тёплых умеренных водах. Её ареал обширен, хотя и прерывист. В западной Атлантике эти акулы обитают от штата Массачусетс и Джоржс Бэнк до Мексиканского залива и от Никарагуа до южной части Бразильского побережья. В восточной Атлантике они попадаются на западе Средиземного моря, у берегов Канарских островов, Кабо-Верде, Сенегала, Сьерра-Леоне и, возможно, в других местах. В Индийском океане они населяют прибрежные воды Южной Африки, Мозамбика и Мадагаскара. Изредка поступают данные о присутствии этих акул в Аравийском море, Бенгальском заливе и, возможно, Красном море. В западной части Тихого океана встречаются от Японии до штатов Квинсленд и Новый Южный Уэльс, Австралия. В восточной части Тихого океана ареал охватывает побережье Американского континента от южной Калифорнии до Чили[5]. Информация о присутствии тёмных акул в северо-восточной и центрально-восточной частях Атлантического океана, вероятно, ошибочна и относится к галапагосским акулам[5][22]. Исследования митохондриальной ДНК и микросателлитов показали, что акулы, обитающие в водах Индонезии и Австралии, принадлежат к отдельным популяциям[23].
Тёмные акулы обитают на континентальном прибережье от зоны прибоя до внешнего края континентального шельфа и прилегающих океанических вод, деля пространство с более специализированными сородичами, такими как предпочитающие держаться у берега серо-голубые акулы, пелагические шёлковые акулы (Сarcharhinus falciformis), океанические длиннокрылые акулы, глубоководный вид Carcharhinus altimus и островные галапагосские и белопёрые серые акулы (Carcharhinus albimarginatus)[7]. В ходе исследования, проведённого путём мечения в северной части Мексиканского залива, было обнаружено, что тёмные акулы большую часть своего времени остаются на глубине 10—80 м, периодически опускаясь ниже 200 м. Известно, что этот вид способен достичь глубины в 400 м. Он предпочитает температуру воды 19—28 °C и избегает зон с пониженной солёностью, таких как устья рек[8][24].
Тёмные акулы совершают миграции на расстояние до 3800 км. У берегов Северной Америки летом они перемещаются на север, а зимой к экватору[5]. В водах Южной Африки молодые самки и самцы, достигшие размера 0,9 м, уплывают к югу и к северу от места рождения Квазулу-Наталь и через несколько лет по неизвестным маршрутам присоединяются к взрослым. Кроме того, подрастающие особи проводят весну и летом в зоне прибоя, а осень и зиму — в открытом море. По достижении в длину 2,2 м они начинают совершать миграции с севера на юг между Квазулу-Наталь в зимнее время и Вестерн Кейп летом. Крупные акулы, чей размер превышает 2,8 м, мигрируют на юг вплоть до Мозамбика[5][7][25]. У берегов Западной Австралии взрослые и молодые тёмные акулы летом и осенью подплывают к побережью, не заходя в мелководные заливы и бухты, где появляются на свет новорождённые[5].
Биология и экология
Будучи хищниками, занимающими вершину пищевой пирамиды, тёмные акулы обычно имеют меньшую численность по сравнению с другими акулами, которые разделяют их ареал[7]. Однако есть места, где встречаются скопления этих акул, особенно молодых[8]. Взрослые акулы часто сопровождают корабли далеко от земли, например, по течению мыса Игольного[25]. Исследования, проведённые в устье реки Кейп-Фир в штате Северная Каролина, сообщили о том, что средняя скорость их движения составляет 0,8 км/ч[26]. Тёмных акул часто сопровождают рыбы-прилипалы (Echeneis naucrates)[27]. Взрослые тёмные акулы не имеют врагов в природе[18]. На молодых акул охотятся обыкновенная песчаная акула (Carcharias taurus), большая белая акула (Carcharodon Carcharias), акула-бык (Сarcharhinus leucas) и тигровая акула (Galeocerdo cuvier). У берегов Квазулу-Наталь использование сетей для защиты пляжей от акул привело к сокращению популяции крупных хищников. Это вызвало резкое увеличение числа неполовозрелых тёмных акул, которые, в свою очередь, уничтожили небольших костистых рыб, что имело в целом негативные последствия для биоразнообразия местной экосистемы[7][28].
На тёмных акулах паразитируют ленточные черви Anthobothrium laciniatum[29], Dasyrhynchus pacificus[30], Platybothrium kirstenae[31], Floriceps saccatus[32], Tentacularia coryphaenae[33] и Triloculatum triloculatum[34], моногенеи Dermophthirius carcharhini[35], Loimos salpinggoides[36], пиявки Stibarobdella macrothela[37], копеподы Alebion sp., Pandarus cranchii[38], P. sinuatus[18] и морские миноги[39]
Рацион и пищевое поведение
Тёмная акула является универсальным хищником, в рацион которой входит широкий спектр морских животных на всех уровнях водной толщи[21][40]. Крупные особи могут потребить более 1/10 пищи от своего собственного веса за один приём[41]. Сила укуса 2-метровой тёмной акулы составляет 60 кг на 2 мм² (площадь вершины зуба). Это самый высокий показатель, достоверно измеренный у какой-либо акулы[42]. В Индийском океане были зарегистрированы плотные скопления молодых тёмных акул на участках с большой концентрацией пищи[5].
Рацион тёмной акулы включает в себя пелагических костистых рыб, таких как сельди и анчоусы, тунцы и скумбрии, марлины, ставриды, рыбы-иглы, летучие рыбы, рыбы-сабли; донных рыб, в том числе кефалей, морских карасей, горбылей, скорпен, угрей и камбал; рифовых рыб, в том числе барракуд, рыб-ежей, барабулей, веслоносов и морских окуней; хрящевых рыб, в том числе акул, рыб-пил, морских ангелов (Squalus squatina), куньих акул, рыб-молотов, лисьих акул, рыб-гитар, скатов, и беспозвоночных, в том числе головоногих, ракообразных, моллюсков и морских звёзд. Очень редко крупные особи поедают морских черепах, морских млекопитающих (в основном как падаль) и человеческий мусор[7][8][40][43].

В северо-западной Атлантике около 60 % рациона тёмной акулы составляют разные рыбы, принадлежащие к более чем 10 семействам, из которых главную роль играют луфари (Pomatomus saltatrix) и летние паралихты (Paralichthys dentatus). Вторым компонентом диеты являются скаты, их яйцевые капсулы. Крабы Ovalipes ocellatus — ещё один значительный источник пищи[40]. В южноафриканских и австралийских водах основу рациона составляют костистые рыбы. Новорождённые и молодые акулы охотятся в основном на мелких пелагических рыб и головоногих, таких как сардины и кальмары.

Акулы длиной свыше 2 м расширяют свой рацион за счёт крупных костистых и хрящевых рыб[44][45]. Каждую зиму у восточного побережья Южной Африки появляются южноафриканские сардины (Sardinops sagax), на которых охотятся тёмные акулы среднего и крупного размеров. Беременные и родившие самки не присоединяются к охоте, возможно, потому, что беременность не позволяет им тратить много энергии, преследуя такую быструю добычу[41]. Одно из исследований показало, что 0,2 % от рассмотренных тёмных акул охотились на афалин[46].
Размножение и жизненный цикл
Подобно прочим представителям рода серых акул, тёмные акулы являются живородящими рыбами; развивающиеся эмбрионы получают питание посредством плацентарной связи с матерью, образованной опустевшим желточным мешком. В северо-западной Атлантике спаривание происходит весной, в то время как в других районах, например у берегов Южной Африки, репродуктивный цикл не носит сезонного характера[7][8]. Самки способны сохранять сперму, вероятно, нескольких самцов, в течение длительного периода от нескольких месяцев до нескольких лет в нидаментальной железе (орган, который выделяет зародышевые сумки). Это важный фактор, обеспечивающий успешное воспроизводство, учитывая совершаемые тёмными акулами длительные миграции и малую концентрацию популяции, которые делают внутривидовые встречи редкими и случайными[47].

Беременность длится до 22—24 месяцев, самка приносит потомство один раз в три года[5]. В помёте от 3 до 16 новорождённых, среднее количество 6—12, численность потомства не зависит от размеров самки[8][41]. В западной части Атлантического океана, как правило, помёт меньше, чем в юго-восточной части Атлантического океана (в среднем 8 против 10 новорождённых)[18]. В зависимости от региона роды протекают круглый год или сезонно: в северо-западной Атлантике отмечалось появление на свет новорождённых в конце зимы, а у берегов Западной Австралии летом и осенью, в водах Южной Африки тёмные акулы рождаются в любой сезон[7][21]. С приближением срока родов беременные самки перемещаются в мелкие прибрежные лагуны, где много пищи и нет крупных хищников (в том числе своего вида), и сразу после появления потомства на свет уплывают[8][45]. Такие природные питомники есть на побережье провинции Квазулу-Наталь, на юго-западе Австралии, в Западной Калифорнии и на востоке Соединённых Штатов в Нью-Джерси и Северной Каролине[5][8].
Длина новорождённых составляет 0,7—1,0 м[5]; их размер напрямую связан с размером матери и уменьшается с численностью приплода. Имеющиеся данные указывают на то, что самки могут определить оптимальный размер, при котором их детеныши должны появиться на свет, с тем, чтобы повысить их шансы на выживание в зависимости от условий окружающей среды. Кроме того, при рождении вес печени акулят составляет 1/5 их веса, и они используют запасённые в печени питательные вещества, пока не научатся охотиться самостоятельно[41]. Тёмные акулы — одни из самых медленнорастущих видов акул. Половой зрелости они достигают лишь становясь крупными[48]: в зависимости от региона обитания самцы достигают половой зрелости при длине 2,8—3 м в возрасте 18—23 лет, для самок это 2,6—3,1 м и 17—32 года соответственно[49][50][51]. Ежегодный прирост составляет 8—11 см в течение первых пяти лет жизни[52]. Максимальная продолжительность жизни 40—50 лет и более[48].
Взаимодействие с человеком

Тёмные акулы из-за больших размеров считаются потенциально опасными для человека, хотя мало известно о том, как они ведут себя по отношению к людям под водой[7]. К 2014 году в списке International Shark Attack File было упомянуто 2 нападения на людей и лодки, одно из которых привело к летальному исходу[53]. Вероятно, нападения, приписываемые этому виду у Бермудских островов, на самом деле были совершены галапагосскими акулами. Для защиты пляжей в Южной Африке и Австралии активно используют противоакульи сети, в которые попадаются в основном взрослые особи. С 1978 по 1999 год у берегов Квазулу-Наталь в среднем ежегодно отлавливали 256 тёмных акул[48]. Молодые тёмные акулы хорошо приспосабливаются к жизни в неволе в аквариумах[7]. Тёмная акула является одним из наиболее востребованных видов акул на рынке, так как у неё крупные плавники с большим количеством внутренних лучей (ceratotrichia), которые используются для приготовления популярного супа[48]. Кроме того, мясо поступает в продажу свежим, замороженным, сушёным, солёным или копчёным, используют кожу и жир[4]. Тёмные акулы являются предметом целевого промышленного рыболовства у берегов восточной части Северной Америки, юго-западной Австралии и восточной Южной Африки. В водах юго-западной Австралии промысел начался в 1940 году и был расширен в 1970-е годы, когда вылавливалось по 500—600 тонн акул в год. Для добычи тёмных акул используются донные жаберные сети, в которые попадаются почти исключительно молодые акулы в возрасте до трёх лет, из них 18—28 % от всех новорождённых в первый год жизни. Демографические модели показывают, что промысел будет оставаться устойчивым при условии, что уровень смертности среди акул более 2 м в длину составит менее 4 %[48]. В дополнение к промышленному рыболовству тёмные акулы попадаются в качестве прилова в ярусы, предназначенные для тунца и рыбы-меч (в таких случаях туши акул обычно выбрасывают, сохраняя лишь ценные плавники). Этот вид ценится в спортивном рыболовстве. Большое количество тёмных акул, в основном неполовозрелых, вылавливают рыболовы-любители у берегов Южной Африки и Восточной Австралии. До резкого сокращения численности эта акула была одним из важнейших видов в турнирах Florida trophy shark[48].
Меры по сохранению вида
Международный союз охраны природы (МСОП) оценил статус сохранности этого вида как «Близкий к уязвимому положению» (NT) во всём мире и «Уязвимый» (VU) в северо-западной Атлантике и Мексиканском заливе. Американское общество рыболовов оценило статус североамериканской популяции тёмных акул как «Уязвимый»[18]. Очень низкий уровень репродуктивного цикла делает этот вид чувствительным к перелову. Проведённая в 2006 году американской Национальной службой морского рыболовства оценка запасов показала, что численность сократилась до 15—20 % от уровня 1970-х годов. В 1998 году была запрещена коммерческая и любительская ловля тёмной акулы, но эти меры имели ограниченную эффективность в силу высокой смертности рыб из-за прилова. Кроме того, несмотря на запрет, в 2003 году рыболовами-любителями были пойманы около 2000 тёмных акул. В 2005 году Северная Каролина объявила о временном прекращении любительского промысла[54]. Для облегчения усилий по сохранению разработан метод молекулярной идентификации, позволяющий определить видовую принадлежность акульих плавников, поступивших на рынок, чтобы выявлять случаи браконьерства[55].
Примечания
- Линдберг Г. У., Герд А. С., Расс Т. С. Словарь названий морских промысловых рыб мировой фауны. — Ленинград: Наука, 1980. — С. 39. — 562 с.
- Решетников Ю. С., Котляр А. Н., Расс Т. С., Шатуновский М. И. Пятиязычный словарь названий животных. Рыбы. Латинский, русский, английский, немецкий, французский. / под общей редакцией акад. В. Е. Соколова. — М.: Рус. яз., 1989. — С. 31. — 12 500 экз. — ISBN 5-200-00237-0.
- Губанов Е. П., Кондюрин В. В., Мягков Н. А. Акулы Мирового океана: Справочник-определитель. — М.: Агропромиздат, 1986. — С. 155. — 272 с.
- Тёмная акула (англ.) в базе данных FishBase.
- Carcharhinus obscurus (англ.). The IUCN Red List of Threatened Species.
- Lesueur C. A. Description of several new species of North American fishes (part 1) // Journal of the Academy of Natural Sciences of Philadelphia. — 1818. — Vol. 1, № (2). — P. 222—235.
- Compagno L. J. V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. — Rome: Food and Agricultural Organization, 1984. — P. 489—491. — ISBN 92-5-101384-5.
- Ebert D.A. Sharks, Rays, and Chimaeras of California. — California: University of California Press, 2003. — P. 149—152. — ISBN 0520234847.
- Jordan D. S., Gilbert C. H. Description of a new shark (Carcharias lamiella) from San Diego, California // Proceedings of the United States National Museum. — 1882. — Vol. 5, № 269. — P. 110—111.
- Heim, B. and Bourdon, J. Species from the Fossil Record: Carcharhinus obscurus. The Life and Times of Long Dead Sharks (20 апреля 2009). Дата обращения: 18 мая 2015.
- Portell R. W., Hubbell G., Donovan S. K., Green J. K., Harpe D. A. T., Pickerill R. Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles // Caribbean Journal of Science. — 2008. — Vol. 44, № (3). — P. 279—286.
- Cook T. D., Murray A. M., Simons E. L., Attia Y. S., Chatrath P. A. Miocene selachian fauna from Moghra, Egypt // Historical Biology: an International Journal of Paleobiolog. — 2010. — P. 1029—2381.
- Sanchez-Villagra M. R., Burnham R. J., Campbell D. C., Feldmann R. M., Gaffney E. S., Kay R. F., Lozsan R., Purdy R., Thewissen J. G. M. A New Near-Shore Marine Fauna and Flora from the Early Neogene of Northwestern Venezuela // Journal of Paleontology. — 2000. — Vol. 74, № 5. — P. 957—968. — doi:10.1666/0022-3360(2000)074<0957:ANNSMF>2.0.CO;2.
- Cicimurri D. J., Knight J. L. Two Shark-bitten Whale Skeletons from Coastal Plain Deposits of South Carolina // Southeastern Naturalist. — 2009. — Vol. 8, № (1). — P. 71—82. — doi:10.1656/058.008.0107.
- Garrick J. A. F. Sharks of the genus Carcharhinus // NOAA Technical Report, NMFS Circ.. — 1982. — Vol. 445.
- Compagno L. J. V. Sharks of the Order Carcharhiniformes. — Princeton University Press. — P. 319—320. — ISBN 0-691-08453-X.
- Naylor G. J. P. The phylogenetic relationships among requiem and hammerhead sharks: inferring phylogeny when thousands of equally most parsimonious trees result // Cladistics. — 1999. — Vol. 8, № (4). — P. 295—318. — doi:10.1111/j.1096-0031.1992.tb00073.x.
- Knickle Craig. Biological report: duscky shark (недоступная ссылка). Florida Museum of Natural History Ichthyology Department. Дата обращения: 18 мая 2015. Архивировано 4 января 2016 года.
- Natanson L. J., Casey J.G., Kohler N.E. Age and growth estimates for the dusky shark, Carcharhinus obscurus, in the western North Atlantic Ocean // Fishery Bulletin. — 1995. — Vol. 93, № 1. — P. 116—126.
- McEachran J. D., Fechhelm J. D. Fishes of the Gulf of Mexico: Myxinformes to Gasterosteiformes. — University of Texas Press, 1998. — P. 87. — ISBN 0-292-75206-7.
- Last P. R., Stevens J.D. Sharks and Rays of Australia. — 3-е. — Harvard University Press, 1993. — P. 254—255. — ISBN 0674034112.
- Compagno L., Dando M., Fowler S. Sharks of the World. — Princeton University Press, 2005. — P. 302—303. — ISBN 978-0-691-12072-0.
- Ovenden J. R., Kashiwagi T., Broderick D., Giles J., Salini J. The extent of population genetic subdivision differs among four co-distributed shark species in the Indo-Australian archipelago // BMC Evolutionary Biology. — 2009. — Vol. 9, № 40. — doi:10.1186/1471-2148-9-40. — PMID 19216767.
- Hoffmayer E. R., Franks J. S., Driggers W. B. (III), Grace M. A. Movements and Habitat Preferences of Dusky (Carcharhinus obscurus) and Silky (Carcharhinus falciformis) Sharks in the Northern Gulf of Mexico: Preliminary Results // 2009 MTI Bird and Fish Tracking Conference Proceedings. — 2009.
- Van der Elst R. A Guide to the Common Sea Fishes of Southern Africa. — 3-е. — Struik, 1993. — P. 35. — ISBN 1868253945.
- Huish M. T., Benedict C. Sonic tracking of dusky sharks in Cape Fear River, North Carolina // Journal of the Elisha Mitchell Scientific Society. — 1977. — Vol. 93, № (1). — P. 21—26.
- Schwartz F. J. Five species of sharksuckers (family Echeneidae) in North Carolina // Journal of the North Carolina Academy of Science. — 2004. — Vol. 120, № (2). — P. 44—49.
- Martin R. A. A Place For Sharks. ReefQuest Centre for Shark Research. Дата обращения: 22 мая 2015.
- Ruhnke T. R., Caira J. N. Two new species of Anthobothrium van Beneden, 1850 (Tetraphyllidea: Phyllobothriidae) from carcharhinid sharks, with a redescription of Anthobothrium laciniatum Linton, 1890 // Systematic Parasitology. — 2009. — Vol. 72, № (3). — P. 217—227. — doi:10.1007/s11230-008-9168-0. — PMID 19189232.
- Beveridge I., Campbell R. A. A revision of Dasyrhynchus Pintner (Cestoda, Trypanorhyncha), parasitic in elasmobranch and teleost fishes // Systematic Parasitology. — 1993. — Vol. 24, № 2. — P. 129—157. — doi:10.1007/BF00009597.
- Healy C. J. A revision of Platybothrium Linton, 1890 (Tetraphyllidea : Onchobothriidae), with a phylogenetic analysis and comments on host-parasite associations // Systematic Parasitology. — 2003. — Vol. 56, № 2. — P. 85—139. — doi:10.1023/A:1026135528505. — PMID 14574090.
- Linton E. Rhynchobothrium ingens spec. nov., a parasite of the dusky shark (Carcharhinus obscurus) // Journal of Parasitology. — 1921. — Vol. 8, № 1. — P. 22—32. — doi:10.2307/3270938.
- Knoff M., De Sao C. S. C., Pinto R. M., Lanfredi R. M., Gomes D. C. New records and expanded descriptions of Tentacularia coryphaenae and Hepatoxylon trichiuri homeacanth trypanorhynchs (Eucestoda) from carcharhinid sharks from the State of Santa Catarina off-shore, Brazil // Revista Brasileira de Parasitologia Veterinaria. — 2004. — Vol. 13, № 2. — P. 72—80.
- Caira J. N., Jensen K. Erection of a new onchobothriid genus (Cestoda: Tetraphyllidea) and the description of five new species from whaler sharks (Carcharhinidae) // Journal of Parasitology. — 2009. — Vol. 95, № 4. — P. 924—940. — doi:10.1645/GE-1963.1. — PMID 19271790.
- Bullard S. A., Dippenaar S. M., Hoffmayer E. R., Benz G. W. New locality records for Dermophthirius carcharhini (Monogenea : Microbothriidae) and Dermophthirius maccallumi and a list of hosts and localities for species of Dermophthirius // Comparative Parasitology. — 2004. — Vol. 71, № 1. — P. 78—80. — doi:10.1654/4093.
- MacCullum G. A. Some new forms of parasitic worms // Zoopathologica; scientific contributions of the New York Zoological Society. — 1917. — Vol. 1, № 2. — P. 1—75.
- Yamauchi T., Ota Y., Nagasawa K. Stibarobdella macrothela (Annelida, Hirudinida, Piscicolidae) from Elasmobranchs in Japanese Waters, with New Host Records // Biogeography. — 2008. — Vol. 10. — P. 53—57.
- Newbound D. R., Knott B. Parasitic copepods from pelagic sharks in Western Australia // Bulletin of Marine Science. — 1999. — Vol. 65, № 3. — P. 715—724.
- Jensen C., Schwartz F. J. Atlantic Ocean occurrences of the sea lamprey, Petromyzon marinus (Petromyzontiformes: Petromyzontidae), parasitizing sandbar, Carcharhinus plumbeus, and dusky, C. obscurus (Carcharhiniformes: Carcharhinidae), sharks off North and South Carolina // Brimleyana. — 1994. — Vol. 21. — P. 69—76.
- Gelsleichter J., Musick J. A., Nichols S. Food habits of the smooth dogfish, Mustelus canis, dusky shark, Carcharhinus obscurus, Atlantic sharpnose shark, Rhizoprionodon terraenovae, and the sand tiger, Carcharias taurus, from the northwest Atlantic Ocean // Environmental Biology of Fishes. — 1999. — Vol. 54, № 2. — P. 205—217.
- Hussey N. E., Cocks D. T., Dudley S. F. J., McCarthy I. D., Wintner S. P. The condition conundrum: application of multiple condition indices to the dusky shark Carcharhinus obscurus // Marine Ecology Progress Series. — 2009. — Vol. 380. — P. 199—212. — doi:10.3354/meps07918.
- Marin A. The Power of Shark Bites. ReefQuest Centre for Shark Research. Дата обращения: 18 мая 2015.
- Gubanov E. P. Morphological characteristics of the requiem shark, Carcharinus obscurus, of the Indian Ocean // Journal of Ichthyology. — 1988. — Vol. 28, № (6). — P. 68—73.
- Simpfendorfer C. A., Goodreid A., McAuley R. B. Diet of three commercially important shark species from Western Australian waters // Marine and Freshwater Research. — 2001. — Vol. 52, № (7). — doi:10.1071/MF01017.
- Smale M. J. Occurrence and feeding of three shark species, Carcharhinus brachyurus, C. obscurus and Sphyrna zygaena, on the eastern Cape coast of South Africa // South African Journal of Marine Science. — 1991. — P. 31—42.
- Cockcroft V. G., Cliff G., Ross G. J. B. Shark predation on Indian Ocean bottlenose dolphins Tursiops truncatus off Natal, South Africa // South African Journal of Zoology. — 1989. — Vol. 24, № (4). — P. 305—310.
- Pratt H. L. (Jr.). The storage of spermatozoa in the oviducal glands of western North Atlantic sharks. — 1993. — Vol. 38. — P. 139—149. — doi:10.1007/BF00842910.
- Fowler S. L., Cavanagh R. D., Camhi M., Burgess G. H., Cailliet G. M., Fordham S. V., Simpfendorfer C. A., Musick J. A. Sharks, Rays and Chimaeras: The Status of the Chondrichthyan Fishes. — International Union for Conservation of Nature and Natural Resources, 2005. — P. 106—109, 287—288. — ISBN 2831707005.
- Natanson L. J., Kohler N. E. A preliminary estimate of age and growth of the dusky shark Carcharhinus obscurus from the South-West Indian Ocean, with comparisons to the western North Atlantic population // South African Journal of Marine Science. — 1996. — Vol. 17. — P. 217—224. — doi:10.2989/025776196784158572.
- White W. T. Catch composition and reproductive biology of whaler sharks (Carcharhiniformes: Carcharhinidae) caught by fisheries in Indonesia // Journal of Fish Biology. — 2007. — Vol. 71, № (5). — P. 1512—1540. — doi:10.1111/j.1095-8649.2007.01623.x..
- Simpfendorfer C. A., McAuley R. B., Chidlow J., Unsworth P. Validated age and growth of the dusky shark, Carcharhinus obscurus, from Western Australian waters // Marine and Freshwater Research. — 2002. — Vol. 53, № (2). — P. 567—573. — doi:10.1071/MF01131.
- Simpfendorfer C. A. Growth rates of juvenile dusky sharks, Carcharhinus obscurus (Lesueur, 1818), from southwestern Australia estimated from tag-recapture data // Fishery Bulletin. — 2000. — Vol. 98, № (4). — P. 811—822. Архивировано 31 декабря 2016 года.
- ISAF Statistics on Attacking Species of Shark. Florida Museum of Natural History Ichthyology Department. Дата обращения: 23 мая 2015.
- Species of Concern: Dusky Shark. (Jan. 6, 2009). NMFS Office of Protected Resources. Retrieved on May 18, 2009.
- Pank M., Stanhope M., Natanson L., Kohler N., Shivji M. Rapid and Simultaneous Identification of Body Parts from the Morphologically Similar Sharks Carcharhinus obscurus and Carcharhinus plumbeus (Carcharhinidae) Using Multiplex PCR // Marine Biotechnology. — 2001. — Vol. 3, № (3). — P. 231—240. — doi:10.1007/s101260000071. — PMID 14961360.
Литература
- Ebert D. A. Sharks, Rays, and Chimaeras of California. — California: University of California Press, 2003. — P. 149—152. — ISBN 0520234847.
- Compagno L. J. V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. — Rome: Food and Agricultural Organization, 1984. — P. 489—491. — ISBN 92-5-101384-5.
- Fowler S. L., Cavanagh R. D., Camhi M., Burgess G. H., Cailliet G. M., Fordham S. V., Simpfendorfer C. A., Musick J. A. Sharks, Rays and Chimaeras: The Status of the Chondrichthyan Fishes. — International Union for Conservation of Nature and Natural Resources, 2005. — P. 106—109, 287—288. — ISBN 2831707005.
