Морские млекопитающие
Морские млекопитающие — сборная группа водных и полуводных млекопитающих, чья жизнь полностью или существенную часть времени проходит в морской среде. В эту категорию входят представители различных систематических групп млекопитающих: сирены, китообразные, ластоногие — ушастые тюлени, настоящие тюлени, моржовые. Помимо этих животных к морским млекопитающим также относят единичных представителей семейств куньих (калан и морская выдра) и медвежьих (белый медведь)[1][2]. Всего к морским млекопитающим относится около 130 ныне живущих видов[3][4], составляющих примерно 2 % от общего числа современных млекопитающих[4].
| Полифилетическая группа животных | |||
|---|---|---|---|
 | |||
| Название | |||
| Морские млекопитающие | |||
| Статус названия | |||
| не определён | |||
| Родительский таксон | |||
| Класс Млекопитающие | |||
| Представители | |||
| |||
Происхождение
Морские млекопитающие — звери, произошедшие от сухопутных животных, которые вторично связали свою жизнь на определённом этапе эволюционного развития с морской водной стихией. Возникновение сирен и китообразных датируется эоценом, их предками были соответственно древние афротерии и парнокопытные[5]. Дальнейшее разделение китообразных на усатых и зубатых китов произошло на границе эоцена и олигоцена. К концу эоцена как китообразные, так и сирены эволюционировали в полностью водные организмы с атрофированными задними конечностями и специализированным хвостом, питающиеся, слышащие и размножающиеся в воде. Эта эволюция проходила в эпоху, когда моря были мелкими и тёплыми, богатыми пищей. У сирен, наряду с хоботными, был ещё один родственный отряд, также относящийся к кладе Tethyteria — десмостилии. Эти животные, также относимые к морским млекопитающим, существовали с начала олигоцена до конца миоцена и известны по находкам в северной части Тихого океана. Изотопный анализ углерода и кислорода в тканях зубов показал, что рацион десмостилий напоминал рацион современных ламантинов и состоял из морских и пресноводных водорослей[6]. Ластоногие эволюционировали примерно на 20 млн лет позже, чем китообразные и сирены, от древних хищных[5]. Наиболее ранний представитель этой группы, эналиаркт, известен по позднеолигоценовым отложениям Орегона. Формы, более близкие к современным ластоногим — Pteronarctos и Pacificotaria — развились в начале и середине миоцена; у них уже наличествует уникальное для современных ластоногих устройство окологлазничных костей[7]. Талассокнусы (Thalassocnus) — род ведших полуводный образ жизни ленивцев — существовали в плиоцене в Южной Америке. Наконец, представители куньих и медвежьих среди морских млекопитающих появились лишь относительно недавно и практически неизвестны по палеонтологическим находкам. Опознание ископаемых морских млекопитающих, в частности медведей, может быть затруднено наличием близких, но не морских видов (определить ископаемые находки с точностью до вида часто невозможно)[6]. Род Enhydra, к которому относится современный калан, по-видимому, сформировался в начале плейстоцена, 1—3 млн лет назад; предполагается, что от рода выдры до него отпочковывались ещё две ветви, порождавшие морских выдр, но до настоящего времени их представители не дожили. Существуют разные оценки, когда белый медведь отделился от бурого (Ursus arctos, с которым до настоящего времени возможна успешная гибридизация) — от 4-5 миллионов до 600 тысяч лет назад, а анализ генома указывает на ещё более недавнее время — около 160 тысяч лет назад. Палеонтологические находки позволяют достоверно утверждать, что белый медведь адаптировался к арктическим холодам и морской диете не позднее, чем 110 тысяч лет назад[8].
Морфологические сходства внутри этой полифилетической группы объясняются конвергентной и параллельной эволюцией[5]. Адаптационные изменения в анатомии морских млекопитающих, облегчающие водный или полуводный образ жизни (см. Адаптация к морскому образу жизни), включают сглаживание тела и приобретение им веретенообразной формы, потерю задних конечностей (как у китообразных и сирен) или их модификацию для движения в воде (как у ластоногих). У каланов и белых медведей морфологические изменения менее выражены, и они всё ещё сильно напоминают внешне своих родичей, ведущих сухопутный образ жизни[1]. Скелеты десмостилий тоже не обладают характерными для водных организмов особенностями и их причисление к морским млекопитающим практически полностью основывается на том факте, что их останки находят только в морских отложениях[6].
Систематика морских млекопитающих
(по книге Marine Mammals of the World, 1993)[9]
Выделяется около 130 современных вида морских млекопитающих, подразделяемых на пять групп, перечисленных ниже[3][4][10].
- Отряд Sirenia: сирены
- семейство Trichechidae: ламантины (3 вида)
- семейство Dugongidae: дюгони (2 вида, в т. ч. 1 вымерший)
- Отряд Cetartiodactyla: китопарнокопытные
- инфраотряд Cetacea: китообразные
- парвотряд Mysticeti: усатые киты (15—16 видов)
- парвотряд Odontoceti: зубатые киты
(73—75 морских видов)
- инфраотряд Cetacea: китообразные
- Отряд Carnivora: хищные
- парвотряд Pinnipedia: ластоногие[11]
- семейство Phocidae: настоящие тюлени
(19 видов, в т. ч. 1 вымерший) - семейство Otariidae: ушастые тюлени
(15—16 видов, в т. ч. 1 вымерший) - семейство Odobenidae: моржовые (1 вид)
- семейство Phocidae: настоящие тюлени
- семейство Mustelidae: куньи
- калан (Enhydra lutris)
- морская (кошачья) выдра (Lontra felina)
- семейство Ursidae: медвежьи
- белый медведь (Ursus maritimus)
- парвотряд Pinnipedia: ластоногие[11]
Ранее ластоногих и китообразных нередко рассматривали в качестве самостоятельных отрядов. Помимо ныне существующих клад, к морским млекопитающим относят и две вымерших:
- отряд Desmostylia (десмостилии, ранний олигоцен — поздний миоцен)[6]
- и род Thalassocnus (талассокнус, плиоцен)[6].
На основании морфологического сходства устройства черепа и зубного аппарата с каланами и кошачьими выдрами к морским млекопитающим могут также относить ископаемый род хищных Kolponomos (колпономос), известный по миоценовым находкам и предположительно родственный ластоногим, но внешне напоминавший медведей[12]. Включая описанные ископаемые, число известных видов китообразных к концу первого десятилетия XXI века превышало 240, сирен — 30 и ластоногих — 60[6].
 Американский ламантин (Trichechus manatus), из отряда Сирены
Американский ламантин (Trichechus manatus), из отряда Сирены Горбач (Megaptera novaeangliae), из инфраотряда Китообразные
Горбач (Megaptera novaeangliae), из инфраотряда Китообразные Калифорнийский морской лев, из семейства Ушастые тюлени
Калифорнийский морской лев, из семейства Ушастые тюлени
- Белый медведь (Ursus maritimus), из семейства Медвежьи
Зафиксировано несколько десятков случаев успешной гибридизации между разными видами китообразных и ластоногих, как в естественной среде обитания, так и в неволе. Эти случаи описаны преимущественно на морфологическом уровне, хотя в последнее время началось и применение молекулярных техник изучения. в частности показавшие, что от 17 до 30 процентов популяции морских котиков на острове Маккуори (Австралия) представляли собой гибриды трёх разных видов — субтропического, кергеленского и новозеландского. Среди китообразных родившиеся в неволе гибриды афалин (с малой косаткой и длиннорылой белобочкой) в свою очередь успешно приносили потомство в первом поколении. Учёными наблюдались случаи межвидового спаривания (между длинномордым и обыкновенным тюленем, южным морским львом и южноамериканским морским котиком и т. д.)[13]
Адаптация к морскому образу жизни
Физиология и модели поведения морских млекопитающих претерпели изменения, позволяющие им лучше приспособиться к жизни в море. Для давно возникших таксонов морских млекопитающих характерны изменения в форме тела, улучшающие её гидродинамичность. У китообразных, сирен и ластоногих тело за время эволюции приобрело обтекаемую форму. Задние конечности у первых двух групп атрофировались, а у третьей поменяли форму так, чтобы обеспечивать более эффективное передвижение в водной среде[1].
Важной характеристикой жизни в водной среде является способность к нырянию. Морские животные могут нырять за пищей или спасаться под водой от хищников, ныряние может быть способом увеличить скорость движения в воде или служить сохранению энергии за счёт сокращения метаболизма. В целом морские млекопитающие превосходят других представителей этого класса в способности к нырянию, хотя у отдельных видов она развита хуже (в частности, у ушастых тюленей и сирен максимальная продолжительность задержки дыхания как правило составляет менее 10 минут). Зафиксированные рекорды продолжительности и глубины погружения к середине 2010-х годов принадлежали кювьерову клюворылу из подотряда зубатых китов — 137,5 минуты и 2991 м соответственно; долгое пребывание под водой и большую глубину погружения демонстрируют также кашалоты, морские слоны, киты-бутылконосы и гринды[14].
Поскольку млекопитающие не могут дышать под водой, это быстро приводит к отрицательным физиологическим эффектам — истощению кислорода и росту концентрации углекислого газа в крови, накоплению продуктов молочной кислоты в тканях, и организм морских млекопитающих адаптировался к этим изменениям. Ещё один фактор, требующий адаптации — повышенное давление на глубине. У ластоногих кровяное давление в мозгу регулируется за счёт варьирующегося диаметра аорты, причём размер так называемой аортной луковицы (англ. aortic bulb) напрямую коррелирует с глубиной погружения у разных видов. Схожая особенность анатомии зафиксирована и у некоторых китообразных. Для кровеносной системы морских млекопитающих характерны также плотные массы свёрнутых спиралями артерий и вен, известные как retia mirabilia, расположенные вдоль внутренней задней стенки грудной полости, а также в конечностях и на периферии тела. Эти скопления сосудов служат резервуарами кислорода и особенно развиты у такого умелого ныряльщика, как кашалот[14]. Запасы кислорода также могут создаваться в мышцах, благодаря связывающему кислород белку миоглобину — это характерно не только для морских млекопитающих, но и для других ныряющих животных, включая пресноводных выдр, бобров и ондатр[15]. Собственные запасы кислорода в мышцах позволяют исключить их из цикла снабжения кислородом через кровь, сохраняя его для мозга и других чувствительных органов. Новую порцию кислорода мышцы получают лишь после возобновления дыхания. Наконец, расход кислорода уменьшается благодаря замедлению ритма сердцебиения (брадикардии)[16].
Водный образ жизни требует хорошего энергетического баланса — водный организм должен или наиболее эффективно получать и накапливать энергию, либо экономно её расходовать. Второе может достигаться за счёт сезонных миграций, вариаций основного обмена веществ или терморегуляции. Поскольку для многих морских млекопитающих характерны длительные периоды голодания, их организмы приспособились к потере значительной части массы тела — так, у северных морских слонов она может достигать 42 %, но бо́льшая часть приходится на жировую ткань, почти не затрагивая другие ткани. Сохранение тепла у морских млекопитающих может осуществляться с помощью как меха, так и жировой прослойки. Меховой покров эффективен при условии, что между ворсинками содержится воздух, служащий теплоизоляцией, и поэтому мех нуждается в уходе и расчёсывании — так, подсчитано, что калан проводит 16 % своего времени в уходе за мехом. Жир требует не ухода, а усиленного питания, и в голодное время, когда он используется организмом для поддержания энергии, его эффективность как теплоизолятора падает. Чтобы избежать прилива крови к коже и лишней теплопотери, тюлени, выходящие на берег в голодное время, сбиваются вместе, прижимаясь друг к другу и таким образом уменьшая площадь открытой поверхности тела. Лучшая утилизация питательных веществ, в частности, достигается за счёт удлинения кишечника — у всех хищных морских млекопитающих, вне зависимости от происхождения, тонкая кишка длиннее, чем у сухопутных млекопитающих сопоставимых габаритов[17].
Поскольку распространение звука в воде и воздухе различно, слух китообразных, сирен и ластоногих адаптирован к водной среде (слух белого медведя и морских выдр, по-видимому, всё ещё аналогичен слуху наземных животных, а в воде они предположительно полагаются на акустическую проводимость кости). У китообразных отсутствуют ушные раковины, внешний слуховой проход истончён, частично блокирован и в целом нефункционален. Звук поступает в ухо напрямую через мягкие ткани головы. Кроме того, видом адаптации к водной среде является способность китообразных к эхолокации[18].
Распространение
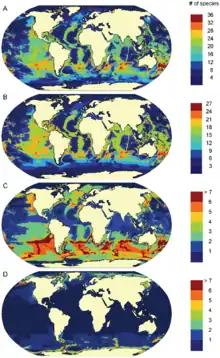
Географическое распространение морских млекопитающих по морям и океанам Земли не равномерно и не случайно. Ареал конкретных видов может определяться глубиной, температурным и океанографическим режимом, и в частности морскими течениями, а для пагофильных видов (среда обитания которых связана со льдами) — формированием и движением морского льда[9]. Распространение морских млекопитающих напрямую связано с распространением основных источников пищи — таких как веслоногие ракообразные для северного гладкого кита или подводные «луга» для ламантинов — а также естественных врагов, что особенно существенно для более мелких видов[20].
С точки зрения заселённых глубин и близости к береговой линии морские млекопитающие могут быть разделены на прибрежных (обитающих в эстуариях и прибрежных водах), неритических (населяющих воды континентального шельфа) и океанических (обитателей открытого моря за пределами подводных окраин материков). Примерами первой категории могут служить каланы, дюгони и бутылконосые дельфины, второй — серые киты и калифорнийские морские львы, третьей — кашалоты и клюворыловые киты. Для кашалотов и морских слонов способность глубоко нырять, добывая пищу на недоступных для конкурирующих видов глубинах, обусловила их проживание в районе подводных каньонов и впадин, тогда как усатые киты в целом, с их ориентацией на океанский планктон как основной источник питания, предпочитают меньшие глубины. В то же время это разделение достаточно условно, так как многие виды морских млекопитающих могут обитать на разных глубинах и расстояниях от берега. Несмотря на название, морские млекопитающие проникают и в пресноводные водоёмы. Естественной средой обитания для речных дельфинов (гангского, амазонского, китайского, а также ла-платского) являются реки, а некоторые настоящие тюлени (каспийская, байкальская, сайменская нерпы и тюлень Унгавы) обитают в пресных озёрах в глубине континента[20].
Другой возможный критерий различения морских млекопитающих — основные географические широты обитания: существуют тропические и субтропические виды, обитатели умеренных широт и жители Арктики и Антарктики. Некоторые виды строго привязаны только к одной из климатических зон — так, белый медведь, нарвал, белуха и гренландский кит обитают исключительно в арктических водах, а ареал сирен ограничен тропическими и субтропическими, — но другие свободно перемещаются между ними. Примером могут служить миграции некоторых усатых китов между холодными полярными и тёплыми тропическими водами или распространение косаток во всех климатических зонах[20]. Другие примеры космополитичных видов — серый дельфин и обыкновенный тюлень. В некоторых случаях близкородственные виды распространены в умеренных водах по обе стороны тропической зоны, без пересечений ареала. Таковы калифорнийская морская свинья, северный плавун и северный морской слон в Северном полушарии и близкие к ним аргентинская морская свинья, южный плавун и южный морской слон — в Южном. Такая специфика распространения, по-видимому, является результатом аллопатрического видообразования — возможно, в процессе плейстоценовых похолоданий, когда вид, изначально зародившийся по одну сторону экватора, распространялся во второе полушарие, а затем популяции разделялись при новом глобальном потеплении[21].
Взаимодействие с человеком
Человеческая деятельность (охота, хищническое рыболовство, ведущее к истощению источников питания, и загрязнение окружающей среды) оказывает важное влияние на морских млекопитающих. В особенности это касается видов, чьё существование хотя бы частично проходит на суше — их естественные условия обитания трансформируются в связи с расширением районов, заселяемых людьми. Примером могут служить тюлени-монахи. Помимо того, что их среда обитания пострадала особенно сильно, они были также объектом коммерческого промысла, и в результате человеческой деятельности один из видов этого рода — карибский тюлень-монах — полностью вымер, а другой — средиземноморский тюлень-монах, раньше предпочитавший открытые пляжи, — к настоящему времени выходит на берег в труднодоступных пещерах, в том числе с подводными входами. Изменения температуры прибрежных вод, вызванные человеческой деятельностью, повлияли на сезонные миграции американского ламантина: популяции, ранее откочёвывавшие зимой в более тёплые широты, остаются севернее, но держатся ближе к тепловым электростанциям, сбрасывающим горячую воду в море. Глобальное потепление, связываемое с человеческой деятельностью, сократило площадь и время существования сезонных морских льдов, от которых зависит выживание белых медведей. Этот же фактор влечёт за собой сокращение популяций антарктического криля в Южной Атлантике, критичного для многих видов усатых китов, а также морских котиков, тюленей-крабоедов и морских леопардов. В то же время освобождение ото льда новых участков моря облегчает доступ к новым видам добычи для косаток, чья диета не специализирована[20].
Будучи в основном крупными животными, морские млекопитающие на протяжении долгого времени привлекали к себе внимание человека как объекты охоты. В отличие от крупных наземных млекопитающих, для которых подобное внимание закончилось полным исчезновением (мамонты и североамериканские дикие лошади в позднем плейстоцене и большие нелетающие птицы в исторический период), труднодоступность мест обитания морских млекопитающих служила им защитой. Важным фактором были и огромные пространства, на которых приходилось вести их поиск. Только на протяжении нескольких последних столетий развитие технологий обеспечило эффективный поиск и добычу морских млекопитающих в промышленных масштабах, быстро приведя многие виды на грань исчезновения. Однако в результате сокращения популяций, делающего добычу нерентабельной, и усиления природоохранных движений истребление в большинстве случаев было остановлено. Тем не менее ряд видов морских млекопитающих в результате человеческой деятельности исчез полностью. Помимо упомянутого выше карибского тюленя-монаха, были истреблены стеллерова корова, японский морской лев, китайский речной дельфин и атлантический серый кит — Eschrichtius robustus gibbosus (почти полностью истреблена также популяция серых китов на севере Тихого океана между Охотским морем и югом Корейского полуострова)[22]. В 1800-е годы была окончательно истреблена охотниками за мехами морская норка (Neogale macrodon) — единственный, помимо калана и кошачьей выдры, представитель куньих среди морских млекопитающих[23]. Угрожаемым остаётся положение таких видов, как средиземноморский тюлень-монах (популяция менее 1000 особей) и калифорнийская морская свинья — представители этого вида гибнут не в результате намеренной охоты, а запутываясь в рыболовных сетях. Популяции крупных полосатиковых китов, северного гладкого кита и серого кита восстанавливаются очень медленно, и угроза этим видам не миновала[22]. В целом, по разным оценкам, под угрозой к концу первого десятилетия XXI века находилось существование от 23 % до 61 % известных видов морских млекопитающих[24].
См. также
Примечания
- Marine Mammals of the World, 1993, p. 1.
- Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1989. — Т. 7 : Млекопитающие / под ред. В. Е. Соколова. — С. 558. — 558 с. : ил. — ISBN 5-09-001434-5.
- List of Marine Mammal Species and Subspecies (англ.). Society for Marine Mammalogy. Дата обращения: 13 сентября 2021. Архивировано 21 июня 2021 года.
- Higher Taxonomy (англ.). ASM Mammal Diversity Database. Дата обращения: 13 сентября 2021.
- Foote, A. D., et al. Convergent evolution of the genomes of marine mammals // Nature Genetics. — 2015. — Vol. 47. — P. 272—275. — doi:10.1038/ng.3198.
- Uhen, M. D. Evolution of Marine Mammals: Back to the Sea After 300 Million Years // The Anatomical Record. — 2007. — Vol. 290. — P. 514—522. — doi:10.1002/ar.20545.
- Berta, Sumich & Kovacs, 2015, pp. 41—43.
- Berta, Sumich & Kovacs, 2015, pp. 119—123.
- Marine Mammals of the World, 1993, p. 2.
- Полная иллюстрированная энциклопедия. «Млекопитающие» Кн. 2 = The New Encyclopedia of Mammals / под ред. Д. Макдональда. — М.: Омега, 2007. — С. 504. — 3000 экз. — ISBN 978-5-465-01346-8.
- Burgin C. J., Widness J., Upham N. S. Introduction to Illustrated Checklist of the Mammals of the World // Illustrated Checklist of the Mammals of the World (англ.) / ed. by Burgin C. J., Wilson D. E., Mittermeier R. A., Rylands A. B., Lacher T. E., Sechrest W. — Lynx Edicions, 2020. — P. 27. — ISBN 978-84-16728-36-7.
- Berta, Sumich & Kovacs, 2015, pp. 115—116.
- Bérubé, M. Hybridism // Encyclopedia of Marine Mammals / W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Eds.). — 2nd Edition. — Rome: Academic Press, 2009. — P. 588—592. — ISBN 978-0-12-373553-9.
- Berta, Sumich & Kovacs, 2015, pp. 299—301.
- Lee, J. J. How Diving Mammals Stay Underwater for So Long (англ.). National Geographic (15 июня 2013). Дата обращения: 16 апреля 2019.
- Blix, A. S. Adaptations to deep and prolonged diving in phocid seals (англ.) // The Journal of Experimental Biology. — The Company of Biologists, 2018. — Vol. 221. — doi:10.1242/jeb.182972.
- Costa, D. P. Energetics // Encyclopedia of Marine Mammals / W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Eds.). — 2nd Edition. — Rome: Academic Press, 2009. — P. 383—391. — ISBN 978-0-12-373553-9.
- Nummela, S. Hearing // Encyclopedia of Marine Mammals / W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Eds.). — 2nd Edition. — Rome: Academic Press, 2009. — P. 553—562. — ISBN 978-0-12-373553-9.
- Kaschner, K., Tittensor, D. P., Ready, J., Gerrodette, T., and Worm, B. Current and future patterns of global marine mammal biodiversity (англ.) // PLOS One. — Public Library of Science, 2011. — Vol. 6, no. 5. — P. e19653. — doi:10.1371/journal.pone.0019653.
- Forcada, J. Distribution // Encyclopedia of Marine Mammals / W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Eds.). — 2nd Edition. — Rome: Academic Press, 2009. — P. 316—321. — ISBN 978-0-12-373553-9.
- Berta, Sumich & Kovacs, 2015, pp. 144—145.
- Duffield, D. A. Extinctions, Specific // Encyclopedia of Marine Mammals / W. F. Perrin, B. Würsig, and J. G. M. Thewissen (Eds.). — 2nd Edition. — Rome: Academic Press, 2009. — P. 402—404. — ISBN 978-0-12-373553-9.
- Berta, Sumich & Kovacs, 2015, p. 119.
- Schipper, J., et al. The Status of the World’s Land and Marine Mammals: Diversity, Threat, and Knowledge (англ.) // Science. — 2008. — Vol. 322. — P. 227. — doi:10.1126/science.1165115.
Литература
- Berta, A., Sumich, J. L., and Kovacs, K. M. Marine Mammals: Evolutionary Biology. — 3nd Ed. — Academic Press, 2015. — ISBN 978-0-12-397002-2.
- Jefferson, T. A., Leatherwood, S., and Webber, M. A. Marine Mammals of the World. — Rome: United Nations Environment Programme, Food and Agriculture Organization of the United Nations, 1993. — ISBN 92-5-103292-0.
- Perrin, W. F., Würsig, B., and Thewissen, J. G. M. Encyclopedia of Marine Mammals. — 2nd Edition. — Rome: Academic Press, 2009. — ISBN 978-0-12-373553-9.