Непарнокопытные
Непарнокопытные, или непарнопалые (лат. Perissodactyla), — отряд крупных наземных плацентарных млекопитающих. В отличие от парнокопытных (Artiodactyla) для них характерно нечётное число пальцев, образующих копыта. Отряд содержит три современных семейства — лошадиных (Equidae), носороговых (Rhinocerotidae) и тапировых (Tapiridae), которые вместе насчитывают 18 видов. Родство этих внешне весьма отличающихся друг от друга семейств впервые установил зоолог Ричард Оуэн в XIX веке, придумав также название «непарнокопытные» (англ. odd-toed ungulates).
| Непарнокопытные | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Разнообразие непарнокопытных. Слева: бурчелловы зебры; Справа: индийский носорог, равнинный тапир. | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Класс: Подкласс: Клада: Инфракласс: Надотряд: Клада: Грандотряд: Отряд: Непарнокопытные |
||||||||||||
| Международное научное название | ||||||||||||
| Perissodactyla Owen, 1848 | ||||||||||||
| Семейства | ||||||||||||
|
||||||||||||
| ||||||||||||
Внешний вид
Общие сведения
Из-за приспособления к разным средам обитания у непарнокопытных со временем возникли значительные различия в строении тела. Общие черты существуют в строении конечностей и зубов. Все современные и подавляющее большинство вымерших видов — довольно крупные животные. Представители семейства носорогов занимают второе место (после слонов) по величине среди наземных млекопитающих. Вымерший индрикотерий, безрогий родственник носорогов из эпохи олигоцена, даже считается самым крупным наземным млекопитающим всех времён. Однако некоторые ранние представители отряда, такие как пралошадка гиракотерий, были небольшими, достигая высоты в холке лишь 20 см. За исключением некоторых искусственно выведенных карликовых пород лошадей, современные непарнокопытные достигают длины тела от 180 до 420 см, а их масса составляет 150–3500 кг. Варьирует и их волосяной покров. В то время как у носорогов он редок и компенсируется толстым эпидермисом, у тапиров и лошадей более густая короткая шерсть. Большинство видов окрашены в серый или коричневый цвета. Для зебр, однако, характерны чёрно-белые вертикальные полосы. Похожий горизонтальный рисунок можно увидеть у детёнышей тапиров.
Конечности
Основная нагрузка передних и задних конечностей приходится на центр, из-за чего самым длинным пальцем у всех видов является третий. Остальные пальцы в той или иной степени атрофировались, менее всего у тапиров. Эти животные из-за мягких почв в своей среде обитания имеют по четыре пальца на передних ногах и по три на задних. У лошадей атрофированность боковых пальцев зашла наиболее далеко, у них имеется всего один палец. Конечности завершаются копытами, которые, однако, лишь у лошадей покрывают пальцы полностью. У носорогов и тапиров копытами покрыта только передняя часть ноги, а нижняя сторона мягкая. У носорогов ступня довольно мягкая.
В конечностях уменьшены бедренная и малоберцовая кости, у лошадей они срослись с большеберцовой костью. Исключительной чертой непарнокопытных является особый сустав между таранной и ладьевидной костью, который сильно ограничивает их подвижность.
Череп и зубы

У непарнокопытных, как правило, продолговатая голова, отличающаяся в первую очередь длинной верхней челюстью. Разные формы челюстного аппарата отдельных семейств происходят от различного строения кости Praemaxillare. На слёзной кости имеется нарост, вдающийся в глазницу. Уникален широкий контакт между слёзной и носовой костью. У видов, питающихся преимущественно травой, челюсти особо массивные. Челюстной сустав расположен довольно глубоко, а нижняя челюсть увеличена.
У носорогов по одному или двум рогам, которые, в отличие от рогов парнокопытных, состоят не из кости, а из кератина.
Количество и строение зубов варьирует в зависимости от питания. Резцы и клыки могут быть очень малыми либо отсутствовать вовсе, как у африканских видов носорогов. У лошадей клыки имеются обычно только у самцов. Из-за продолговатой верхней челюсти между передними и боковыми зубами существует пространство, называемое диастема. Премоляры развиты обычно как моляры. Поверхность и высота моляров сильно зависит от того, питается ли животное больше мягкой листвой или жёсткой травой. На каждой половине челюсти имеются от трёх до четырёх премоляров и по три моляра. Зубная формула выглядит следующим образом: I 0-3/0-3 °C 0-1/0-1 P 3-4/3-4 M 3/3.
Внутренняя анатомия
Строение пищеварительного тракта непарнокопытных сильно отличается от его строения у парнокопытных, которые также являются растительноядными животными. Непарнокопытные, как и грызуны, переваривают пищу главным образом в толстой кишке. В отличие от парнокопытных, желудок у представителей непарнокопытных однокамерный и построен относительно просто. Пищеварение проходит в очень крупном аппендиксе, который у лошадей вмещает до 90 л, и в двухпетельной ободочной кишке (Colon). Кишечник очень длинный и может достигать у лошадей 26 м. Использование питательности пищи довольно низкое. Это, вероятно, является причиной того, что не существует малых непарнокопытных, так как у крупных животных потребность в пище на кг массы ниже, а соотношение поверхности к объёму выгоднее в плане теплосберегаемости. В области мочеполовой системы самки охарактеризованы «двухроговой маткой» (Uterus bicornis). Яичники расположены у носорогов и тапиров в своеобразном «кармане» брюшины (Peritoneum), у лошадей они находятся частично вне его. У самцов тапиров и носорогов яички находятся внутри туловища, мошонка есть только у лошадиных.
Распространение
Современный ареал непарнокопытных состоит лишь из небольшой части исконного ареала, охватывавшего почти всю сушу Земли. Дикие представители непарнокопытных сегодня обитают в Центральной и Южной Америке, в Восточной и Южной Африке, а также в Средней, Южной и Юго-Восточной Азии. В Северной Америке они вымерли около 10 тысяч лет назад, в Европе — в XIX веке, когда был убит последний тарпан. Охота и сокращение жизненного пространства привели к тому, что и сегодняшние дикие виды встречаются лишь в разрозненных реликтовых популяциях. В отличие от них, домашние лошади и домашние ослы обрели широкое распространение по всему миру и встречаются в наши дни в том числе и в регионах, где изначально не существовало непарнокопытных, например в Австралии.
Поведение и питание
В зависимости от среды обитания различные виды непарнокопытных ведут различный образ жизни. Они преимущественно активны в сумеречное или ночное время. Тапиры ведут одиночный образ жизни и обитают главным образом в тропических и иных лесах. Носороги живут тоже поодиночке и встречаются в Африке скорее в засушливых саваннах, а в Азии во влажных болотных и лесных местностях. Лошадиные населяют открытые местности, такие как саванны, степи и полупустыни, держась группами. Непарнокопытные — исключительно растительноядные, питающиеся в разной степени травами, листьями и другими частями растений.
Размножение

Непарнокопытные отличаются длинным сроком беременности и немногочисленным потомством. Как правило, на свет рождается по одному детёнышу. Продолжительность беременности составляет от 330 до 500 дней и наиболее велика у носорогов. Новорожденные непарнокопытные уже спустя несколько часов после рождения следуют за матерью. Исключение составляют лишь детёныши тапиров, проводящие первые дни жизни в защищённом месте. Потомство более чем год вскармливается молоком и достигает половую зрелость в возрасте от 2 до 8 лет. Непарнокопытные живут довольно долго. В неволе их возраст доходил до 50 лет у носорогов.
Эволюционная история и систематика
Внутренняя систематика



Только 3 из примерно 12 семейств непарнокопытных дожили до наших дней. Всего насчитывается 18 современных (из не менее 500 существовавших) видов в шести родах. Все они перечислены ниже (слово «вид» не указывается).
- Семейство лошадиные (Equidae)
- Род лошади, или настоящие лошади (Equus)
- Подрод зебры
- Бурчеллова зебра (Equus quagga) (ранее Equus burchelli)
- Горная зебра (Equus zebra)
- Зебра Греви (Equus grevyi)
- Дикая лошадь (Equus ferus)
- Домашняя лошадь (Equus caballus)
- Кианг (Equus kiang)
- Кулан (Equus hemionus)
- Дикий осёл (Equus africanus)
- Домашний осёл (Equus asinus)
- Подрод зебры
- Род лошади, или настоящие лошади (Equus)
- Семейство тапировые (Tapiridae)
- Род тапиры (Tapirus)
- Горный тапир (Tapirus pinchaque)
- Равниный, или бразильский тапир (Tapirus terrestris)
- Центральноамериканский тапир, или тапир Бэйрда (Tapirus bairdii)
- Чепрачный, или малайский тапир (Tapirus indicus)
- Род тапиры (Tapirus)
- Семейство носороговые (Rhinocerotidae)
- Род Dicerorhinus
- Суматранский носорог (Dicerorhinus sumatrensis)
- Род Rhinoceros
- Индийский носорог (Rhinoceros unicornis)
- Яванский носорог (Rhinoceros sondaicus)
- Род Diceros
- Чёрный носорог (Diceros bicornis)
- Род Ceratotherium
- Белый носорог (Ceratotherium simum)
- Род Dicerorhinus
Ранее кваггу (истреблена) и бурчеллову зебру выделяли в два отдельных вида зебр. Теперь они считаются подвидами равнинной зебры (Equus quagga quagga и Equus quagga burchelli соответственно). Семейства тапировых и носороговых иногда объединяют в подотряд Ceratomorpha.
Внешняя систематика
Непарнокопытных традиционно объединяли с отрядами парнокопытных, даманов и хоботных в расширенную группу копытных (Ungulata). В частности, предполагалось наиболее близкое родство с даманами, что аргументировалось общим строением уха и позицией сонной артерии. Однако недавние молекулярно-генетические исследования вызвали большие сомнения в родстве копытных между собой. Вероятно, что копытные являются полифилетической группой, то есть их сходство обусловлено конвергентной эволюцией, а не общим происхождением. Хоботных и даманов сегодня относят к надотряду афротерий, которых более не рассматривают в качестве родичей непарнокопытных. Последние состоят в надотряде лавразиотериий, который происходит с древнего континента Лавразия. Результаты молекулярно-генетических исследований позволяют предполагать, что сестринским таксоном являются Ferae, который объединяет отряды хищных и панголинов. Вместе c Ferae непарнокопытные образуют таксон Zooamata.

Возможная кладограмма, иллюстрирующая положение непарнокопытных среди лавразиотерий, выглядит следующим образом[3]:
Лавразиотерии ├─ Насекомоядные (Eulipotyphla) └─ Scrotifera ├─ Рукокрылые (Chiroptera) └─ Fereuungulata ├─ Китопарнокопытные (парнокопытные и киты) └─ Zooamata ├─ Непарнокопытные (Perissodactyla) └─ Ferae ├─ Панголины (Pholidota) └─ Хищные (Carnivora)
Потенциальной сестринской группой Zoomata считаются на сегодняшний день китопарнокопытные (Cetartiodactyla). Вместе они образуют таксон Fereuungulata. В 2006 году японскими учёными также предполагалось общее родство Zooamata с рукокрылыми. Их общий таксон предложили назвать Pegasoferae[4].
Эволюционная история
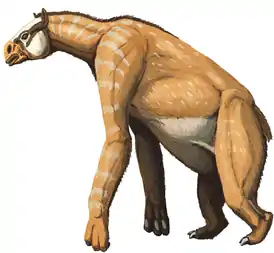
История развития непарнокопытных относительно хорошо известна благодаря многочисленным ископаемым. Этот отряд ранее был более разнообразным и распространённым, чем в наши дни. Наиболее древние находки из Европы и Северной Америки датируются началом эоцена. Представители разных линий непарнокопытных поначалу были весьма похожими друг на друга. Гиракотерий, считающийся предком лошадиных, был близок к гирахиусу, первому представителю линии носорогов. Оба вида были довольно малого размера по сравнению с более поздними формами и питались листвой, обитая в лесной местности. Период расцвета непарнокопытных продолжался от эоцена до миоцена, когда представители этого весьма разветвлённого отряда населяли весь земной шар, за исключением Австралии, Антарктиды и Южной Америки. Южноамериканский континент был заселён непарнокопытными после появления Панамского перешейка в плиоцене.
Период упадка непарнокопытных начался в среднем миоцене. Как правило, его связывают с распространением парнокопытных, которые занимали схожие экологические ниши и обладали преимуществом более эффективной пищеварительной системы. Несмотря на это, ряд родов непарнокопытных смог выстоять конкуренцию и продолжать существовать параллельно с парнокопытными. Вымирание мегафауны в конце плейстоцена затронуло и отряд непарнокопытных. В этот период исчезли лошадиные Нового Света, а также шерстистый носорог. Стало ли причиной изменение климата в конце ледникового периода, охота со стороны человека или оба фактора вместе взятых, является предметом научных споров.
К основным эволюционным линиям непарнокопытных относятся следующие группы (подотряды):
- Бронтотериеобразные (Brontotheria) были наиболее древними из крупных млекопитающих. Их наиболее известным представителем является бронтотерий. У них имелся костяной рог на носу. Плоские боковые зубы были приспособлены к пережёвыванию мягкой растительной пищи. Эти животные, распространённые в Северной Америки и Азии, полностью вымерли в начале олигоцена.
- Лошадеобразные (Hippomorpha) также появились в эоцене. Палеотериевые, известные прежде всего по находкам из Европы и включавшие гиракотериев, вымерли в олигоцене. Собственно лошадиные (Equidae), однако, широко распространились по всему миру. В развитии этой группы по ископаемым находкам хорошо прослеживается уменьшение количества пальцев, удлинение конечностей и прогрессирующие приспособление зубов к жёсткой травянистой пище.
- Анцилоподы (Ancylopoda) были подотрядом, у представителей которого вместо копыт развились когти и резко удлинились передние конечности. Наиболее значимым семейством анцилопод стали халикотериевые (Chalicotheriidae), в составе которых были халикотерии и моропусы. Первые вымерли лишь в плейстоцене.
- Носорогообразные (Rhinoceratoidea) были весьма многочисленными и разнообразными в период между эоценом и олигоценом. Встречались как пожиратели листвы величиной в собаку, животные с водным образом жизни наподобие современных бегемотов, а также огромные длинношеие виды. Рогом на носу обладали лишь немногие из них. Гирахиус является наиболее древним известным представителем этой группы. Водный образ жизни вело семейство аминодонтовых (Amynodontidae). Другое семейство, гиракодонтовые (Hyracodontidae) обладали длинными конечностями и шеями. Наиболее выраженным это было у парацератерий (ранее также называемых индрикотериями), являвшихся самыми крупными из известных наземных млекопитающих. Собственно носороги (Rhinocerotidae) появились в позднем эоцене или раннем олигоцене, до наших дней выжило пять видов.
- Тапирообразные (Tapiroidea) достигли наибольшего распространения в эоцене, когда различные роды обитали в Евразии и Северной Америке. У них более чем у других сохранилась древняя форма телосложения, однако примечательно развитие короткого хобота. К вымершим семействам относятся среди прочего Helaletidae и Lophiodontidae.

Отношения между этими крупными группами до сих пор остаются спорными. Более-менее бесспорным является то, что бронтотериевые были сестринской группой остальных непарнокопытных, а также тот факт, что носороги и тапировые более близки друг к другу, чем к лошадиным. Возможная кладограмма выглядит следующим образом[5]:
Непарнокопытные (Perissodactyla) ├─ † Бронтотериевые (Brontotheria) └─ Lophodontomorpha ├─ † Анцилоподы (Ancylopoda) └─ Euperissodactyla ├─ Лошадинообразные (Hippomorpha) │ ├─ † Палеотериевые (Palaeotheriidae) │ └─ Лошадиные (Equidae) └─ Ceratomorpha ├─ Носорогообразные (Rhinoceratoidea) │ ├─ † Аминодонтовые (Amynodontidae) │ ├─ † Гиракодонтовые (Hyracodontidae) │ └─ Носороги (Rhinocerotidae) └─ Тапирообразные (Tapiroidea) ├─ † Гелалетовые (Helaletidae) ├─ † Лофиодонтовые (Lophiodontidae) └─ Тапировые (Tapiridae)
Согласно генетическому исследованию, опубликованному в 2015 г., к непарнокопытным также относились литоптерны и примитивные дидолодонтиды[6][7].
Непарнокопытные и человек

Домашняя лошадь и домашний осёл сыграли в человеческой истории большую роль в качестве животных для верховой езды, перевозки грузов и сельскохозяйственных работ. Одомашнивание обоих видов началось за несколько тысяч лет до нашей эры. Ввиду моторизации сельского хозяйства и распространения автотранспорта значение этих животных в развитых странах снизилось. Сейчас их содержанием занимаются главным образом в спортивных целях или как хобби. Однако в менее развитых регионах планеты использование непарнокопытных всё ещё широко распространено. В меньшей мере их содержат и разводят ради мяса и молока.
В отличие от домашних лошадей и ослов, популяции всех остальных видов непарнокопытных вследствие охоты и разрушения жизненного пространство резко сократились. Исчезла квагга, близка к исчезновению также лошадь Пржевальского. Четыре вида — африканского осла, суматранского носорога, яванского носорога и чёрного носорога — МСОП определяет как подверженных критической опасности исчезновения (critically endangered). Пять других видов — зебра Греви, горная зебра, горный тапир, центральноамериканский тапир и индийский носорог — считаются подверженными опасности исчезновения (endangered).
Примечания
- Lisa Noelle Cooper et al. Anthracobunids from the Middle Eocene of India and Pakistan Are Stem Perissodactyls (англ.) // PLOS ONE. — 2014. — Vol. 9, no. 10. — doi:10.1371/journal.pone.0109232.
- Предок слонов оказался древним носорогом. PaleoNews (14 октября 2014). Дата обращения: 14 октября 2014. Архивировано 17 октября 2014 года.
- Согласно Westheide/Rieger (2004), стр. 503.
- Hidenori Nishihara, Masami Hasegawa и Norihiro Okada: Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions, в Proceedings of the National Academy of Sciences 103, 2006; p. 9929—9934 (Весь текст доступен в качестве PDF Архивная копия от 23 марта 2008 на Wayback Machine)
- J. J. Hooker и D. Dashzeveg: The origin of chalicotheres (Perissodactyla, Mammalia). в: Palaeontology: Vol. 47 Part 6, 2004, S. 1363—1386 ([www.blackwell-synergy.com/doi/pdf/10.1111/j.0031-0239.2004.00421.x#search=%22lophodontomorpha%22 PDF-Version] (недоступная ссылка))
- Welker F. et al. Ancient proteins resolve the evolutionary history of Darwin’s South American ungulates // Nature Letter. — 2015. — doi:10.1038/nature14249.
- Палеонтологи разобрались в родственных связях южноамериканских копытных. Дата обращения: 29 июля 2021. Архивировано 29 июля 2021 года.