Бета-шпилька
Бета-шпилька (иногда также называемая бета-лентой или бета-бета-единицей) представляет собой простой структурный мотив белка, включающий две бета-нити, которые похожи на шпильку. Мотив состоит из двух смежных по первичной структуре нитей, ориентированных в антипараллельном направлении (N-конец одного листа примыкает к C-концу следующего), связанных короткой петлей из двух-пяти аминокислот. Бета-шпильки могут встречаться изолированно или как часть ряда нитей, связанных водородными связями, которые вместе составляют бета-лист.

Такие исследователи, как Франсиско Бланко и др. использовали ЯМР белков, чтобы показать, что бета-шпильки могут быть образованы из изолированных коротких пептидов в водном растворе, предполагая, что шпильки могут образовывать сайты зарождения для сворачивания белка[1].
Классификация
Бета-шпильки изначально классифицировались исключительно по количеству аминокислотных остатков в их последовательностях петель, так что они были названы одноостаточными, двухостаточными и т. д.[2] Эта система, однако, несколько неоднозначна, поскольку не принимает во внимание, связаны ли остатки, которые сигнализируют о конце шпильки, одинарной или двойной водородной связью друг с другом. С тех пор Милнер-Уайт и Поэт предложили усовершенствованный способ классификации[3].
Бета-шпильки разделены на четыре отдельных класса. Каждый класс начинается с наименьшего возможного числа остатков в петле и постепенно увеличивает размер петли, удаляя водородные связи в бета-листе. Первичная шпилька класса 1 представляет собой петлю с одним остатком, в которой связанные остатки имеют две водородные связи. Затем удаляется одна водородная связь, образуя петлю из трех остатков, которая является вторичной шпилькой класса 1. Односвязанные остатки подсчитываются в последовательности петли, но также сигнализируют об окончании петли, таким образом определяя эту шпильку как петлю с тремя остатками. Затем эта одинарная водородная связь удаляется, чтобы создать третичную шпильку; петля из пяти остатков с дважды связанными остатками. Этот шаблон продолжается бесконечно и определяет все бета-шпильки внутри класса. Класс 2 следует той же схеме, начиная с петли из двух остатков с концевыми остатками, которые разделяют две водородные связи. Класс 3 начинается с трех остатков, а класс 4 — с четырёх остатков. Класса 5 не существует, поскольку данная основная шпилька уже определена в классе 1. Эта классификационная схема не только учитывает различные степени водородных связей, но также говорит о биологическом поведении шпильки. Замена одной аминокислоты может разрушить конкретную водородную связь, но не развернет шпильку или не изменит её класс. С другой стороны, вставки и делеции аминокислот должны будут развернуть и преобразовать всю бета-цепь, чтобы избежать бета-выпуклости во вторичной структуре. Это изменит класс шпильки в процессе. Поскольку замены являются наиболее распространенными аминокислотными мутациями, белок потенциально может подвергаться преобразованию, не влияя на функциональность бета-шпильки[3].
Динамика складывания и переплета
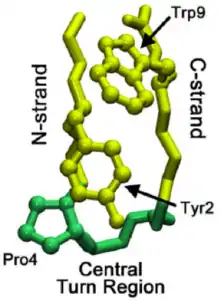
Понимание механизма сворачивания микродоменов может помочь пролить свет на паттерны сворачивания целых белков. Исследования бета-шпильки под названием чигнолин (см. «Чигнолин в Proteopedia») выявили ступенчатый процесс складывания, который приводит в движение бета-шпильку. Эта шпилька имеет особенности последовательности, сходные с более чем 13000 известных шпилек, и, таким образом, может служить более общей моделью образования бета-шпильки. Формирование области нативного поворота сигнализирует о начале каскада складывания, где нативный поворот — это тот, который присутствует в окончательной складчатой структуре.
При сворачивании всех белков поворот может происходить не в области нативного поворота, а в С-цепи бета-шпильки. Затем этот поворот распространяется через C-цепь (бета-цепь, ведущая к C-концу), пока не достигнет области нативного поворота. Иногда взаимодействия остатков, ведущих к области нативного поворота, слишком сильны, вызывая его разворачивание. Однако, как только нативный поворот действительно формируется, взаимодействия между пролинами и остатками триптофана (видно на изображении справа) в области помогают стабилизировать поворот, предотвращая «откат» или разворачивание.
Исследователи полагают, что повороты не возникают в N-цепи из-за повышенной жесткости (часто вызываемой пролином, ведущим к области нативного витка) и меньшего количества вариантов конформации. Формирование начального витка происходит примерно за 1 мкс. Как только начальный поворот был установлен, были предложены два механизма относительно того, как складывается остальная часть бета-шпильки: гидрофобный коллапс с перестройками уровня боковой цепи или более общепринятый механизм, подобный застежке-молнии[4].
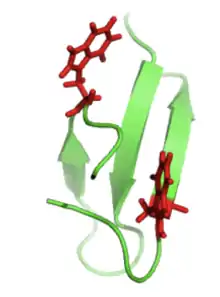
Мотив петли β-шпильки можно найти во многих макромолекулярных белках. Однако небольшие и простые β-шпильки могут существовать и сами по себе. Чтобы ясно это увидеть, в качестве примера слева показан белок домена Pin1.
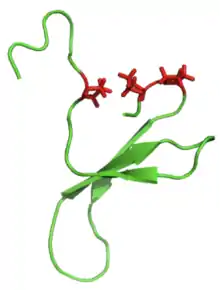
Белки, богатые β-слоями, также называемые доменами WW, функционируют путем присоединения к богатым пролином и / или фосфорилированным пептидам, опосредуя межбелковые взаимодействия. «WW» относится к двум остаткам триптофана (W), которые консервативны в последовательности и способствуют сворачиванию β-листов с образованием небольшого гидрофобного ядра[5]. Остатки триптофана можно увидеть ниже (справа) красным цветом.
Этот фермент связывает свой лиганд за счет сил Ван-дер-Ваальса консервативных триптофанов и богатых пролином участков лиганда. Другие аминокислоты могут затем связываться с гидрофобным ядром структуры β-шпильки для обеспечения надежного связывания[6].
Также часто можно обнаружить остатки пролина внутри петлевой части β-шпильки, поскольку эта аминокислота является жесткой и способствует образованию «витка». Эти остатки пролина можно увидеть как красные боковые цепи на изображении WW домена Pin1 ниже (слева).
Искусственно созданная бета-шпилька
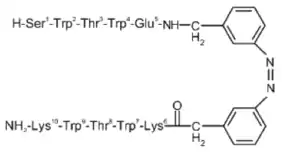
Конструкция пептидов, которые принимают структуру β-шпильки (без опоры на связывание металлов, необычные аминокислоты или дисульфидные поперечные связи), достигла значительного прогресса и позволила понять динамику белков. В отличие от α-спиралей, β-шпильки не стабилизируются регулярным рисунком водородных связей. В результате ранние попытки требовали по крайней мере 20-30 аминокислотных остатков для достижения стабильных третичных складок β-шпилек. Однако этот нижний предел был снижен до 12 аминокислот из-за повышения стабильности за счет включения пар перекрестных цепей триптофан-триптофан. Было показано, что две пары триптофана, не связывающиеся с водородом, сцепляются в мотив, похожий на застежку-молнию, стабилизируя структуру β-шпильки, но при этом позволяя ей оставаться водорастворимой . Структура ЯМР β-пептида триптофановой застежки-молнии (trpzip) показывает стабилизирующий эффект благоприятных взаимодействий между соседними индольными кольцами[7].
Синтез пептидов β-шпильки trpzip включает фотопереключатели, которые облегчают точный контроль за складыванием. Некоторые аминокислоты, в свою очередь, заменяются азобензолом, который можно заставить переключиться из транс-конформации в цис-конформацию под действием света с длиной волны 360 нм. Когда азобензольный фрагмент находится в цис-конформации, аминокислотные остатки выравниваются правильно, принимая образование β-шпильки. Однако транс-конформация не имеет правильной геометрии поворота для β-шпильки[8]. Это явление можно использовать для исследования конформационной динамики пептидов с помощью фемтосекундной абсорбционной спектроскопии[8].
Примечания
- Blanco, F. J. (1994). “A short linear peptide that folds into a native stable beta-hairpin in aqueous solution”. Nat Struct Biol. 1 (9): 584—590. DOI:10.1038/nsb0994-584. PMID 7634098.
- Sibanda, B.L.; Blundell, T.L.; Thorton, J.M. (1985). «Conformations of Beta-Hairpins in Protein Structures». Nature(London) 316 170—174.
- Milner-White, J.; Poet, R. (1986). «Four Classes of Beta-Hairpins in Proteins». Biochemical Journal 240 289—292.
- Enemark, Søren (11 September 2012). “β-hairpin forms by rolling up from C-terminal: Topological guidance of early folding dynamics”. Scientific Reports. 2: 649. Bibcode:2012NatSR...2E.649E. DOI:10.1038/srep00649. PMID 22970341.
- Jager, Marcus (2008). “Understanding the mechanism of β-sheet folding from a chemical and biological perspective”. Biopolymers. 90 (6): 751—758. DOI:10.1002/bip.21101. PMID 18844292.
- Kay, B.K.; Williamson, M.P.; Sudol, M. The Importance of Being Proline: the interaction of proline-rich motifs in signaling proteins with their cognate domains. The FASEB Journal. 2000, 14, 231—241.
- Cochran, Andrea G. (2001-05-08). “Tryptophan zippers: Stable, monomeric β-hairpins”. Proceedings of the National Academy of Sciences [англ.]. 98 (10): 5578—5583. Bibcode:2001PNAS...98.5578C. DOI:10.1073/pnas.091100898. ISSN 0027-8424. PMID 11331745.
- Dong, Shou-Liang (2006-01-23). “A Photocontrolled β-Hairpin Peptide”. Chemistry – A European Journal [англ.]. 12 (4): 1114—1120. DOI:10.1002/chem.200500986. ISSN 1521-3765. PMID 16294349.
Вторичная структура белка | ||
|---|---|---|
| Спирали |  | |
| Расширения |
| |
| Сверхвторичная структура | ||