Xist
Xist (англ. X-inactive specific transcript) — ген, кодирующий РНК и локализованный на Х-хромосоме плацентарных млекопитающих, является ключевым эффектором в инактивации Х-хромосомы[1]. Он входит в состав комплекса Xic (англ. X-chromosome inactivation centre)[2], наряду с двумя другими РНК-кодирующими генами (Jpx и Ftx) и двумя белоккодирующими генами (Tsx и Cnbp2)[3]. Продукт гена Xist — Xist-РНК — представляет собой крупный транскрипт (17 кб у человека)[4], который экспрессируется на неактивной хромосоме и не экспрессируется на активной. Процессинг этого транскрипта напоминает процессинг мРНК и тоже включает этапы сплайсинга (в т. ч. альтернативного[1]) и полиаденилирования, однако он остаётся в ядре и не транслируется. Было высказано предположение, что ген Xist по крайней мере частично образовался как часть белоккодирующего гена, который впоследствии стал псевдогеном[5]. Инактивированная Х-хромосома покрыта Xist-РНК, который необходим для процесса инактивации[6]. Х-хромосома, лишённая гена Xist, не будет инактивирована, однако дупликация этого гена на другой хромосоме вызывает инактивацию и первой хромосомы тоже[7].
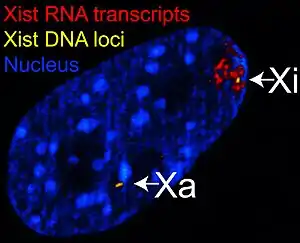
Инактивация Х-хромосомы, которая происходит на ранних стадиях развития самок плацентарных млекопитающих, обеспечивает сайленсинг на уровне транскрипции одной из двух Х-хромосом, что обеспечивает дозовое равенство (т. е. по количеству активных вариантов одного гена) самок и самцов (см. подробнее «Дозовая компенсация»). Функциональная роль Xist-РНК была наглядно продемонстрирована на эмбриональных стволовых клетках самок мышей. В ходе этих экспериментов введение в клетки антисмысловой РНК длиной 19 нуклеотидов, мишенью которой была Xist-РНК, предотвращало образование Xic и цис-сайленсинг Х-связанных генов. Впрочем, в дальнейшем было показано, что инактивация Х-хромосомы у мышей всё-таки происходит даже в отсутствие гена Xist посредством эпигенетической регуляции, но Xist-РНК необходима для стабилизации такого сайленсинга[8].
Было показано, что ген Xist взаимодействует с геном BRCA1, связанным с раком молочной железы[9][10].
Организация гена
У человека ген Xist локализован на длинном (q) плече Х-хромосомы. Он включает в себя большое количество повторов[4] и состоит из А-региона, содержащего 8 повторов, разделённых U-спейсерами. А-регион содержит две крупные шпильки, в каждую из которых входит по 4 повтора[11]. Ортолог гена Xist у человека был идентифицирован у мыши, его длина составляет 15 кб, однако он не содержит консервативных повторов[12].
Организация транскрипта
Xist-РНК состоит из двух регионов: А и С. Консервативный А-регион содержит до 9 повторяющихся элементов[11]. Недавно было показано, что у мыши и человека А-регион Xist-РНК состоит из двух длинных шпилек, в состав каждой из которых входит по 4 повтора[4][11]. Хотя конкретная функция А-региона неизвестна, было показано, что он необходим для эффективного связывания с белком Suz12[11].
Связывание Xist-РНК с инактивированной Х-хромосомой осуществляется через хроматин-связывающий участок, расположенный на самом транскрипте. Впервые такой участок на Xist-РНК был описан в фибробластах самки мыши. Он оказался локализованным в области С-повтора. Вышеописанные эксперименты по введению в клетки 19-нуклеотидной антисмысловой РНК, комплементарной Xist, нарушали связывание этого транскрипта с гистоном H2A[13].
Регуляция Xist
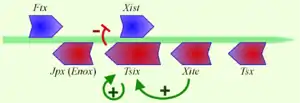
Как упоминалось выше, ген Xist-РНК входит в состав инактивационного центра Xic (англ. X Inactivation Center)[14]. Xic локализуется на q-плече Х-хромосомы (Xq13). Ключевую роль в инактивации Х-хромосомы играет промотор Xist, входящий в состав Xiс[15]. Кроме гена Xist, в Xic входит также ген Tsix, антисмысловой по отношению к Xist. Антисмысловые транскрипты гена Xist действуют как цис-регуляторы транскрипции Xist, понижая экспрессию этого гена. Механизм такой цис-регуляции экспрессии Xist с помощью Tsix пока плохо понятен, хотя существует несколько объясняющих гипотез. Согласно одной из них, Tsix участвует в модификации хроматина в локусе Xist[16] (подробнее об этом см. ниже).
Предполагается, что антисмысловой транскрипт Tsix активирует ДНК-метилтрансферазы, которые метилируют промотор Xist, что приводит к подавлению этого промотора и, следовательно, экспрессии гена Xist[17]. Показана роль ацетилирования гистонов в регуляции Xist[18].
Возможно, двуцепочечные РНК и интерферирующие РНК также играют роль в регуляции промотора Xist. По-видимому, фермент Dicer, специализирующийся на разрезании двуцепочечных РНК, разрезает дуплекс Xist и Tsix в начале процесса инактивации Х-хромосомы, в результате чего образуются короткие РНК длиной около 30 нуклеотидов (xiРНК). Предполагается, что эти xiРНК репрессируют Xist на той Х-хромосоме, которая останется активной. Это предположение подтверждается экспериментом, в ходе которого в недифференцированных клетках количество эндогенного Dicer было снижено на 5 %, что привело к увеличению Xist в этих клетках[19].
По-видимому, транскрипционные факторы плюрипотентных клеток Nanog, Oct4 и Sox2 играют роль в сайленсинге гена Xist. В отсутствие Tsix в плюрипотентных клетках Xist, тем не менее, тоже репрессирован. Возможным объяснением этому может быть то, что вышеперечисленные факторы вызывают сплайсинг по интрону 1 в сайте связывания этих факторов в гене Xist, в результате чего сайт разрушается и экспрессия Xist подавляется[16]. В плюрипотентных клетках, лишённых Nanog или Oct4, уровень экспрессии Xist повышался[20].
Была показана роль репрессорного комплекса polycomb 2 (англ. Polycomb Repressor Complex 2 (PRC2)) в сайленсинге Xist независимо от Tsix, хотя конкретные механизмы этого неизвестны. PRC2 — это класс белков группы polycomb, которые вызывают триметилирование гистона Н3 по остатку лизина 27 (К27), что подавляет транскрипцию путём перестройки хроматина. Вышеупомянутый белок Suz12 входит в группу PRC2 и имеет домен цинкового пальца, который, по-видимому, связывается с молекулой РНК[21].
Значение и механизмы инактивации
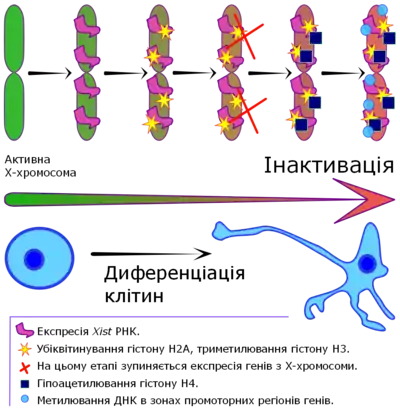
Процесс инактивации Х-хромосомы начинается с распространением Xist-РНК от Xic по всей хромосоме, где Xist-РНК, по-видимому, индуцирует образование и расширение области гетерохроматина. Гетерохроматин инактивированной Х-хромосомы характеризуется не только связью с Xist-РНК, но также наличием особого варианта гистона Н2А (macroH2A), гипоацетилированием гистонов Н3 и Н4, убиквитинированием гистона H2А и метилированием специфических позиций на гистоне Н3, а также метилирования ДНК. Именно комбинация подобных модификаций обусловливает большую часть инактивированной Х-хромосомы чрезвычайно невосприимчивой к транскрипции. Кроме того, способность этих модификаций к самораспространению обеспечивает поддержание инактивированного состояния Х-хромосомы в течение многих клеточных делений. Впрочем, стоит отметить, около 10 % генов на Х-хромосоме остаются транскрипционно активными[22].
Экспрессия Xist и инактивация Х-хромосомы изменяются в процессе эмбрионального развития. У мыши в раннем эмбриональном развитии существует две волны инактивации Х-хромосомы. Первая волна происходит до образования бластоцисты, и является неслучайной — в клетках зародыша инактивируется Х-хромосома, полученная от отца, то есть инактивация Х-хромосомы на этом этапе носит импринтированный характер. На стадии бластоцисты, в клетках внутренней клеточной массы, из которой формируются в дальнейшем все органы и ткани будущего организма, этот импринтинг снимается, экспрессия Xist исчезает, инактивация отцовской Х-хромосомы отменяется, и обе Х-хромосомы становятся транскрипционно активными. Недавно полученные данные показали, что в процессе реактивации Х-хромосомы участвуют антисмысловые транскрипты[23]. В формирующихся далее клетках эпибласта начинается процесс дифференцировки, и наступает вторая волна инактивации, в которой выбор инактивируемой Х-хромосомы носит случайный характер. На одной из Х-хромосом начинает экспрессироваться Xist, и эта Х-хромосома вступает в процесс инактивации. Вследствие случайного выбора инактивируемой Х-хромосомы каждый женский организм представляет собой мозаику из клональных групп клеток, у которых либо инактивирована Х-хромосома, доставшаяся от отца, либо Х-хромосома, оставшаяся от матери. Инактивация Х-хромосомы поддерживается в течение тысячи клеточных делений[22], и только в развивающихся гоноцитах экспрессия Xist снижается и Х-хромосома вновь реактивируется[24]. В экстраэмбриональных тканях у мыши импринтированная инактивация отцовской Х-хромосомы, установившаяся во время первой волны инактивации, сохраняется в течение всего эмбрионального развития.
Инактивация Х-хромосомы играет ключевую роль в механизме дозовой компенсации, благодаря которому обеспечивается равенство доз генных продуктов Х-хромосомы у обоих полов[22][25]. У различных видов дозовая компенсация обеспечивается различными путями, но во всех таких путях имеет место регуляции экспрессии Х-хромосомы у одного из двух обоих полов[25]. Если одна из двух Х-хромосом не будет инактивирована или будет частично экспрессироваться, то получающаяся в результате избыточная экспрессия Х-хромосомы может быть летальной[22].
Клиническое значение
У человека мутации в промоторе Xist обусловливают семейную неслучайную инактивацию Х-хромосомы[1].
В 2013 году было показано, что внедрение гена Xist в одну из 21-х хромосом стволовой клетки с трисомией по 21-ой хромосоме (причина синдрома Дауна) позволяет инактивировать эту хромосому. Таким образом, ген Xist может лечь в основу нового подхода к лечению синдрома Дауна[26].
Примечания
- Entrez Gene: XIST X (inactive)-specific transcript.
- Chow J. C., Yen Z., Ziesche S. M., Brown C. J. Silencing of the mammalian X chromosome. (англ.) // Annual review of genomics and human genetics. — 2005. — Vol. 6. — P. 69—92. — doi:10.1146/annurev.genom.6.080604.162350. — PMID 16124854.
- Chureau C., Prissette M., Bourdet A., Barbe V., Cattolico L., Jones L., Eggen A., Avner P., Duret L. Comparative sequence analysis of the X-inactivation center region in mouse, human, and bovine. (англ.) // Genome research. — 2002. — Vol. 12, no. 6. — P. 894—908. — doi:10.1101/gr.152902. — PMID 12045143.
- Brown C. J., Hendrich B. D., Rupert J. L., Lafrenière R. G., Xing Y., Lawrence J., Willard H. F. The human XIST gene: analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. (англ.) // Cell. — 1992. — Vol. 71, no. 3. — P. 527—542. — PMID 1423611.
- Duret L., Chureau C., Samain S., Weissenbach J., Avner P. The Xist RNA gene evolved in eutherians by pseudogenization of a protein-coding gene. (англ.) // Science (New York, N.Y.). — 2006. — Vol. 312, no. 5780. — P. 1653—1655. — doi:10.1126/science.1126316. — PMID 16778056.
- Ng K., Pullirsch D., Leeb M., Wutz A. Xist and the order of silencing. (англ.) // EMBO reports. — 2007. — Vol. 8, no. 1. — P. 34—39. — doi:10.1038/sj.embor.7400871. — PMID 17203100.
- Penny G. D., Kay G. F., Sheardown S. A., Rastan S., Brockdorff N. Requirement for Xist in X chromosome inactivation. (англ.) // Nature. — 1996. — Vol. 379, no. 6561. — P. 131—137. — doi:10.1038/379131a0. — PMID 8538762.
- Kalantry S., Purushothaman S., Bowen R. B., Starmer J., Magnuson T. Evidence of Xist RNA-independent initiation of mouse imprinted X-chromosome inactivation. (англ.) // Nature. — 2009. — Vol. 460, no. 7255. — P. 647—651. — doi:10.1038/nature08161. — PMID 19571810.
- Ganesan S., Silver D. P., Drapkin R., Greenberg R., Feunteun J., Livingston D. M. Association of BRCA1 with the inactive X chromosome and XIST RNA. (англ.) // Philosophical transactions of the Royal Society of London. Series B, Biological sciences. — 2004. — Vol. 359, no. 1441. — P. 123—128. — doi:10.1098/rstb.2003.1371. — PMID 15065664.
- Ganesan S., Silver D. P., Greenberg R. A., Avni D., Drapkin R., Miron A., Mok S. C., Randrianarison V., Brodie S., Salstrom J., Rasmussen T. P., Klimke A., Marrese C., Marahrens Y., Deng C. X., Feunteun J., Livingston D. M. BRCA1 supports XIST RNA concentration on the inactive X chromosome. (англ.) // Cell. — 2002. — Vol. 111, no. 3. — P. 393—405. — PMID 12419249.
- Maenner S., Blaud M., Fouillen L., Savoye A., Marchand V., Dubois A., Sanglier-Cianférani S., Van Dorsselaer A., Clerc P., Avner P., Visvikis A., Branlant C. 2-D structure of the A region of Xist RNA and its implication for PRC2 association. (англ.) // Public Library of Science Biology. — 2010. — Vol. 8, no. 1. — P. e1000276. — doi:10.1371/journal.pbio.1000276. — PMID 20052282.
- Brockdorff N., Ashworth A., Kay G. F., McCabe V. M., Norris D. P., Cooper P. J., Swift S., Rastan S. The product of the mouse Xist gene is a 15 kb inactive X-specific transcript containing no conserved ORF and located in the nucleus. (англ.) // Cell. — 1992. — Vol. 71, no. 3. — P. 515—526. — PMID 1423610.
- Beletskii A., Hong Y. K., Pehrson J., Egholm M., Strauss W. M. PNA interference mapping demonstrates functional domains in the noncoding RNA Xist. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2001. — Vol. 98, no. 16. — P. 9215—9220. — doi:10.1073/pnas.161173098. — PMID 11481485.
- Herzing L. B., Romer J. T., Horn J. M., Ashworth A. Xist has properties of the X-chromosome inactivation centre. (англ.) // Nature. — 1997. — Vol. 386, no. 6622. — P. 272—275. — doi:10.1038/386272a0. — PMID 9069284.
- Lee J. T., Davidow L. S., Warshawsky D. Tsix, a gene antisense to Xist at the X-inactivation centre. (англ.) // Nature genetics. — 1999. — Vol. 21, no. 4. — P. 400—404. — doi:10.1038/7734. — PMID 10192391.
- Senner C. E., Brockdorff N. Xist gene regulation at the onset of X inactivation. (англ.) // Current opinion in genetics & development. — 2009. — Vol. 19, no. 2. — P. 122—126. — doi:10.1016/j.gde.2009.03.003. — PMID 19345091.
- Nesterova T. B., Popova B. C., Cobb B. S., Norton S., Senner C. E., Tang Y. A., Spruce T., Rodriguez T. A., Sado T., Merkenschlager M., Brockdorff N. Dicer regulates Xist promoter methylation in ES cells indirectly through transcriptional control of Dnmt3a. (англ.) // Epigenetics & chromatin. — 2008. — Vol. 1, no. 1. — P. 2. — doi:10.1186/1756-8935-1-2. — PMID 19014663.
- Csankovszki G., Nagy A., Jaenisch R. Synergism of Xist RNA, DNA methylation, and histone hypoacetylation in maintaining X chromosome inactivation. (англ.) // The Journal of cell biology. — 2001. — Vol. 153, no. 4. — P. 773—784. — PMID 11352938.
- Ogawa Y., Sun B. K., Lee J. T. Intersection of the RNA interference and X-inactivation pathways. (англ.) // Science (New York, N.Y.). — 2008. — Vol. 320, no. 5881. — P. 1336—1341. — doi:10.1126/science.1157676. — PMID 18535243.
- Navarro P., Chambers I., Karwacki-Neisius V., Chureau C., Morey C., Rougeulle C., Avner P. Molecular coupling of Xist regulation and pluripotency. (англ.) // Science (New York, N.Y.). — 2008. — Vol. 321, no. 5896. — P. 1693—1695. — doi:10.1126/science.1160952. — PMID 18802003.
- de Napoles M., Mermoud J. E., Wakao R., Tang Y. A., Endoh M., Appanah R., Nesterova T. B., Silva J., Otte A. P., Vidal M., Koseki H., Brockdorff N. Polycomb group proteins Ring1A/B link ubiquitylation of histone H2A to heritable gene silencing and X inactivation. (англ.) // Developmental cell. — 2004. — Vol. 7, no. 5. — P. 663—676. — doi:10.1016/j.devcel.2004.10.005. — PMID 15525528.
- Альбертс и др., 2013, с. 729.
- Mak W., Nesterova T. B., de Napoles M., Appanah R., Yamanaka S., Otte A. P., Brockdorff N. Reactivation of the paternal X chromosome in early mouse embryos. (англ.) // Science (New York, N.Y.). — 2004. — Vol. 303, no. 5658. — P. 666—669. — doi:10.1126/science.1092674. — PMID 14752160.
- Nesterova T. B., Mermoud J. E., Hilton K., Pehrson J., Surani M. A., McLaren A., Brockdorff N. Xist expression and macroH2A1.2 localisation in mouse primordial and pluripotent embryonic germ cells. (англ.) // Differentiation; research in biological diversity. — 2002. — Vol. 69, no. 4-5. — P. 216—225. — doi:10.1046/j.1432-0436.2002.690415.x. — PMID 11841480.
- Nguyen D. K., Disteche C. M. Dosage compensation of the active X chromosome in mammals. (англ.) // Nature genetics. — 2006. — Vol. 38, no. 1. — P. 47—53. — doi:10.1038/ng1705. — PMID 16341221.
- Jiang J., Jing Y., Cost G. J., Chiang J. C., Kolpa H. J., Cotton A. M., Carone D. M., Carone B. R., Shivak D. A., Guschin D. Y., Pearl J. R., Rebar E. J., Byron M., Gregory P. D., Brown C. J., Urnov F. D., Hall L. L., Lawrence J. B. Translating dosage compensation to trisomy 21. (англ.) // Nature. — 2013. — Vol. 500, no. 7462. — P. 296—300. — doi:10.1038/nature12394. — PMID 23863942.
Литература
- Б. Альбертс, А. Джонсон, Д. Льюис и др. Молекулярная биология клетки / Перевод с английского - А.Н. Дьяконовой, А.В. Дюбы и А.А. Светлова. Под ред. - Е.С. Шилова, Б.П. Копнина, М.А. Лагарьковой, Д.В. Купраша.. — М.—Ижевск: НИЦ «Регулярная и хаотическая динамика», 2013. — Т. 1. — С. 1213. — 1052 с. — ISBN 978-5-4344-0112-8.
- Шевченко А. И., Захарова И. С., Закиян С. М. Эволюционный путь процесса инактивации Х-хромосомы у млекопитающих // Acta Naturae. — 2013. — Т. 5, № 2. — С. 40-54.
Ссылки
- Кочанова, Наталья. Загадочное путешествие некодирующей РНК Xist по X-хромосоме. // Сайт Biomolecula.ru (21 октября 2013). Дата обращения: 21 марта 2018.