Фотоингибирование
Фотоингибирование — индуцируемое светом снижение фотосинтетической активности растений, водорослей или цианобактерии. Фотосистема II (ФСII) более чувствительна к свету, чем другие фотосинтетические машины, поэтому большинство исследователей определяют термин как светоиндуцируемое повреждения ФСII. В живых организмах, поврежденные избыточным освещением фотосистемы II постоянно репарируются за счёт деградации и синтеза белка D1 фотосинтетического реакционного центра ФСII. Понятие фотоингибирование также используется в более широком смысле, в значении «динамическое фотоингибирование», чтобы описать все реакции, которые приводят к снижению эффективности фотосинтеза, когда растения подвергаются воздействию света.
.svg.png.webp)
История
Первым учёным, который на практике измерившим уровень фотоингибирования был Бессель Кок, который опубликовал свои результаты в 1956 году[1]. Уже на ранних этапах исследования феномена, стало очевидно, что растения имеют некий механизм репарации, который постоянно устраняет повреждения. В 1966 году, Джонс и Кок измерили спектр действия фотоингибирования и обнаружили, что наибольший эффект оказывал ультрафиолетовый свет[2]. Было также установлено, что в видимой части спектра пик фотоингибирования находился в красном регионе. В 1980-х годах фотоингибирование стало популярной темой среди исследователей фотосинтеза, а концепция повреждения и репарации была переоткрыта. Началу исследований положила статья Кайла Охада и Арнтцена 1984 года, в которой они показали, что фотоингибирование сопровождается селективной потерей белка 32-кДа, позже идентифицированного как белок D1[3]. Фоточувствительность ФСII с химически инактивированным водоокисляющим комплексом изучалась в 1980-х и начале 1990-х годов[4][5]. В статье Имре Васса и коллег от 1992 года была описана акцепторная сторона механизма фотоингибирования[6]. Измерение скорости образования синглетного кислорода фотосистемой II в условиях фотоингибирования представило дополнительные доказательства в пользу акцепторного механизма[7]. Концепция цикла репарация, в котором постоянно происходит репарация повреждений, эволюционировала и была рассмотрен Аро и соавт. в 1993 году[8]. С тех пор были обнаружены многие детали цикла репарации, включая тот факт, что FtsH протеаза играет важную роль в деградации белка D1[9]. В 1996 году в статье Таэстчжавы и Аро было показано, что константа скорости фотоингибирования прямо пропорциональна интенсивности света, что противоречило более раннему предположению о том, что фотоингибирование вызвано долей световой энергии, которая превышает максимальные возможности фотосинтеза[10]. В следующем году группа Ицхака Охады провела эксперимент по фотоингибированию фотосистемы II лазерным импульсом, и на основании полученных данных высказали предположение, что разделение зарядов может быть вредными, поскольку оно может привести к образованию синглетного кислорода[11]. В научной среде постоянно обсуждаются возможный молекулярный механизм (или механизмы) фотоингибирования. Новейшим кандидатом является марганцевый механизм, предложенной в 2005 группой Эсы Таэстчжавы[12]. Сходный механизм был предложен группой Норио Мурата в том же 2005 году[13].
Что ингибируется
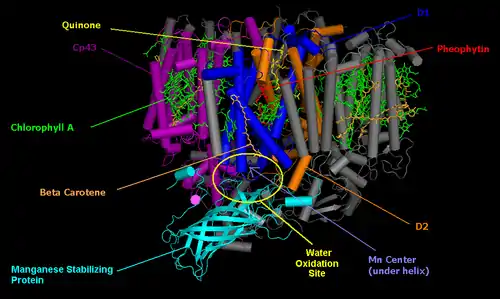
Фотоингибирование происходит у всех организмов, способных к оксигенному фотосинтезу, от сосудистых растений и до цианобактерий[14][15]. Во всех случаях синий свет вызывает более сильный эффект, чем другие длины волн видимого света, а из всех длин волн самое сильное деструктивное влияние оказывает ультрафиолетовый свет[14]. По сути своей фотоингибирование представляет серию фотохимических реакций, которые подавляют ту или иную активность ФСII, но нет консенсуса относительно того, что это за реакции. Первым как правило утрачивает свою активность водоокисляющий комплекс[12][13][16][17]. Однако, ингибирование ФСII в анаэробных условиях приводит в основном к ингибированию переноса электронов на акцепторной стороне ФСII (перенос электрона от специальной пары хлорофиллов к пластохинону)[6]. Ультрафиолетовый свет ингибирует водоокисляющий комплекс ещё до того, как остальная ФСII потеряет активность. Фотосистема I (ФСI) менее чувствительна к повреждению светом чем ФСII, но всё таки наблюдается её медленное ингибирование с течением времени[18]. Фотоингибирование ФСI происходит у холодочувствительных растений и зависит от потока электронов от ФСII к ФСI.
Как часто происходят повреждения?
Фотосистема II повреждается светом независимо от его интенсивности. Квантовый выход повреждающей реакции в типичных листьях высших растений или в препаратах тилакоидной мембраны находится в диапазоне от 10−8 до 10−7 и не зависит от интенсивности света[10][19]. Это означает, что повреждается примерно один комплекс фотосистемы II на каждые 10-100 миллионов захваченных фотонов. Из этих данных следует, что фотоингибирование происходит при любой интенсивности света и константа скорости фотоингибирования прямо пропорциональна его интенсивности. Некоторые результаты указывают на то, что тусклый свет вызывает даже более сильные повреждения, чем яркий свет[11].
Молекулярные механизмы
Механизм(ы) фотоингибирования находятся в стадии обсуждения, было предложено несколько разных механизмов[16]. Активные формы кислорода, особенно синглетного кислорода, играют роль в механизме ингибирования акцепторной стороны фотосистемы II, механизме синглетного кислорода и механизме ингибирования при низкой освещённости. Однако активные формы кислорода не играют непосредственную роль в марганцевом механизме и механизме ингибирования донорной стороны фотосистемы II. Фотоингибирование ФСII приводит к образованию синглетного кислорода[7] и других активных форм кислорода, которые ингибируют цикл репарации ФСII, подавляя синтез белка в хлоропласте[20].
Фотоингибирование акцепторной стороны
Яркий свет вызывает восстановление пула пластохинона, что приводит к протонированию и двойному восстановлению (и двойному протонированию) электронного акцептора QА Фотосистемы II. Протонированная и полностью восстановленная форма QА не может участвовать в транспорте электронов. Кроме того, реакция разделения зарядов в заингибированной фотосистеме II с большой вероятностью приводит к переходу первичного донора электронов (П680) в триплетное состояние. Триплет П680 может вступать в реакцию с кислородом и образовывать высоко реакционноспособный синглетный кислород[6].
Фотоингибирование донорной стороны
Если водоокисляющий комплекс инактивировать химически, то остальные элементы переноса электронов становится очень чувствительны к свету[4][19]. Было высказано предположение, что даже в здоровом листе водоокисляющий комплекс не всегда функционируют во всех комплексах фотосистемы II, и те из них, в которых он не работает, склонны к быстрому и необратимому фотоингибированию[21].
Марганцевый механизм
Фотон поглощённый ионами марганца в марганцевом кластере водоокисляющего комплекса вызывает его инактивацию. Дальнейшее торможение реакций переноса электронов происходит по механизму ингибирования донорной стороны. В пользу этого механизма говорят данные о спектре действия фотоингибирования[12].
Механизм синглетного кислорода
Ингибирование фотосистема II вызвано синглетным кислородом, который образуется либо слабо сопряжёнными молекулами хлорофилла[22] или восстановленными цитохромами и железосерными центрами[23].
Кинетика и спектр действия
Если измерить кинетику фотоингибирования в листьях, цианобактериях или водорослях, окрашенных линкомицином или в изолированной тилакоидной мембране в которой цикл репарации не нарушает кинетику, то фотоингибирование будет описывается уравнением реакции первого порядка. Данные от группы С. У. Чоу указывают на то, что в листьях перца (Capsicum annuum) вместо реакция первого имеется псевдоравновесие, даже когда цикл репарации заблокирован. Этому факту можно дать объяснение, если предположить, что ингибирование части ФСII защищает оставшиеся активные реакционные центры от повреждений[24]. Как видимый, так и ультрафиолетовый свет вызывают фотоингибирование, причём действие первого оказывается значительно более разрушительным[12][23][25]. Некоторые исследователи считают, что ультрафиолет и видимый свет индуцируют фотоингибирование по двум разным механизмам[26], в то время как другие подчёркивают сходство между реакциями ингибирования, протекающими под воздействием разных диапазонов длин волн[12][13].
Цикл репарации фотосистемы II
Под воздействием света в фотосинтезирующих организмах на свету постоянно происходит фотоингибирование и поэтому они должны постоянно исправлять возникающие повреждения[8]. Цикл репарации фотосистемы II происходит в хлоропластах и цианобактериях: он заключается в деградации старого и синтезе нового D1 белка ФСII с последующей сборки реакционного центра. Остальные белки фотосистемы рециркулируют и вновь используются для сборки ФСII. Из-за быстрой репарации наиболее реакционные центры ФСII не фотоингибируется даже если растение растёт на ярком свету. Однако, экологические стрессы, например, экстремальные температуры, засоление и засуха, ограничивают поступлении диоксида углерода в цикл фиксации углерода, что уменьшает скорость репарации ФСII[27].
При исследовании фотоингибирования цикл репарации часто блокируют антибиотиками (линкомицин или левомицетин), которые останавливают синтез белка в хлоропласте. Синтез белка происходит только в не разрушенном образце, так что линкомицин не нужен, когда фотоингибирование измеряется в изолированных мембранах[27].
Защитные механизмы
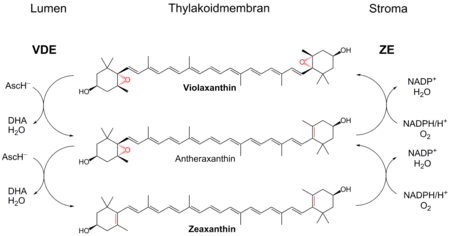
Растения имеют механизмы, защищающие их от неблагоприятного воздействия яркого света. Наиболее изучен биохимический защитный механизм нефотохимического тушения энергии возбуждения[28]. Фотоингибирование, вызванное видимым светом, у мутанта Arabidopsis thaliana без нефотохимического тушения происходит на ~25 % быстрее у дикого типа. Очевидно также, что поворот или складывания листьев, как это происходит, например, у кислицы в ответ на воздействие яркого света, защищает от фотоингибирования.
Измерение
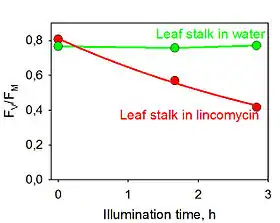
Фотоингибирование можно измерить в изолированных тилакоидных мембранах или их подфракциях, или же в интактных клетках цианобактерий путём измерения скорости выделения кислорода в условиях полного насыщения светом и в присутствии искусственных акцепторов электронов (реактивы хилла).
Степень фотоингибирования в интактных листьях можно померить используя флуориметр для измерения отношения вариабельного и максимального значение флуоресценции хлорофилла a (FV/FM)[16]. Из этого показателя можно получит значение степени фотоингибрования, поскольку значительная часть энергии высвобождается в виде флуоресценции хлорофилла а в условиях, когда много возбужденных электронов от ФСII не переносятся на акцептор, а вместо этого переходят обратно в основное энергетическое состояние.
Перед измерением отношения FV/FM, листья должны инкубироваться в темноте в течение не менее 10 минут, а желательно дольше, для того чтобы снять нефотохимическое тушение.
Вспышки света
Фотоингибрование также можно индуцировать короткими вспышками света, используя импульсный лазера или ксеноновые лампы-вспышки. При очень коротких вспышках эффективность фотоингибирования зависит от паузы между вспышками[11]. Такую зависимость объяснили тем, что вспышки вызывают разделение зарядов в ФСII с последующим производством синглетного кислорода. Интерпретация была подвергнута критике, поскольку эффективность фотоингибирования, вызванного вспышками ксеноновой лампы всё же зависит от энергии вспышки даже при таких сильных вспышках, что они насыщают реакцию разделения зарядов[12].
Динамическое фотоингибирование
Некоторые исследователи предпочитают определять термин «фотоингибирование» так, что под ним подразумеваются все реакции, которые снижают квантовый выход фотосинтеза, когда растение подвергается воздействию света[29][30]. Термин «динамическое фотоингибирование» включает в себя явления, которые обратимо снижают уровень фотосинтеза на свету, а термины «фотоповреждения» или «необратимое фотоингибирование» охватывают понятие фотоингибирования, связанное непосредственно с разрушительным действием света. Основной механизм динамического фотоингибирования — нефотохимическое тушение энергии возбуждения, поглощённой ФСII. Динамические фотоингибирование — это акклиматизация к условиям яркого света, а не вызванные светом повреждения и поэтому «динамическое фотоингибирование» на самом деле защищает растения от «фотоингибирования».
Экология фотоингибирования
Фотоингибирование может привести к обесцвечиванию кораллов[27].
См. также
Примечания
- Kok B. On the inhibition of photosynthesis by intense light (англ.) // Biochimica et Biophysica Acta : journal. — 1956. — Vol. 21, no. 2. — P. 234—244. — doi:10.1016/0006-3002(56)90003-8. — PMID 13363902.
- Jones LW & Kok B. Photoinhibition of Chloroplast Reactions. I. Kinetics and Action Spectra (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 1966. — Vol. 41, no. 6. — P. 1037—1043. — doi:10.1104/pp.41.6.1037. — PMID 16656345.
- Kyle D.J., Ohad I., Arntzen C.J. Membrane protein damage and repair: Selective loss of a quinone-protein function in chloroplast membranes (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1984. — Vol. 81, no. 13. — P. 4070—4074. — doi:10.1073/pnas.81.13.4070. — PMID 16593483.
- Callahan F.E., Becker DW & Cheniae G.M. Studies on the Photoactivation of the Water-Oxidizing Enzyme: II. Characterization of Weak Light Photoinhibition of PSII and Its Light-Induced Recovery (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 1986. — Vol. 82, no. 1. — P. 261—269. — doi:10.1104/pp.82.1.261. — PMID 16665003.
- Jegerschöld C., Virgin I & Styring S. Light-dependent degradation of the D1 protein in photosystem II is accelerated after inhibition of the water splitting reaction (англ.) // Biochemistry : journal. — 1990. — Vol. 29, no. 26. — P. 6179—6186. — doi:10.1021/bi00478a010. — PMID 2207066.
- Vass I., Styring S., Hundal T., Koivuniemi M., Aro E-M & Andersson B. Reversible and irreversible intermediates during photoinhibition of photosystem II: Stable reduced QA species promote chlorophyll triplet formation (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 89, no. 4. — P. 1408—1412. — doi:10.1073/pnas.89.4.1408.
- Hideg É, Kálai T., Hideg K & Vass I. Photoinhibition of photosynthesis in vivo results in singlet oxygen production detection via nitroxide-induced fluorescence quenching in broad bean leaves (англ.) // Biochemistry : journal. — 1998. — Vol. 37, no. 33. — P. 11405—11411. — doi:10.1021/bi972890. — PMID 9708975.
- Aro E-M, Virgin I & Andersson B. Photoinhibition of Photosystem II – inactivation, protein damage and turnover (англ.) // Biophysica et Biochimica Acta : journal. — 1993. — Vol. 1143, no. 2. — P. 113—134. — doi:10.1016/0005-2728(93)90134-2.
- Bailey S., Thompson E., Nixon P.J., Horton P., Mullineaux C.W., Robinson C & Mann N.H. A critical role for the Var2 FtsH homologue of Arabidopsis thaliana in the Photosystem II repair cycle in vivo (англ.) // Journal of Biological Chemistry : journal. — 2002. — Vol. 277, no. 3. — P. 2006—2011. — doi:10.1074/jbc.M105878200. — PMID 11717304.
- Tyystjärvi, E & Aro, E-M. The rate constant of photoinhibition, measured in lincomycin-treated leaves, is directly proportional to light intensity (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1996. — Vol. 93, no. 5. — P. 2213—2218. — doi:10.1073/pnas.93.5.2213. — PMID 11607639.
- Keren N., Berg A., van Kan PJM, Levanon H & Ohad I. Mechanism of photosystem II photoinactivation and D1 protein degradation at low light: The role of back electron flow (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1997. — Vol. 94, no. 4. — P. 1579—1584. — doi:10.1073/pnas.94.4.1579. — PMID 11038602.
- Hakala M., Tuominen I., Keränen M., Tyystjärvi T & Tyystjärvi E. Evidence for the role of the oxygen-evolving manganese complex in photoinhibition of Photosystem II (англ.) // Biochimica et Biophysica Acta (BBA) - Bioenergetics : journal. — 2005. — Vol. 1706, no. 1—2. — P. 68—80. — doi:10.1016/j.bbabio.2004.09.001. — PMID 15620366.
- Ohnishi N., Allakhverdiev S.I., Takahashi S., Higashi S., Watanabe M., Nishiyama Y & Murata N. Two-Step Mechanism of Photodamage to Photosystem II: Step 1 Occurs at the Oxygen-Evolving Complex and Step 2 Occurs at the Photochemical Reaction Center (англ.) // Biochemistry : journal. — 2005. — Vol. 44, no. 23. — P. 8494—8499. — doi:10.1021/bi047518q. — PMID 15938639.
- Tyystjärvi T., Tuominen I., Herranen M., Aro E-M, Tyystjärvi E. Action spectrum of psbA gene transcription is similar to that of photoinhibition in Synechocystis sp. PCC 6803 (англ.) // FEBS Letters : journal. — 2002. — Vol. 516, no. 1—3. — P. 167—171. — doi:10.1016/S0014-5793(02)02537-1. — PMID 11959126.
- Nishiyama Y., Allakhverdiev SI & Murata N. Inhibition of the repair of Photosystem II by oxidative stress in cyanobacteria (англ.) // Drugs : journal. — Adis International, 2002. — Vol. 516, no. 1—3. — P. 167—171. — doi:10.1007/s11120-004-6434-0. — PMID 16049747.
- Tyystjärvi E. Photoinhibition of Photosystem II and photodamage of the oxygen-evolving manganese cluster (англ.) // Coordination Chemistry Reviews : journal. — 2008. — Vol. 252, no. 3—4. — P. 361—376. — doi:10.1016/j.ccr.2007.08.021.
- Krieger-Liszkay A., Fufezan C & Trebst A. Singlet oxygen production in photosystem II and related protection mechanism (англ.) // Drugs : journal. — Adis International, 2008. — Vol. 98, no. 1—3. — P. 551—564. — doi:10.1007/s11120-008-9349-3. — PMID 18780159.
- Sonoike K. Photoinhibition of photosystem I: Its physiological significance in the chilling sensitivity of plants (англ.) // Plant and Cell Physiology : journal. — 1996. — Vol. 37. — P. 239—247. — doi:10.1093/oxfordjournals.pcp.a028938.
- Eckert H.J., Geiken B., Bernarding J., Napiwotzki A., Eichler HJ & Renger G. Two sites of photoinhibition of the electron-transfer in oxygen evolving and Tris-treated PS-II membrane-fragments from spinach (англ.) // Drugs : journal. — Adis International, 1991. — Vol. 27, no. 2. — P. 97—108. — doi:10.1007/BF00033249.
- Nishiyama Y., Allakhverdiev SI & Murata N. A new paradigm for the action of reactive oxygen species in the photoinhibition of photosystem II (англ.) // Biophysica et Biochimica Acta - Bioenergetics : journal. — 2006. — Vol. 1757, no. 7. — P. 742—749. — doi:10.1016/j.bbabio.2006.05.013. — PMID 16784721.
- Anderson J.M., Park Y-I & Chow W.S. Unifying model for the photoinactivation of Photosystem II in vivo: a hypothesis (англ.) // Drugs : journal. — Adis International, 1998. — Vol. 56. — P. 1—13. — doi:10.1023/A:1005946808488.
- Santabarbara S., Cazzalini I., Rivadossi A., Garlaschi F.M., Zucchelli G & Jennings R.C. Photoinhibition in vivo and in vitro involves weakly coupled chlorophyll-protein complexes (англ.) // Photochemistry and Photobiology : journal. — 2002. — Vol. 75, no. 6. — P. 613—618. — doi:10.1562/0031-8655(2002)0750613PIVAIV2.0.CO2.
- Jung J & Kim H-S. The chromophores as endogenous sensitizers involved in the photogeneration of singlet oxygen in spinach thylakoids (англ.) // Photochemistry and Photobiology : journal. — 1990. — Vol. 52, no. 5. — P. 1003—1009. — doi:10.1111/j.1751-1097.1990.tb01817.x.
- Lee H.Y., Hong YN & Chow W.S. Photoinactivation of photosystem II complexes and photoprotection by non-functional neighbours in Capsicum annuum L. leaves (англ.) // Planta : journal. — 2001. — Vol. 212, no. 3. — P. 332—342. — doi:10.1007/s004250000398.
- Sarvikas P., Hakala M., Pätsikkä E., Tyystjärvi T & Tyystjärvi E. Action spectrum of photoinhibition in leaves of wild type and npq1-2 and npq4-1 mutants of Arabidopsis thaliana (англ.) // Plant and Cell Physiology : journal. — 2006. — Vol. 47, no. 3. — P. 391—400. — doi:10.1093/pcp/pcj006. — PMID 16415063.
- Sicora C., Mate Z & Vass I. The interaction of visible and UV-B light during photodamage and repair of Photosystem II (англ.) // Drugs : journal. — Adis International, 2003. — Vol. 75, no. 2. — P. 127—137. — doi:10.1023/A:1022852631339. — PMID 16245083.
- Takahashi S & Murata N. How do environmental stresses accelerate photoinhibition (англ.) // Trends in Plant Science : journal. — Cell Press, 2008. — Vol. 13, no. 4. — P. 178—182. — doi:10.1016/j.tplants.2008.01.005. — PMID 18328775.
- Krause GH & Jahns P (2004) «Non-photochemical energy dissipation determined by chlorophyll fluorescence quenching: Characterization and function» in Papageorgiou GC & Govindjee (eds.
- Powles S.B. Photoinhibition of photosynthesis induced by visible light (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 1984. — Vol. 35. — P. 15—44. — doi:10.1146/annurev.pp.35.060184.000311.
- Hall DO & Rao K. K. Photosynthesis (неопр.). — Cambridge University Press, Cambridge, 1999. — ISBN 978-0-521-64497-6.
Литература
- Alves, P L da C A., Magalhães A C N., Barja P R. The phenomenon of photoinhibition of photosynthesis and its importance in reforestation (англ.) // The Botanical Review : journal. — 2002. — Vol. 68, no. = 2. — P. 193—208. — doi:10.1663/0006-8101(2002)068[0193:TPOPOP]2.0.CO;2. Архивировано 21 июля 2011 года.
- Vass I & Cser K. Janus-faced charge recombinations in photosystem II photoinhibition (англ.) // Trends in Plant Science : journal. — Cell Press, 2009. — Vol. 14, no. 4. — P. 200—205. — doi:10.1016/j.tplants.2009.01.009. — PMID 19303349.
- Mohanty P., Allakhverdiev SI & Murata N. Application of low temperatures during photoinhibition allows characterization of individual steps in photodamage and the repair of photosystem II (англ.) // Drugs : journal. — Adis International, 2007. — Vol. 94, no. 2—3. — P. 217—224. — doi:10.1007/s11120-007-9184-y. — PMID 17554634.
- Telfer A. Too much light? How beta-carotene protects the photosystem II reaction centre (англ.) // Photochemical & Photobiological Sciences : journal. — 2005. — Vol. 4, no. 12. — P. 950—956. — doi:10.1039/b507888c. — PMID 16307107.
- Adir N., Zer H., Shochat S & Ohad I. Photoinhibition - a historical perspective (англ.) // Drugs / Govindjee; Beatty, J. Thomas; Gest, Howard; Allen, John F.. — Adis International, 2003. — Vol. Advances in Photosynthesis and Respiration, no. 1—3. — P. 343—370. — ISBN 1-4020-3323-0. — doi:10.1007/1-4020-3324-9. — PMID 16228592.
- Niyogi K.K. Photoprotection revisited: Genetic and molecular approaches (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 1999. — Vol. 50. — P. 333—359. — doi:10.1146/annurev.arplant.50.1.333. — PMID 15012213.