Ксантофилловый цикл
Ксантофилловый цикл, или виолаксантиновый цикл, в случае высших растений, выполняет функцию защиты фотосинтетического аппарата от избытка энергии при повышенной инсоляции. Он позволяет избежать фотоингибирования, значительно увеличив нефотохимическое тушение. Цикл включает в себя ферментативные взаимопревращения между виолаксантином и зеаксантином (промежуточный продукт — антероксантин)[1].
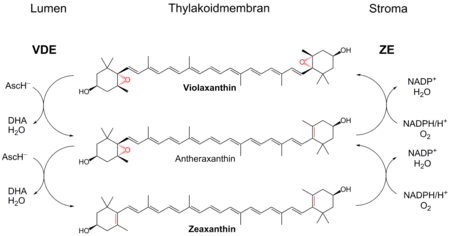
Взаимопревращение ксантофиллов
Виолаксантиновый цикл протекает в минорных субъединицах светособирающего комплекса фотосистемы II (белки CP29, CP26, CP23, CP22 и др.). При высокой интенсивности света вследствие активной работы ЭТЦ фотосинтеза происходит закисление люмена тилакоидов. При падении pH до 5,0 активируются фермент деэпоксидаза, которая с люменальной стороны мембраны осуществляет восстановление эпоксидных групп виолоксантина, используя в качестве восстановителя аскорбиновую кислоту. Двукратное восстановление приводит к образованию зеаксантина, который и выполняет фотопротекторную функцию. При снижении интенсивности освещения, начинает преобладать обратная реакция катализируемая эпоксидазой, расположенной со стромальной стороны мембраны с pH оптимумом 7,5. Для введения эпоксидных групп необходим молекулярный кислород и восстановитель (НАДФН). В результате образуется виолаксантин, который может выполнять роль светособирающего пигмента[2].
Антенная функция
Ксантофиловый цикл высших растений и водорослей играет важную роль в регуляции перераспределения энергии света между виолаксантином, зеаксантином и хлорофиллом а. Роль фотопротектора в цикле выполняет зеаксантин, содержащий сопряжённую систему из 11 двойных связей (у виолоксантина всего 9 сопряженных связей). Увеличение сопряжения приводит к снижению энергии первого синглетного возбужденного уровня пигмента. Для виолоксантина характерно поглощение при 657 нм, а для зеаксантина при 704 нм (поглощение хлорофилла при 680 нм). Таким образом, поскольку энергия возбужденного состояния зеаксантина ниже энергии возбужденного состояния хлорофилла, возможен прямой синглет-синглетный перенос энергии от возбуждённого хлорофилла (S1) к зеаксантину. Поглощение избыточной энергии возбуждения хлорофилла зеаксантином защищает фотосинтетический аппарат. С другой стороны энергия возбуждённого S1-состояния молекулы виолоксантина выше чем у хлорофилла, и поэтому виолоксантин может стать донором энергии для хлорофилла. В результате виолоксантин играет роль антенны, собирающей фотоны и переносящей их на хлорофилл.
Таким образом, при низком освещении ксантофиллы действуют преимущественно как светособирающие пигменты, а при высоком помогают рассеять избыточную энергию и защитить растение от фотоингибирования. Этот же механизм помогает подготовить растение к смене дня и ночи.
Защитная функция
Длинные молекулы каротиноидов с большим количеством сопряжённых связей особенно эффективно преобразуют энергию возбуждения в тепловую через внутреннюю конверсию. Благодаря большому количеству сопряжённых двойных связей, которые могут достаточно значительно изменять свою длину, молекулы каротиноидов совершают постоянные колебания сжимания разжимания наподобие гармошки. Такое поведение обеспечивает быструю диссипацию энергии в тепло. Это же свойство позволяет ксантофиллам и каротиноидам эффективно тушить активные, синглетные формы кислорода и возбуждённого, триплетного хлорофилла[3].
Кроме того, конформационные изменения в молекуле виолоксантина протекающие при деэпоксидации приводят к тому, что зеаксантин легче образует агрегаты. Образование агрегатов зеаксантина в минорных белках ССК II приводит к блокированию миграции энергии от внешних антенн ССКII к реакционному центру ФСII и тепловой диссипации энергии возбуждения. Известно, что при ярком солнечном освещении растения рассеивают в форме тепла от 50 до 70 % энергии поглощённых квантов. Поэтому именно тепловая диссипация энергии играет ключевую роль в защите фотосинтетического аппарата (считается, что вклад реакции Мелера и фотодыхания значительно меньше)[4].
При исследовании шпината было обнаружено, что при ярком освещении увеличение концентрации зеаксантина превосходит снижение концентрации виолоксантина. Возможно, такие условия побуждают растения к более активному синтезу зеаксантина из β-каротина[5].
Действие на мембрану
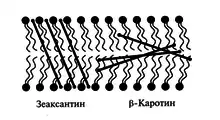
Также зеаксантин выполняет важную структурную роль в мембранах тилакоидов. При повышенном освещении ксантофиллы распределяются между светособирающими комплексами и липидным бислоем. Поскольку зеаксантин имеет две гидрофильные -OH группы, он располагается поперек бислоя, тем самым увеличивая вязкость мембраны. Увеличение вязкости мембраны благодаря зеаксантину и некоторым терпеноидам (α-токоферол) снижает её проницаемость для кислорода и защищает липиды от перекисного окисления его активными формами[6].
Другие подобные циклы
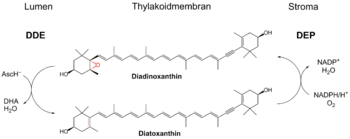
У диатомовых водорослей и динофлагеллят ксантофиловый цикл состоит из пигмента диадиноксантина, который в условиях избыточного освещения превращается в диатоксантин у диатомовых или диноксантин у динофлагеллят[7].
У некоторых высших растений (например из рода Инга[8]) кроме виолаксантинового цикла обнаружен дополнительный лютеиновый цикл. В ходе этого цикла происходит обратимое превращения лютеин-5,6-эпоксида на ярком свету в лютеин, что также по-видимому способствует защите фотосинтетического аппарата[9][10]. Этот цикл есть у некоторых таксономически далеко отстоящих групп, но он консервативен и сохраняется в пределах семейства и рода.
Примечания
- Falkowski, P. G. & J. A. Raven, 1997, Aquatic photosynthesis. Blackwell Science, 375 pp
- Taiz, Lincoln and Eduardo Zeiger. 2006. Plant Physiology. Sunderland, MA: Sinauer Associates, Inc. Publishers, Fourth edition, 764 pp
- Ермаков, 2005, с. 143.
- Ермаков, 2005, с. 145.
- Wright. The interrelationship between the lower oxygen limit, chlorophyll fluorescence and the xanthophyll cycle in plants (недоступная ссылка). Дата обращения: 6 января 2016. Архивировано 24 сентября 2019 года.
- Ермаков, 2005, с. 146.
- Jeffrey, S. W. & M. Vesk, 1997. Introduction to marine phytoplankton and their pigment signatures. In Jeffrey, S. W., R. F. C. Mantoura & S. W. Wright (eds.), Phytoplankton pigments in oceanography, pp 37-84. — UNESCO Publishing, Paris.
- MATSUBARA SHIZUE, KRAUSE G. HEINRICH, SELTMANN MARTIN, VIRGO AURELIO, KURSAR THOMAS A., JAHNS PETER, WINTER KLAUS. Lutein epoxide cycle, light harvesting and photoprotection in species of the tropical tree genus Inga // Plant, Cell & Environment. — 2008. — Апрель (т. 31, № 4). — С. 548—561. — ISSN 0140-7791. — doi:10.1111/j.1365-3040.2008.01788.x.
- García-Plazaola, JI. et al. (2007): The lutein epoxide cycle in higher plants: its relationships to other xanthophyll cycles and possible functions. In: Functional Plant Biology 34(9); 759—773; doi:10.1071/FP07095.
- Bungard, RA. et al. (1999): Unusual carotenoid composition and a new type of xanthophyll cycle in plants. In: Proc Natl Acad Sci USA 96(3); 1135—1139; PMID 9927706; PMC 15363
Литература
- Фотосинтез. Физиолого-экологические и биохимические аспекты / А.Т Мокроносов, В. Ф. Гавриленко, Т. В. Жигалова; под ред. И. П. Ермакова. — М. : «Академия», 2006. — 448 с. — ISBN 5-7695-2757-9
- Физиология растений / Под ред. И. П. Ермакова. — М.: Академия, 2005. — 634 с. — ISBN 978-5-7695-36-88-5.
- Хелдт Г. В. Биохимия растений. — М.: БИНОМ. Лаборатория знаний, 2011. — 471 с. — ISBN 978-5-94774-795-9.