Турухтан
Турухта́н[1] (лат. Philomachus pugnax) — птица из семейства бекасовых. Гнездится на травяных болотах и во влажных лугах в северной части Евразии. Перелётная птица, зимует в Западной и Южной Европе, Африке южнее Сахары, Южной Азии и Австралии[2]. На пролёте держится большими стаями, в местах зимовок нередко образует колонии до нескольких сотен тысяч птиц. Считается монотипичным видом, ближайшие родственные виды — грязовик и острохвостый песочник[3].
| Турухтан | ||||||||
|---|---|---|---|---|---|---|---|---|
 | ||||||||
| Научная классификация | ||||||||
| Международное научное название | ||||||||
| Philomachus pugnax (Linnaeus, 1758) | ||||||||
| Ареал | ||||||||
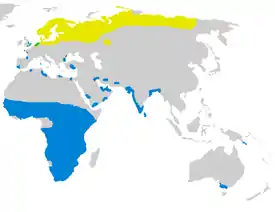 Гнездовой ареал Миграции Круглогодично |
||||||||
| Охранный статус | ||||||||
| ||||||||

Довольно стройный длинноногий кулик, внешне напоминающий крупного песочника. Самцы и самки заметно отличаются друг от друга размерами, а весной также окрасом и деталями оперения. В брачном наряде самцы имеют ярко-окрашенный пышный воротник перьев на груди, своего рода «уши» и мелкие выросты в виде бородавок на оголённой коже головы. В остальное время года самцы выделяются лишь более крупными размерами; оба пола в этот период окрашены в серовато-бурые тона с беловатым брюхом. Молчаливая птица, лишь изредка издающая негромкие звуки. Несмотря на огромное разнообразие расцветок, различают 3 типа весеннего оперения самцов, по которому определяют отличную друг от друга стратегию размножения[4]. В общем случае самцы выясняют отношения между собой на току, который время от времени посещают самки. Яйца откладывают один раз в сезон, в полной кладке обычно 4 яйца. Самки насиживают и ухаживают за потомством в одиночку[5]. Кормится вдоль илистых берегов водоёмов. В сезон размножения употребляет в пищу преимущественно животные корма, на зимовках — большей частью растительные. Питается мелкими беспозвоночными и семенами трав, зерновыми культурами. Обычный, местами многочисленный вид.
Систематическое положение и название
Турухтан выделен в отдельный род Philomachus, который в свою очередь относится к достаточно обширному семейству бекасовых. Сотрудники британского Университета Бата, построившие филогенетическое дерево всех куликов, пришли к выводу, что ближайшими родственниками турухтана являются грязовик и острохвостый песочник[3]. Несмотря на цветовое разнообразие самцов, на всём протяжении ареала не отмечено каких-либо морфологических особенностей, характерных только для данной местности[6].
Первое описание турухтана появилось в «Системе природы» — фундаментальной работе шведского врача и натуралиста Карла Линнея, положившей начало научной систематике организмов. Автор поместил птицу в один ряд с улитами и присвоил ей биноминальное название Tringa pugnax[7]. Современное научное именование птицы закрепилось после того, как в 1804 году немецкий натуралист Блазиус Меррем в журнале Allgemeine Literatur-Zeitung специально для турухтана описал монотипичный род Philomachus[8]. Как родовое, так и видовое названия ссылаются на агрессивное поведение самцов во время токовых игр. Слово Philomachus является производным из двух древнегреческих слов — φίλος («любитель») и μάχη («битва»)[9][10], слово pugnax имеет латинское происхождение и буквально переводится как «воинственный»[10][11]. Русскоязычное название, по всей видимости, образовано в результате звукоподражания[12].
Описание
Внешний вид

Среднего размера кулик, крупнее чернозобика. Определение в полевых условиях обычно не вызывает затруднений. Необычно маленькая голова на фоне упитанного туловища, вытянутая шея и узкий, слегка изогнутый книзу клюв хорошо выделяют турухтана среди других куликов, в том числе схожих по окрасу и образу жизни[13]. В сравнении с другими некрупными куликами, в полёте турухтан делает более медленные и глубокие взмахи крыльями, подолгу парит. Кроме того, он хорошо узнаётся по белой размытой полосе по верхнему краю крыла и белым пятнам по краям хвоста[13]. Крылья заострённые и несколько уже, чем у других куликов, хвост слегка закруглённый. Между основаниями среднего и внешнего пальцев развита небольшая перепонка[14]. Ноги довольно длинные; у взрослых птиц оранжевые либо почти красные, у двухлетних серовато-жёлтые (иногда с серыми пятнами), у более молодых бурые, зеленовато- или тёмно-серые[15].

Самец и самка обладают ярко выраженными отличиями друг от друга, в связи с чем охотники в XIX веке иногда даже принимали их за разных птиц[16]. Самец выглядит существенно крупнее и массивнее самки: его длина 28—33 см, размах крыльев 50—58 см, масса 120—310 г, в то время как аналогичные характеристики у самки 22—27 см, 46—52 см и 70—150 г соответственно[5]. Диморфизм также проявляется в окраске и рисунке оперения в период размножения. Зимой обе птицы окрашены примерно одинаково: однообразный серовато-бурый верх и беловатый низ без каких-либо резких отметин. В феврале—марте после частичной замены перьев у самца передняя часть спины, плечи, кроющие крыла и рулевые заметно темнеют; на этих почти чёрных участках оперения появляется лиловый металлический отлив[17].
В апреле самец приобретает окончательный брачный наряд, своим многообразием нехарактерный для куликов. По бокам головы и на шее вырастают удлинённые перья, известные как «уши» и «воротник». На оголённых участках головы развиваются так называемые «бородавки» — кожистые складки красного, оранжевого или жёлтого цветов. Другие неоперённые участки тела также приобретают яркие краски: клюв становится жёлтым, оранжевым либо розовато-серым с тёмной вершиной, ноги — жёлтые с красноватым, зеленоватым или буроватым оттенком[18][19]. Окраска воротника, а также груди, спины и крыльев, может быть настолько разносторонней, что в одной стае редко попадаются самцы, похожие друг на друга. Попадаются белые, бледно-охристые, ярко- и тёмно-рыжие, оливковые, коричневые, чёрно-синие с зеленоватым или пурпурным отливом, чёрные кулики, а также с любой комбинацией этих тонов. Другие части тела также пёстрые, но выглядят одинаково у всех самцов — задняя часть спины, надхвостье и центральные кроющие хвоста серовато-бурые, брюхо и крайние кроющие хвоста белые[14][20]. По мнению орнитологов, такое разнообразие нарядов помогает молчаливым от природы птицам распознавать друг друга во время брачных церемоний[21].
По окончании летней линьки у самцов исчезают перьевые украшения и кожистые бородавки, клюв и ноги теряют яркие краски и бледнеют. Оперение приобретает покровительственные тона — серовато-бурые в верхней части тела и беловато-серые с тёмными крапинами снизу и по бокам[13]. Самки в любое время года имеют наряд, схожий с осенним у самца, однако в брачный сезон он несколько более тёмный и пёстрый. Молодые птицы похожи на взрослых зимой, однако имеют отчётливый чешуйчатый рисунок из рыжих и охристых каёмок тёмных перьев на спине и крыльях, а также лёгкий охристый оттенок на брюхе[15][13].
Линька
_near_Hodal_W_IMG_6498.jpg.webp)
Взрослый турухтан в течение года линяет два или три раза, в зависимости от пола и возраста. Послебрачная линька, в ходе которой происходит полная замена оперения, начинается в июле и продолжается до сентября или октября, изредка до декабря[18]. С конца зимы до первой декады апреля происходит частичная замена перьев у обоих полов, после которой самка приобретает окончательный брачный вид, в то время как самец промежуточный — спустя месяц-полтора только что образовавшиеся тёмные перья, равно как и оставшиеся зимние, вновь выпадают и заменяются более пышными и яркими (феномен, также характерный для малого веретенника)[22]. Двухгодовалые самцы, которые выглядят мельче и слабее своих старших собратьев, приступают к линьке несколько позже. По мнению белорусских и польских орнитологов, такая задержка позволяет птицам использовать меньше энергии во время дальнего перелёта[23].
У взрослых самцов и большинства взрослых самок летняя линька начинается ещё до отлёта на юг, однако окончательная смена пера происходит уже в местах зимовок. В Кении самцы линяют на 3—4 недели раньше самок и приобретают зимний наряд ещё до начала декабря, в то время как у их партнёрш этот процесс продолжается до конца декабря — начала января. Молодые в первый год жизни линяют с сентября по ноябрь, а к последующей весенней линьке приступают одновременно со взрослыми[24].
Отличия от схожих видов

Самца в брачном наряде достаточно сложно спутать с какой-либо другой птицей, однако в других случаях определение вида может потребовать специальных сравнительных характеристик. В зимнем наряде турухтан по форме тела, рисунку оперения, окраске клюва и ног несколько напоминает травника. Однако у травника клюв прямой и более длинный, задняя часть спины и надхвостье белые. Остальной верх травника также более светлый и монотонно окрашенный, в то время как у турухтана центральная часть пера более тёмная[25]. Молодой острохвостый песочник величиной лишь немногим уступает самке турухтана и так же, как и молодой турухтан, имеет рыжевато-охристую грудь. Однако он выглядит более упитанным, имеет вытянутую (не округлую) форму головы, короткие ноги и короткий клюв. Над глазом острохвостого песочника хорошо развита белая полоса[26]. Канадский песочник так же, как и острохвостый, выглядит более упитанным. К тому же этот вид значительно крупнее и длинноклювее и выделяется чешуйчатым рисунком на спине[27]. У щёголя и фифи в зимнем наряде отсутствуют рыжие тона в оперении. Кроме того, у щёголя прямой и более длинный клюв, а фифи заметно мельче. Меньшими размерами и более короткими ногами отличаются краснозобик и чернозобик[28].
Голос
Почти беззвучная птица. Лишь гнездящаяся самка иногда издаёт негромкий крик, который можно охарактеризовать как короткое глухое кряканье или кряхтенье, напоминающее хорканье вальдшнепа. Изредка подобный звук издают самцы на току[5][29].
Распространение
Гнездовой ареал

Типичная перелётная птица, гнездится на болотах и влажных травяных лугах в северной части Евразии от Британских островов и Скандинавии к востоку до низовьев Колымы и Анадыря[30][31]. Зимует в тропиках к югу от гнездового ареала, в основном в Африке. В Европе и Азии турухтан распространён на большей части лесной и тундровой зон от Великобритании и северо-западной Франции на восток до Чукотки и побережий Берингова и Охотского морей. В Европе птицы поднимаются к северу вплоть до арктического побережья, в районе Ямала и Гыданского полуострова до 72° с. ш., на Таймыре до 73° с. ш., восточнее вновь до берегов Северного Ледовитого океана[2].
Наибольшей численности, более 1 млн пар, гнездится на территории России[32]. Другие крупные скопления птиц отмечены в Северной Европе — в Швеции (61 тыс. пар), Финляндии (39 тыс. пар) и Норвегии (14 тыс. пар). Общая численность турухтанов, гнездящихся в узкой прибрежной полосе от Великобритании и Франции на восток до Дании, Германии и Польши, не превышает 2 тыс. пар[31]. Нижняя граница гнездового ареала во многом условна, поскольку кулик иногда селится значительно южнее тундры и полосы северной тайги, где наблюдается наибольшая концентрация этих птиц[5]. Ведущий советский орнитолог Елизавета Козлова в начале 60-х годов XX века приводила свидетельства кладок яиц в Прибалтике, северных областях Украины, Псковской, Орловской и Новгородской областях, однако это были единичные находки, не приводившие к образованию постоянных токовищ[33]. Лео Степанян указывает на 54-ю параллель как приблизительную южную границу гнездовий, что соответствует таким городам, как Смоленск, Рязань и Ульяновск[2]. В Сибири основная область распространения — типичные и кустарниковые тундры, а также лесотундра. Южнее распространение спорадическое и не постоянное: кладки находили в долине реки Хобда (Актюбинская область, Казахстан), в Минусинской котловине, окрестностях села Сон (Хакасия)[33][34]. Восточнее турухтаны доказано гнездятся к югу до южного Байкала, дельты Селенги и северного побережья Охотского моря. Кроме того, небольшие изолированные участки отмечены на Украине в границах Житомирской, Киевской, Полтавской и Черниговской областей, в долине верхнего течения Оки, в среднем течении Урала, в среднем течении Илека, в бассейне Хобды, в промежутке между Барнаулом и Семипалатинском[2].
Зимний ареал
_W_IMG_6570.jpg.webp)
На большей части территории турухтан — типичный дальний мигрант. Так, гнездящиеся в низовьях Колымы птицы ежегодно совершают перелёты протяжённостью до 30 тыс. км, следуя через Северную Европу к водоёмам Западной Африки[35]. На пролёте турухтаны держатся большими стаями, численность птиц в которых может достигать сотен и тысяч особей. В местах зимовок они также нередко образуют огромные и плотные поселения: например, в дельте Сенегала ежегодно скапливается более полумиллиона птиц, а в отдельные годы их число может достигать миллиона[36]. Основные зимние колонии образуются в полосе Сахель, области Африки южнее Сахары — долинах рек Сенегал и Нигер, на озере Чад, в равнинной местности южнее города Томбукту (Мали), в промежутке между Эритреей и Замбией[36]. За пределами Африки зимует значительно меньшее количество птиц, периодически колонии возникают вдоль атлантического и средиземноморского побережья Европы, в Мьянме, южном Китае[37], Новой Гвинее и спорадически в южной Австралии[38].
В Великобритании и примыкающих к ней районах материковой Европы гнездовой и зимний ареалы пересекаются, и на этой территории турухтанов можно наблюдать в течение всего года[32]. Кроме того, часть не гнездящихся птиц весной не отлетает на север и проводит лето в тёплом тропическом климате. Случайные залёты этих куликов зарегистрированы на североамериканском континенте — на Аляске (где они иногда приступают к размножению), в Канаде и некоторых штатах в основной части США. Кроме того, странствующие птицы отмечены в Исландии, Центральной и северной части Южной Америки, Мадагаскаре и Новой Зеландии[37][39]. Гнездящиеся птицы были зарегистрированы к югу от основного ареала в северном Казахстане, где обычно птицы делают остановку во время миграции[40][41].

Самцы, не принимающие участия в насиживании и ухаживании за потомством, покидают гнездовья в конце июня или начале июля. Самки отлетают позже в июле, а набравшие вес птенцы в августе — сентябре[42]. Самцы также склонны зимовать в более северных широтах, чем самки, и по этой причине проделывают более короткий путь: например, практически все зимующие в Британии турухтаны принадлежат к мужскому полу[43], тогда как в более жаркой Кении большинство — к женскому[24]. Такое дифференцированное разделение характерно для многих перелётных видов птиц и имеет своё объяснение. Это и уменьшение конкуренции за доступ к корму, и возможность самцам как можно раньше вернуться к местам гнездовий, что увеличивает их шансы на успешное воспроизводство. К тому же самцы турухтана крупнее самок и, следовательно, легче переносят прохладную погоду[43].
На весеннем пролёте птицы всегда придерживаются традиционных маршрутов и на отдых останавливаются в определённых местах. Специалисты отмечают, что окольцованные турухтаны из года в год встречаются на одних и тех же стоянках. Теоретические расчёты показывают, что они останавливаются на отдых и подкормку раньше, чем позволяет их средний вес, что говорит об определённой стратегии при перелёте с использованием излюбленных мест для отдыха[42]. Накопленный подкожный жир у птиц используется в качестве «горючего», однако они, в отличие от млекопитающих, используют липиды как основной источник физической энергии (включая миграцию), а для поддержания температуры тела в случае необходимости встряхиваются. Процесс окисления липидов пока остаётся малоизученным[35].
Места обитания
В гнездовой период населяет травянистые или мохово-травянистые болота, влажные луга, разнообразные, за исключением сухих, тундры[5]. В тундре обычно встречается в поймах рек, в тайге и лесостепи также и на плоских водоразделах[34]. Самки турухтанов отдают предпочтение сырым, нередко топким низинам, поросшим осокой, ситником и злаками, в то время как самцы, наоборот, выбирают сухие площадки, лужайки и моховые болота. Кормится вблизи стоячих луж и водоёмов с илистыми берегами[14]. На зимовках птицы держатся на лугах вблизи водоёмов с пресной или солоноватой водой, на временных разливах, связанных с сезонными дождями, рисовых полях. В общем случае турухтаны не связаны напрямую с морскими побережьями, однако на крайнем юге Африки встречаются в лагунах и вдоль илистых берегов, заливаемых во время прилива и обнажаемых при отливе[36].
Питание

В отличие от большинства куликов, у турухтана развито чёткое сезонное разделение между животными и растительными кормами (кроме него, подобная характеристика известна у исландского песочника и большого веретенника)[36]. Летом основу питания составляют водные и наземные насекомые и их личинки: жуки, двукрылые (мухи, комары, долгоножки), водяные клопы, личинки ручейников, хирономид, шаровок и пр.[44] В меньшей степени питается моллюсками и ракообразными[36]. Кроме белковых кормов, иногда употребляет в пищу семена некоторых растений, таких как осока и камнеломка, а также луковицы горца живородящего[44].

В зимнее время на большей части территории преобладают растительные корма: семена трав, водных растений, осоки. В Западной Африке турухтаны в компании с обыкновенными горлицами нередко наносят значительный ущерб сельскому хозяйству, добывая семена на полях, засеянных рисом, пшеницей, просом и другими зерновыми культурами. Прибывая на зимовку в разгаре сезона дождей, птицы находят себе вдоволь растительной пищи с цветущих трав среди затопленной саванны. Позднее, когда наступает засуха и травы засыхают, турухтаны выискивают семена на голой потрескавшейся земле. Лишь на крайнем юге континента турухтаны зимуют вблизи морских побережий, где кроме всего прочего находят себе животную пищу — насекомых, моллюсков, ракообразных, пауков, червей, мелких лягушек и мелкую рыбу[36].
Активен как в светлое, так и в тёмное время суток[45]. Корм добывает с поверхности земли или растений, иногда погружает клюв в жидкую грязь или ищет пропитание на мелководье. В Восточной Африке иногда плавает подобно плосконосым плавунчикам и собирает пищу с поверхности воды[37]. Во время кормёжки турухтаны нередко взъерошивают перья спины, образуя некое подобие рыхлого хохолка на пояснице; такая же особенность отмечена у большого веретенника[13].
Размножение
К размножению приступает в возрасте двух лет[46]. Полигамен, пар не образует[47].
Брачные игры

Одна из особенностей турухтана, свойственных лишь немногим видам токующих птиц — брачное демонстративное поведение самцов направлено по отношению не к самкам, а к другим самцам. Кроме того, у самцов турухтанов необычно разнообразное брачное оперение (среди сотен птиц трудно подобрать двух совершенно одинаковых), переходящее по наследству[48][49]. Среди самцов выделяют три брачные формы: типичные территориальные самцы, подчинённые самцы с белым оперением воротника и очень редкий, открытый в 2006 году, вариант с самкоподобным оперением. К первому типу принадлежит подавляющее большинство, около 84 % от общего количества птиц. Они имеют красочно раскрашенные чёрные либо тёмно-рыжие воротники и неотлучно находятся на токовищах, где занимают небольшой, около одного метра в диаметре, пятачок поближе к центру (их иногда называют «резидентами»[14]). Эти самцы активно ухаживают за самками и выражают высокую степень агрессии по отношению к другим самцам. Постоянное токовище, на котором дежурят 5—20 территориальных самцов[32], может представлять собой возвышенность в тундре, поляну в пойме реки, сухой островок среди болота или даже просто ничем не приметное место[5].
Каждый территориальный самец может стать организатором общественного тока. Обнаружив в небе пролетающих самок либо стайку самцов, он зазывает их, всячески привлекая к себе внимание — подпрыгивает, кружится на одном месте и сигнализирует взмахами крыльев. Чем больше в одном месте собирается самцов, тем больше вероятности, что сюда прилетят и самки[50]. На току возбуждённые птицы машут крыльями, топорщат перья, кланяются, приседают, подпрыгивают, наскакивают друг на друга. Внешне это выглядит скорее как немой спектакль, нежели как выяснение отношений. Внезапно ток прекращается, и самцы застывают в затейливых позах либо приступают к поиску корма. Однако стоит мимо пролететь птице, хотя бы немного походящей на турухтана, как ток немедленно возобновляется[5][51].
Около 90 % территориальных самцов ежегодно посещают одно и то же токовище и находятся на нём до образования пары либо окончания сезона. Зная особенности поведения своих постоянных соперников, самец может определить их конкурентоспособность — таким образом в группе всегда возникает чёткая иерархия, в которой одни птицы доминируют над другими. Такое поведение уменьшает конфликтность и возможность получения травмы, что в свою очередь меньше отпугивает самок. Доминируемые самцы также получают пользу от постоянного использования участка, поскольку находятся на нём неотлучно и спокойно дожидаются, пока доминирующие первыми удаляются вслед за самкой[52].

Самцы более низкого ранга, количество которых составляет около 16 %, обладают белым либо пёстрым оперением воротника и не имеют постоянного токовища. Они посещают токовища резидентов, где держатся на периферии участка, и иногда перелетают с одного тока на другой, пытаясь спариться с самкой, пока самцы выясняют между собой отношения[21]. Такой тип поведения орнитологи называют клептостратегией размножения[47]. Территориальные самцы не сопротивляются присутствию гостей, поскольку хотя они и являются конкурентами за право обладания самкой, однако последние предпочитают посещать ток с бо́льшим количеством самцов[53][54]. Кроме того, самки отдают предпочтение участкам, вблизи которых есть подходящие для устройства гнезда места, в первую очередь сырые поросшие высокой травой лужайки[55].
Окрас оперения и особенности поведения отдельной особи сохраняются в течение жизни, поскольку определены простым генетическим полиморфизмом[56]. Изначально предполагали, что различие между двумя типами самцов связано с половым генетическим фактором, однако исследования показали, что в действительности локус (положение) гена, ответственного за демонстративное поведение, находится в аутосоме — парных хромосомах, не связанных с определённым полом. Это означает, что не только самцы, но и самки могут иметь две различные формы генов. В обычных условиях нетерриториальные самки никак не проявляют свой генотип, однако, если в их организм ввести имплантаты с мужским гормоном тестостероном, их территориальное поведение начинает походить на соответствующее поведение самцов с тем же набором генов[57].
Ранее полагали, что рацион питания птенцов в дальнейшем влияет на репродуктивную стратегию, поскольку подчинённые самцы в среднем выглядят мельче доминантов и обладают более светлым оперением. На самом деле наоборот — репродуктивная стратегия отражается на размерах птиц. Опыт показал, что при одинаковом количестве корма птенцы территориального типа набирают вес быстрее, чем птенцы субдоминантного. Последним не нужно тратить энергию на защиту территории, и они могут позволить себе больше времени на поиски корма. С другой стороны, подчинённые самцы часто меняют токовища, и затраченная на перелёты энергия влияет на их вес в сторону уменьшения[56].
Третий тип самцов, на постоянной основе имитирующих внешний вид самок, был описан в 2006 году — это первый известный случай подобной характеристики среди птиц. Около одного процента петушков имеют размеры, усреднённые между обычными самцами и самками. Они не надевают пышный брачный наряд, характерный для территориальных и подчинённых птиц, хотя в сравнении с ними имеют значительно более крупные семенники. Эти тайные самцы посещают токовища вместе с самками и буквально «воруют» спаривание у других самцов, когда готовые к соитию самки прижимаются к земле[4]. Самцы этого типа также подвержены весенней, добрачной линьке, когда зимние перья заменяются полосатыми. Однако если после неё у обычных самцов наступает следующий этап линьки — брачный, известный прежде всего длинными перьями воротника, то у самкоподобных этого не происходит. Специалисты полагают, что весенняя линька изначально являлась брачной до того, как в результате эволюции появились другие формы самцов[23]. Несмотря на имитацию внешнего вида самок, самцы третьего типа мигрируют в одной стае с другими самцами и проводят зиму вместе с ними[58].
-_acquiring_breeding_plumage-_near_Hyderabad_W_IMG_5022.jpg.webp)
С самцами третьего типа иногда пытаются спариться другие самцы — как территориальные, так и субдоминантные, хотя они способны распознать их как принадлежащих к своему полу[4]. На основании предварительных исследований полагают, что характеристика этого поведения заложена в отдельном «доминантном» гене[59]. Самки нередко отдают предпочтение третьему типу, но также и самцы имеют связь с ними чаще, чем с самками. Так же как и присутствие на токовище подчинённых самцов, дополнительные самцы третьего типа привлекают на него большее количество самок[60].
Не любое спаривание происходит обязательно на токовище. Самцы могут напрямую преследовать самку либо подкараулить её в хорошем месте, где та остановится на кормёжку. Петушки последовательно меняют тактику, однако отдают предпочтение току, если предыдущий день на нём был насыщенным либо часть самок уже приступила к гнездованию. В начале сезона в прохладную погоду самцы проводят больше времени в поисках пропитания, и по этой причине результативность тока остаётся низкой[61].
Полиандрия встречается у нескольких видов бекасовых, в том числе у песчанки, пятнистого перевозчика и щёголя[62]. Эта форма размножения также характерна для турухтана, причём в значительно большей степени, чем для других перечисленных птиц: более половины самок последовательно спаривается с двумя и более самцами. По мнению специалистов, такой «неразборчивости» способствует организация насиживания и выкармливания потомства, в которых самец участия не принимает[63].
Насиживание и птенцы


Гнездо — небольшое углубление в земле с выглаженными краями, хорошо укрытое в нависающей траве, на расстоянии не более 400 м от токовища[37][32]. Оно может быть расположено на кочке среди осокового болота, поросшей пушицей моховине, посреди сырого луга либо ином подобном месте[28]. Самка обильно выстилает гнездо прошлогодними листьями, стебельками трав и другим сухим растительным материалом. Как правило, гнездится в одиночку, однако иногда несколько птиц могут устроить гнездо в пределах прямой видимости от токовища[37][32]. Кладка, обычно состоящая из 4 яиц, приходится на середину марта — начало июня в зависимости от широты. Яйца слегка блестящие, с желтоватой, охристой, временами сероватой или зеленоватой скорлупой, на которой имеются красноватые, коричневые либо бурые пятна разной формы и интенсивности. Размеры яиц: (39—48) × (27—35) мм[64].
Насиживание продолжается 20—23 дня[65]. При приближении человека в начале насиживания самка незаметно сходит с гнезда, прячась в густой траве, позднее отводит от гнезда, опустив крылья, либо взлетая возле пришельца и всячески привлекая к себе внимание. Птенцы выводкового типа, покрыты пухом — сверху желтовато-бурым, снизу беловатым, с лёгким красновато-жёлтым налётом, более ярко выраженным в области зоба[66]. С первых дней они вполне самостоятельно добывают себе корм — мелких беспозвоночных, однако в первые дни обогреваются матерью[67]. На крыло становятся через 25—28 дней[65].
Враги и выживаемость

В гнездовой период на взрослых турухтанов и птенцов охотятся многие хищные птицы, среди которых крупные виды чаек, ворон, чёрная и серая вороны, большой и короткохвостый поморники. Из наземных животных опасность представляют песец и обыкновенная лисица. Влияние на популяцию турухтанов таких хищников, как горностаи и дикие кошки, остаётся малоизученным[68][69][70]. В местах массового скопления токующих турухтанов их смертность возрастает, поскольку токовище представляет собой удобную площадку для охоты, а возбуждённые птицы частично теряют бдительность[70]. В условиях содержания в питомниках основные причины смертности потомства связаны со стрессом и болезнью, имеющей название кривошея[67]. Эктопаразиты имеют слабое влияние на взрослых птиц[71], однако в местах зимовок турухтаны могут подхватить различные тропические болезни, такие как птичья малярия[72].
Средняя продолжительность жизни турухтана составляет 4,4 года[73]. Максимально известный возраст в Европе, 13 лет 11 месяцев, был зарегистрирован в Финляндии[74].
Природоохранный статус
_-_Philomachus_pugnax_(028b).jpg.webp)
Гнездовая область распространения турухтана оценивается в районе 8580 тыс. кв. км, общая численность 2—2,6 млн пар[75]. Европейская популяция, занимающая 50—74 % всего гнездового ареала, составляет от 200 до 510 тыс. пар. Полагают, что за 1999—2009 годы она могла сократиться до 30 %, однако это скорее связано с переменой районов гнездования, нежели чем с общим падением численности. Азиатская популяция, по оценкам экологов, остаётся стабильной. По вышеперечисленным причинам Международный союз охраны природы рассматривает данный вид как таксон наименьшего риска (категория LC)[75].
Наиболее крупные группировки отмечены на территории России и Швеции — в этих странах распространение турухтанов не вызывает опасений. В Норвегии ареал расширяется за счёт южных районов, а в Финляндии, Польше, Латвии и Нидерландах численность за последние годы сократилась более чем в два раза, что является продолжением общей тенденции в Европе за последние два столетия. Основными причинами деградации называют осушение болот, интенсивное использование минеральных удобрений, уменьшение площадей для пастбищ и заготовки сена[31].
Ископаемые находки, относящиеся к плейстоцену, указывают на то, что в межледниковый период турухтаны гнездились в более южных широтах, чем сейчас[76]. Чувствительность этого вида к общему изменению климатических условий, уровню грунтовых вод и скорости воспроизводства травянистой растительности даёт основание полагать, что изменение географии его распространения произошло под влиянием глобального потепления. Таким образом, турухтан может стать индикатором при наблюдении за изменениями климата[77]. Потенциальную опасность для птиц также могут представлять эпидемии болезней, к которым они чувствительны — птичьего гриппа, ботулизма и птичьей малярии[75].
Птица и человек
Турухтан традиционно относится к промысловой дичи, наряду с некоторыми другими куликами, такими как вальдшнеп, гаршнеп, бекас и дупель. Особенно популярной была охота на эту птицу в Средние века — например, в Англии по случаю избрания на пост архиепископа Йоркского Джорджа Невилла (англ. George Neville) в 1465 году на стол было подано 2400 тушек птиц. В те времена их отлавливали на токовище с помощью силка (сети), а также перед убоем иногда откармливали хлебом, молоком и семенами[78][79]. Позднее на куликов стали охотиться мелкой дробью из огнестрельного оружия. Вот как в середине XIX века описывает охоту на эту дичь известный русский писатель и заядлый охотник С. Т. Аксаков[16]:
Весною они появляются большими и даже огромными стаями по краям болотных луж, по берегам озёр, разлившихся рек и прудов, отдалённых от жилья. Весело, бывало, смотреть на них, проворно бегающих по начинающим зеленеть лужайкам, сверкающих на солнце яркостью своих перьев с их разноцветными золотистыми отливами. Точно широкая радуга легла на землю и бежит по ней, волнуясь и изгибаясь. В это время надобно стрелять их с подъезда. Несмотря на то, что они не дики и не сторожки, много их не убьёшь, потому что они не стоят на одном месте, а все рассыпаются врозь и бегут, не останавливаясь, как ручьи воды, по всем направлениям. Видя перед собой целые сотни этих красивых птичек, не захочешь от жадности стрелять по одному курахтану, а вместе они сбегаются редко, и поэтому часто пропускаешь удобное время.
В 1804 году русский поэт и будущий герой Отечественной войны Денис Давыдов пишет басню «Орлица, Турухтан и Тетерев», в которой под орлицей подразумевает императрицу Екатерину II, под турухтаном — зашедшего на престол после её смерти Павла I, а под тетеревом — Александра I. Басня намекает на убийство Павла I в Михайловском замке в марте 1801 года при молчаливом попустительстве Александра I, сына Павла. При этом турухтану (Павлу) даётся нелестная характеристика[80][81][82]:

А он —
Лишь шаг на трон,
То хищной тварью всей себя и окружил:
Сычей, сорок, ворон — в павлины нарядил,
И с сею сволочью он тем лишь забавлялся,
Что доброй дичью всей без милости ругался:
Кого велит до смерти заклевать,
Кого в леса дальнейшие сослать,
Кого велит терзать сорокопуту —
И всякую минуту
Несчастья каждый ждал,
Томился птичий род, стонал…
В Санкт-Петербурге известна так называемая «дорога на Турухтанные острова». Сами эти болотистые острова, существовавшие в районе Угольной гавани в нижнем течении реки Екатерингофки, были засыпаны в 1930-х годах. Известные орнитологи и писатели А. С. Мальчевский и Ю. Б. Пукинский, со ссылкой на работу «Птицы С.-Петербургской губернии» русского зоолога Е. А. Бихнера, утверждают, что острова были названы в честь турухтанов, водившихся на них во второй половине XIX века[83]. Другая версия никак не связана с птицами — острова могли быть названы в честь мануфактуры по выделке крахмала и пудры, открытой в этом районе в 1720 году. Мануфактуру на немецкий манер называли «крухмальной» или «трухманной», а специалистов — «трухманными»[84]. Тем не менее, турухтаны действительно связаны с Санкт-Петербургом. Чучела этих птиц, ныне хранящиеся на кафедре зоологии позвоночных в Санкт-Петербургском университете, были созданы из материала, добытого на островах Вольный и Голодай (ныне остров Декабристов). Ранее птицы нередко гнездились в районе Лахты и до последнего времени встречаются в сырых местах на окраине города, например, в районе железнодорожной станции Броневая[83].

Основоположник российской орнитологии М. А. Мензбир в своей книге «Охотничьи и промысловые птицы европейской России и Кавказа» (1900) приводит другие распространённые варианты названия птицы — курухтан и курахтан, а также названия, используемые в отдельных районах Российской империи — просяник, драчун (Астраханская губерния), брыжач (Херсонская губерния), таратайка (Харьковская губерния), трухтанчик (Воронежская губерния), пеунец (Ярославская губерния), петушок (Пермская губерния, частично Поволжье) и морской петушок (Сибирь)[85]. В настоящее время все эти названия почти не используются. Украинский и российский лингвист Л. А. Булаховский полагал, что именно названия курухтан или курахтан («птица, похожая своим образованием на курицу») являлись оригинальными и были заимствованы из восточных языков[86].
Примечания
- Коблик Е. А., Редькин Я. А., Архипов В. Ю. Список птиц Российской Федерации. — М.: Товарищество научных изданий КМК, 2006. — 256 с. ISBN 5-87317-263-3
- Степанян, 2003, с. 194.
- Thomas et al., 2004, pp. 1—18.
- Jukema & Piersma, 2006, pp. 161—164.
- Рябицев, 2001, с. 219.
- Hayman et al., 1986, p. 387.
- Linnaeus, 1998.
- Merrem, 1804, с. 542.
- Liddell & Scott, 1980, p. 181.
- Jobling, 1992, p. 181.
- Simpson, 1977, p. 454.
- Фасмер, 1973, с. 125.
- Mullarney et al., 2000, p. 156.
- Иванов и др., 1953, с. 96—99.
- Рябицев, 2001, с. 218.
- Аксаков, 1966, с. 230.
- Козлова, 1962, с. 21—22.
- Дементьев и Гладков, 1951, с. 176.
- Козлова, 1962, с. 22.
- Дементьев и Гладков, 1951, с. 177—178.
- Lank & Dale, 2001, pp. 759—765.
- Jukema & Piersma, 2000, pp. 289—296.
- Karlionova et al., 2008, p. 39—45.
- Pearson, 1981, p. 158—182.
- Hayman et al., 1986, p. 386.
- Britton, 1980, pp. 333—345.
- Vinicombe, 1983, p. 203—206.
- Рябицев, 1986, с. 101.
- Козлова, 1962, с. 32.
- Козлова, 1962, с. 27.
- Ruff Philomachus pugnax (PDF). United Kingdom Special Protection Areas. Joint Nature Conservation Committee. Дата обращения: 30 апреля 2010. Архивировано 20 августа 2011 года. given in Snow & Perrins (1998) 628—632
- Snow & Perrins, 1998, p. 628—632.
- Козлова, 1962, с. 29.
- Рогачёва, 1988.
- Vaillancourt et al., 1983, pp. 317—325.
- Alerstam & Christie, 2008, pp. 55—57.
- Hayman et al., 1986, pp. 386—387.
- Slater, 1970, p. 310.
- Johnson et al., 2007, pp. 277—293.
- Saryarka — steppe and lakes of northern Kazakhstan (недоступная ссылка). World heritage sites. United Nations Environment Programme and World Conservation Management Centre. Дата обращения: 30 апреля 2010. Архивировано 14 января 2009 года.
- Хроков, 1988, с. 224—225.
- Baccetti et al., 1998, pp. 365—369.
- Berthold et al., 2001, pp. 51—52.
- Сыроечковский и Рогачёва, 1980.
- del Hoyo et al., 1996, p. 531.
- Longevity, ageing, and life history of Philomachus pugnax. Human Ageing Genomic Resources. Integrative Genomics of Ageing Group, University of Liverpool. Дата обращения: 2 мая 2010. Архивировано 20 августа 2011 года.
- Philomachus pugnax (Linnaeus, 1758) — Турухтан. Позвоночные животные России. Институт Северцова. Дата обращения: 3 мая 2010.
- Roulin, 2004, pp. 1—34.
- Payne, 1984, p. 13—14.
- Мальчевский и Пукинский, 1983, с. 183—184.
- Рябицев, 1986, с. 102.
- Widemo, 1997, pp. 211—217.
- Hill, 1991, pp. 367—372.
- Hugie & Lank, 1997, pp. 218—225.
- Hoglund et al., 1998, pp. 370—376.
- Lank et al.1995, p. 59—62.
- Lank et al., 1999, p. 2323—2330.
- Verkuil et al., 2008, pp. 241—246.
- Lank, David B. Ruff Project. Simon Fraser University. Дата обращения: 11 июня 2009. Архивировано 20 августа 2011 года.
- Hooijmeijer, Jos (ed.). Newsletter Ruff Research 2009. University of Groningen. Дата обращения: 3 мая 2010. Архивировано 20 августа 2011 года.
- Lank & Smith, 1987, pp. 137—145.
- del Hoyo et al., 1996, p. 469.
- Lank et al., 2002, pp. 209—215.
- Рябицев, 2001, с. 220.
- Ruff Philomachus pugnax [Linnaeus, 1758]. Bird facts. British Trust for Ornithology. Дата обращения: 3 мая 2010. Архивировано 20 августа 2011 года.
- Дементьев и Гладков, 1951, с. 177.
- Achim, Johann. EAZA Husbandry guidelines for the Ruff (Philomachus pugnax) (недоступная ссылка). European Association of Zoos and Aquaria (EAZA). Дата обращения: 3 мая 2010. Архивировано 10 мая 2004 года.
- Management of wet grassland habitat to reduce the impact of predation on breeding waders: Phase 2. Science and research projects. Department for Environment, Food and Rural Affairs (DEFRA). Дата обращения: 4 мая 2010. Архивировано 20 августа 2011 года.
- Bolton et al., 2007, pp. 534—544.
- van der Wal & Palmer, 2008, pp. 256—258.
- Threlfall & Wheeler, 1986, pp. 273—275.
- Mendes et al., 2005, pp. 396—404.
- Lozano & Lank, 2004, pp. 315—329.
- European Longevity Records. The European Union for Bird Ringing (1 октября 2008). Дата обращения: 4 мая 2010. Архивировано 19 августа 2011 года.
- Ruff (Philomachus pugnaxies). BirdLife species factsheet. BirdLife International. Дата обращения: 3 мая 2010. Архивировано 20 августа 2011 года.
- Bocheński, 2002, pp. 239—252.
- Zockler, 2002, pp. 19—29.
- Cocker & Mabey, 2005, pp. 211—212.
- Pennant, 1776, p. 460.
- Давыдов, 2010.
- Барков, 2013.
- Давыдов, Д. В. Стихотворения. Дата обращения: 5 мая 2010.
- Мальчевский и Пукинский, 1983, с. 183.
- Турухтанный остров. Всё о реке Неве: мосты, притоки, наводнения…. Дата обращения: 5 мая 2010.
- Мензбир, 1900—1912, с. 172.
- Голев Н. Д. Функции мотивизации и народная этимология. Труды по лингвистике. Алтайский государственный университет. Дата обращения: 6 мая 2010.
Литература
- Аксаков С. Т. Записки ружейного охотника Оренбургской губернии // Собрание сочинений в 5 т. — М.: Правда, 1966. — Т. 5. — 488 с.
- Барков, А. Денис Давыдов. — НИГМА, 2013. — 32 с. — ISBN 978-5-4335-0077-8.
- Давыдов Д. В. Гусарская исповедь. Стихотворения. — М.: Директ-Медиа, 2010. — 171 с. — ISBN 9785998911521.
- Дементьев Г. П., Гладков Н. А. Птицы Советского Союза. — Советская наука, 1951. — Т. 3. — 680 с.
- Иванов А. И., Козлова Е. В., Портенко Л. А., Тугаринов А. Я. Птицы СССР. — М.: Издательство Академии наук СССР, 1953. — Т. 2. — 344 с.
- Козлова Е. В. Ржанкообразные. Подотряд Кулики Ч. 3 // Фауна СССР. Птицы. — М.—Л.: Издательство АН СССР, 1962. — Т. 2, вып. 1. — 434 с. — (Новая серия № 81).
- Мальчевский А. С., Пукинский Ю. Б. Птицы Ленинградской области и сопредельных территорий. — Л.: Издательство Ленинградского университета, 1983.
- Мензбир М. А. Охотничьи и промысловые птицы европейской России и Кавказа. — Типо-литография т-ва И. Н. Кушнарёв и Ко, 1900—1912. — Т. 1. — 343 с.
- Рогачёва Э. В. Птицы Средней Сибири. Распространение, численность, зоогеография. — М.: Наука, 1988. — 310 с. — ISBN 5-02-005252-3.
- Рябицев В. К. Птицы тундры. — Свердловск: Средне-Уральское книжное издательство, 1986. — 192 с.
- Рябицев В. К. Птицы Урала, Приуралья и Западной Сибири: Справочник-определитель. — Екатеринбург: Изд-во Уральского университета, 2001. — 608 с. — ISBN 5-7525-0825-8.
- Степанян Л. С. Конспект орнитологической фауны России и сопредельных территорий. — М.: Академкнига, 2003. — 727 с.
- Сыроечковский Е. Е., Рогачёва Э. В. Животный мир Красноярского края. — Красноярск: Красноярское книжное издательство, 1980. — 360 с.
- Фасмер М. Этимологический словарь русского языка: Пер. с нем. (с доп.). — М.: Прогресс, 1973. — Т. 4. — 855 с.
- Хроков В. В. Низовья р. Тургай — новое место гнездования турухтана // Орнитология. — 1988. — Т. 23. — С. 224—225.
- Alerstam, Thomas; Christie, David A. Bird Migration. — Cambridge University Press, 2008. — 432 p. — ISBN 0521448220.
- Baccetti, N.; Gambogi, R.; Magnani, A.; Piacentini, D.; Serra, L. Stop-over strategy of Ruff Philomachus pugnax during the spring migration // International Wader Studies. — 1998. — Т. 10. — P. 365—369. Архивировано 7 июня 2011 года. (англ.)
- Berthold, Peter; Bauer, Hans-Günther; Westhead, Valerie. Bird migration: a general survey (англ.). — Oxford: Oxford University Press, 2001. — 272 p. — ISBN 0198507879. (англ.)
- Bolton, Mark; Tyler, Glen; Smith, Ken; Bamford, Roy. The impact of predator control on lapwing Vanellus vanellus breeding success on wet grassland nature reserves // Journal of Applied Ecology. — 2007. — Т. 44. — P. 534—544. — doi:10.1111/j.1365-2664.2007.01288.x. (англ.)
- Bocheński, Zygmunt. Bird remains from Oblazowa — zoogeographical and evolutionary remarks // Acta Zoologica Cracoviensia. — 2002. — Т. 45. — P. 239—252. (англ.)
- Britton, David. Identification of Sharp-tailed Sandpipers // British Birds. — 1980. — Т. 73, № 8. — P. 333—345. (англ.)
- Cocker, Mark; Mabey, Richard. Birds Britannica. — Chatto & Windus, 2005. — 484 p. — ISBN 0701169079.
- Jobling, James А. A Dictionary of Scientific Bird Names (англ.). — United States: Oxford University Press, 1992. — ISBN 0198546343.
- Johnson, James A.; Lanctot, Richard B.; Andres, Brad A.; Bart, Jonathan R.; Brown, Stephen C.; Kendall, Steven J.; Payer, David C. Distribution of Breeding Shorebirds on the Arctic Coastal Plain of Alaska // Arctic. — 2007. — Т. 60, № 3. — P. 277—293. (англ.)
- Jukema, Joop; Piersma, Theunis. Contour feather moult of Ruffs Philomachus pugnax during northward migration, with notes on homology of nuptial plumages in scolopacid waders (англ.) // Ibis. — Wiley-Blackwell, 2000. — Vol. 142, no. 2. — P. 289—296. doi:10.1111/j.1474-919X.2000.tb04868.x (англ.)
- Jukema, Joop; Piersma, Theunis. Permanent female mimics in a lekking shorebird (англ.) // Biology Letters. — Royal Society Publishing, 2006. — Vol. 2, no. 2. — P. 161—164. — doi:10.1098/rsbl.2005.0416. Архивировано 24 июля 2011 года. (англ.)
- Hayman, Peter; Marchant, John; Prater, Tony. Shorebirds: an identification guide to the waders of the world. — Boston: Houghton Mifflin, 1986. — 412 p. — ISBN 0-395-60237-8. (англ.)
- Hill, Wendy L. Correlates of male mating success in the ruff Philomachus pugnax, a lekking shorebird // Behavioral Ecology and Sociobiology. — 1991. — Т. 29, № 5. — P. 367—372. doi:10.1007/BF00165962 (англ.)
- Hoglund, Jacob; Widemo, Fredrik; Sutherland, William J.; Nordenfors, Helena. Ruffs, Philomachus pugnax, and distribution models: can leks be regarded as patches? // Oikos. — 1998. — Т. 82, № 2. — P. 370—376. (англ.)
- del Hoyo, Josep; Elliott, Andrew; Sargatal, Jordi. Hoatzin to Auks. Volume 3 // Handbook of the Birds of the World. — Barcelona: Lynx Edicions, 1996. — 821 p. — ISBN 8487334202. (англ.)
- Hugie, Don M.; Lank, David B. The resident's dilemma: a female choice model for the evolution of alternative mating strategies in lekking male ruffs (Philomachus pugnax) // Behavioral Ecology. — 1997. — Т. 8, № 2. — P. 218—225. doi:10.1093/beheco/8.2.218 (англ.)
- Karlionova, Natalia; Meissner, Wlodzimierz; Pinchuk, Pavel. Differential development of breeding plumage in adult and second-year male Ruffs Philomachus pugnax // Ardea. — 2008. — Т. 96, № 1. — P. 39—45. (англ.)
- Lank, D. B.; Coupe, M.; Wynne-Edwards, K. E. Testosterone-induced male traits in female ruffs (Philomachus pugnax): autosomal inheritance and gender differentiation // Proceedings of the Royal Society of London: Biology. — 1999. — Т. 22. — P. 2323—2330. doi:10.1098/rspb.1999.0926 (англ.)
- Lank, David B.; Dale, James. Visual signals for individual identification: the silent «song» of Ruffs (англ.) // American Ornithological Society. — 2001. — Vol. 118, no. 3. — P. 759—765. doi:10.1642/0004-8038(2001)118[0759:VSFIIT]2.0.CO;2 (англ.)
- Lank, David B.; Smith, Constance M. Conditional lekking in ruff (Philomachus pugnax) // Behavioral Ecology and Sociobiology. — 1987. — Т. 20, № 2. — P. 137—145. doi:10.1007/BF00572636 (англ.)
- Lank, David B.; Smith, Constance M.; Hanotte, Olivier; Burke, Terry; Cooke, Fred. Genetic polymorphism for alternative mating behaviour in lekking male ruff Philomachus pugnax // Behavioral Ecology. — 1995. — Т. Nature. — P. 59—62. doi:10.1038/378059a0 (англ.)
- Lank, David B.; Smith, Constance M.; Hanotte,Olivier; Ohtonen, Arvo; Bailey, Simon; Burke, Terry. High frequency of polyandry in a lek mating system // Behavioral Ecology. — 2002. — Т. 13, № 2. — P. 209—215. doi:10.1093/beheco/13.2.209 (англ.)
- Liddell, Henry George; Scott, Robert. A Greek-English Lexicon (Abridged edition) (англ.). — Oxford: Oxford University Press, 1980. — 808 p. — ISBN 0199102074.
- Linnaeus, C. Editio decima, reformata // Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. — Holmiae: Laurentii Salvii, 1998. (лат.)
- Lozano, G. A.; Lank, D. B. Immunocompetence and testosterone-induced condition traits in male ruffs (Philomachus pugnax) // Animal Biology. — 2004. — Т. 54, № 4. — P. 315—329. Архивировано 12 июля 2013 года. doi:10.1163/1570756042729555 (англ.)
- Mendes, Luisa; Piersma, Theunis; Lecoq, M.; Spaans, B.; Ricklefs, Robert E. Disease-limited distributions? Contrasts in the prevalence of avian malaria in shorebird species using marine and freshwater habitats // Oiko. — 2005. — Т. 109. — P. 396—404. doi:10.1111/j.0030-1299.2005.13509.x (англ.)
- Merrem, Blasius. Philomachus (нем.) // Allgemeine literatur-zeitung. — 1804. — Nr. 168. — S. 542.
- Mullarney, Killian; Svensson, Lars; Zetterström, Dan; Grant, Peter J. Birds of Europe. — United States: Princeton University Press, 2000. — 400 p. — ISBN 978-0-691-05054-6. (англ.)
- Payne, Robert B. Sexual Selection, Lek and Arena Behavior, and Sexual Size Dimorphism in Birds. — American Ornithologists Union, 1984. — 52 p. — ISBN 0943610400. (англ.)
- Pearson, D. J. The wintering and moult of ruffs Philomachus pugnax in the Kenyan Rift Valley (англ.) // Ibis. — Wiley-Blackwell, 1981. — Vol. 123, no. 2. — P. 158—182. doi:10.1111/j.1474-919X.1981.tb00922.x (англ.)
- Pennant, Thomas. British Zoology. — London: B. White, 1776. — Т. Volume 2: Water Birds. (англ.)
- Roulin, Alexandre. The evolution, maintenance and adaptive function of genetic colour polymorphism in birds // Biological Reviews. — 2004. — Т. 79. — P. 1—34. (недоступная ссылка) doi:10.1017/S1464793104006487 (англ.)
- Simpson, D. P. Cassell's Latin Dictionary. — London: Webster's New World, 1977. — 912 p. — ISBN 978-0025225800. (англ.)
- Slater, Peter. A field guide to Australian birds: non-passerines. — Adelaide: Rigby, 1970. — ISBN 0851791026. (англ.)
- Snow, David; Perrins, Christopher M. The Birds of the Western Palearctic concise edition (2 volumes) (англ.). — Oxford: Oxford University Press, 1998. — 1830 p. — ISBN 019854099X. (англ.)
- Thomas, Gavin H.; Wills, Matthew A.; Székely, Tamás. A supertree approach to shorebird phylogeny (англ.) // BioMed Central. — 2004. — Vol. 4, no. 28. — P. 1—18. doi:10.1186/1471-2148-4-28 (англ.)
- Threlfall, William; Wheeler, Terry A. Ectoparasites from birds in Newfoundland // Journal of Wildlife Disposes. — 1986. — Т. 22, № 2. — P. 273—275. (англ.)
- Vaillancourt, Eric; Prud'Homme, Sophie; Haman, François; Guglielmo, Christopher G; Weber, Jean-Michel. Energetics of a long-distance migrant shorebird (Philomachus pugnax) during cold exposure and running (англ.) // The Journal of Experimental Biology. — The Company of Biologists, 1983. — Vol. 208, no. Pt 2. — P. 317—325. doi:10.1242/jeb.01397 (англ.)
- Verkuil, Yvonne I.; Jukema, Joop; Gill, Jennifer A.; Karlionova, Natalia; Melter, Johannes; Hooijmeijer, Jos C. E. W; Piersma, Theunis. Non-breeding faeder Ruffs Philomachus pugnax associate according to sex, not morphology: capsule faeders (males that are female look-alikes) associate with males rather than females, at several different spatial scales // Bird Study. — 2008. — Т. 55, № 3. — P. 241—246. doi:10.1080/00063650809461529 (англ.)
- Vinicombe, Keith. Identification pitfalls and assessment problems: 4. Buff-breasted Sandpiper Tryngites subruficollis // British Birds. — 1983. — Т. 76, № 5. — P. 203—206. (англ.)
- van der Wal, R.; Palmer, Stephen C. Is breeding of farmland wading birds depressed by a combination of predator abundance and grazing? // The Royal Society of London: Biology Letters. — 2008. — Т. 8, № 2. — P. 256—258. doi:10.1098/rsbl.2008.0012 (англ.)
- Widemo, Fredrik. The social implications of traditional use of lek sites in the ruff Philomachus pugnax // Behavioral Ecology. — 1997. — Т. 8, № 2. — P. 211—217. doi:10.1093/beheco/8.2.211 (англ.)
- Zockler, Christoph. Declining Ruff Philomachus pugnax populations: a response to global warming? // Wader Study Group Bulletin. — 2002. — Т. 97. — P. 19—29. (англ.)
Ссылки
- Описание и распространение турухтана. Позвоночные животные России. Институт РАН им. А. Н. Северцова. Дата обращения: 6 мая 2010.
- Фотографии, звуковые файлы и видеоклипы с участием турухтана. Internet Bird Collection. Lynx. Дата обращения: 6 мая 2010. Архивировано 20 августа 2011 года.
| Таксономия | |
|---|---|
| В библиографических каталогах |