Скелет анкилозавров
Скелет акилозавров, как и у всех позвоночных животных, являлся частью опорно-двигательной системы, и был устроен по общему принципу четвероногих динозавров. Тем не менее анкилозавры представляли собой достаточно своеобразную группу, характеризующуюся обширными продольными рядами дермальных окостенений на верней и боковой поверхностях шеи, туловища и хвоста, в результате чего в скелете наблюдается приспособление к ношению и поддержке остеодермального панциря. В дополнение к этому анкилозавры обладают необычной черепной архитектурой, которая включает костный «орнамент» (результат окостенения кожи)[1].
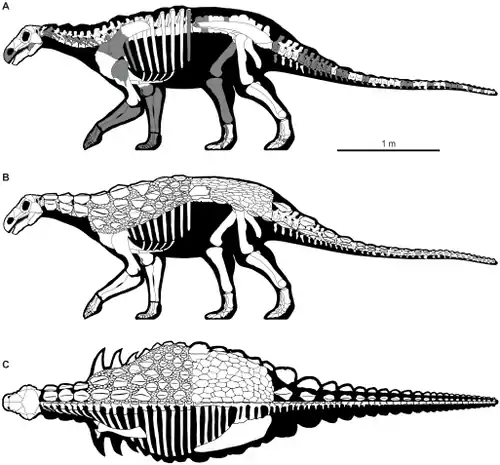
Морфология анкилозавров указывает на их сильную специализированность и большую примитивность по сравнению с остальными динозаврами[2]. По степени выраженности некоторых морфологических особенностей анкилозавры делятся на два семейства: более примитивных нодозаврид и более специализированных анкилозаврид[3]. Основные различия заключаются в форме и пропорциях черепа, характере остеодермального перекрытия крыши черепа, величине зубов, строении нёба, мозговой коробки, затылочной области, в строении ряда элементов посткраниального скелета, а также в наличии или отсутствии костной булавы на конце хвоста[4].
Детальное изучение морфологии анкилозавров, их сопоставление с другими динозаврами и рептилиями дают возможность расширить знания об их систематическом положении, родственных связях, происхождении, о взаимоотношениях североамериканских и азиатских представителей, позволяет ставить вопросы по эволюции динозавров в целом[2].
Череп
Общее описание

Черепа анкилозавров отличаются сильным окостенением и массивностью. Длина черепа взрослых особей варьируется от 20 до 60 см. Ширина в области глазниц может превышать длину[5][6][7]. Самый большой из когда-либо найденных черепов анкилозавров в ширину достигает 66,2 см (образец CMN 8880 рода анкилозавр)[8], череп рода Cedarpelta имеет длину, равную примерно 60 см[9]. Архитектура черепа указывает на его акинетичность, то есть верхняя челюсть не перемещаться вертикально относительно черепной коробки[1].
Общая синапоморфия черепов анкилозавров заключается в наличии хорошо развитого черепного «орнамента»[6][1], а также в закрытии двух черепных окон — предглазничного и верхнего височного[5][1]. Вместо двух височных окон, типичных для динозавров, у анкилозавров — одно, приближающееся по положению к нижнему височному окну. Оно расположено между заглазничными, чешуйчатыми, квадратными и квадратно-скуловыми костями, однако у представителей семейства анкилозаврид они закрыты боковыми разросшимися остеодермами[2]. Верхние височные окна ни у одного из известных анкилозавров не обнаружены[2][10]. Существовало предположение об их зарастании остеодермами[11], однако их нет и на молодом экземпляре пинакозавра[12][2]. Было предложено по крайней мере два сценария для объяснения закрытия черепных отверстий у анкилозавров. Ряд авторов предположили, что закрытие является результатом расширения окружающих костей черепа[13][14]. Другие исследователи выдвинули гипотезу, что эта особенность является следствием общего утолщения черепных костей и покрытия отверстий остеодермами[12][15][16][17]. Наличие незакрытого предглазничного окна (англ. antorbital fenestra) в виде небольшой ямки у некоторых представителей анкилозавров свидетельствует о том, что череп принадлежит молодой особи[18].
Глазницы анкилозавров средних размеров, расположенные по бокам черепа, направленные у большинства несколько вперёд, а у шамозавра — вбок[2]. Их форма у разных видов разница[19] и вероятно является результатом деформации при захоронении[20]. Мозговая коробка панцирных динозавров почти прямая, низкая[2][10], что обусловлено направленностью основной затылочной кости, тогда как мозговая коробка других динозавров представляет собой высокую конструкцию[2].
Форма черепа
При виде сверху черепа большинства анкилозавров представляют собой некое подобие треугольника или трапеции (у семейства нодозаврид передняя часть морды уже, чем у семейства анкилозаврид)[2], либо, в редких случаях, имеют субпрямоугольный контур (например Ziapelta)[21].
У примитивных анкилозаврид череп больше в длину, чем ширину, в отличие от более широких черепов производных анкилозаврид[22]. Самая широкая часть крыши черепа образована верхними заглазничными шипами[19]. При виде сбоку, верх черепа от уровня глазниц к кончику морды имеют дугообразный (например у Gastonia, Euoplocephalus и Ankylosaurus) или почти плоских вид[23]. Предполагается, что эти различия являются результатом деформации в процессе захоронения[18]. Сзади от глазниц теменная область варьирует от плоской до вогнутой.
Ростральная и теменная области
Сильным сдерживающим фактором в изучении морфологии черепа анкилозавров является сплошное перекрытие крыши черепа остеодермальными щитками[2], из-за чего черепные швы как правило не обнаруживаются у взрослых анкилозавров[10][24][1]. Шовные контакты черепа известны только по нескольким таксонам: Pinacosaurus grangeri, Liaoningosaurus, Kunbarrasaurus и Cedarpelta[3][17].
Парные теменные (англ. parietal), лобные (англ. frontal) и носовые (англ. nasal) кости формируют крышу черепа. Эти кости полностью закрыты у взрослых особей сращиванием щитков по всей их верхней поверхности. Носовые кости дополнительно закрыты и на боковой поверхности, в нижней части они скрыты вторичным нёбом верхней челюсти[25]. Носовые кости большие, обеспечивают переднюю часть крыши черепа и часть боковой стенки носовой полости[26][17], спереди они граничат с предчелюстными костями, сзади соприкасаются с лобными и предлобными костями[27]. Теменные кости покрывают заднюю часть черепной коробки, расположены над верхней затылочной костью и контактируют с последней через выступающий вниз шишковидный отросток[28].
Предчелюстные кости (англ. premaxilla) состоят из основной кости, а на переднем кончике морды заканчивается клювом[3]c.368. Они разделяют наружные ноздри на две части, поднимаясь отростком высоко между ноздрями к носовым костям и внедряясь в них[29][3]c.369. Задним краем контактируют с верхнечелюстными костями[26]. У большинства анкилозаврид при виде сбоку видна только самая нижняя часть верхней челюсти, остальная часть закрыта боковыми щитками[25].
Предлобная кость (англ. prefrontal) расположена между носовой, лобной, слёзной и надглазничными костями[27]. Эта кость анкилозавров характерна тем, что у некоторых видов она распространяется на боковую поверхность черепа (Pinacosaurus)[30], либо вовсе контактирует с нижнечелюстной костью (Jinyunpelta)[18]. Перед глазницей располагается слёзная кость (англ. lacrimal). Лишь несколько анкилозавров, таких как молодые Pinacosaurus grangeri и Kunbarrasaurus ieversi демонстрируют чётко идентифицируемые слёзные кости[25]. У разных видов она значительно отличается, от тонкой в форме клина у Minmi до субпрямоугольной у Pinacosaurus и Cedarpelta. В нижней части слёзная кость контактирует с верхней челюстью[27]. Вдоль переднего центрального края глазницы слёзная кость может быть перфорирована носослёзным отверстием (англ. nasolacrimal aperture[30]; также слёзное отверстие[15] или слёзный проток[31]), через которое происходит удалением накопившихся излишков постоянно выделяющейся слёзной жидкости.
Все анкилозавры с видимыми черепными швами (например Pinacosaurus) обладают тремя надглазничными (англ. supraorbital) элементами — передняя надглазничная кость, средняя надглазничная кость и задняя надглазничная кость[17][28]. Надглазничные кости расположены как комплекс или серия элементов, охватывающих верхний край каждой орбиты. В дополнение к их расположению по окружности орбиты надглазничные кости могут характеризоваться частым появлением декоративных образований, напоминающих клиновидные выступы (например у Saichania) или округлые выступы (у нодозаврид)[30]. У некоторых видов выступы могут сливаться в единый надглазничный гребень (Akainacephalus, Jinyunpelta)[25][18]. Для большинства таксонов анкилозавров идентификация надглазничных костей ограничивается их декоративным украшением (то есть небольшим выступом) и следовательно их количество, и точная морфология обычно неизвестны[28].
Заглазничная кость (англ. postorbital) формирует самый задний край орбиты и контактирует спереди с надглазничными костями, снизу со скуловой и квадратично-скуловой костями, сзади с чешуйчатой костью. С внутренней стороны кость формирует поперечную стенку, образующая заднюю границу глазниц[25][28]. Скуловая кость (англ. jugal) образует нижний край глазницы и дно орбитальной полости, контактирует с задним краем верхней челюсти и передними частями квадратично-скуловой и заглазничной костей. В нижней части скуловая кость часто включает в себя продолжение задних тупых краёв клюва. Она может также образовывать узкий контакт с эктоптеригоидом, позади зубных рядов[28].
Чешуйчатая кость (squamosal), вместе с расположенной ниже квадратично-скуловой костью (quadratojugal), образуют заднебоковые части черепа. У большинства таксонов чешуйчатая кость вносит свой вклад в характерные черепные украшения, как правило верхний заглазничный шип (у анкилозаврид) или шишковидный выступ (например у Edmontonia)[32]. Квадратично-скуловая кость практически у всех таксонов анкилозавров развиваются в виде выростов, морфология которых варьируется от округлых бугорков (у нодозаврид) до полноценных выступов в виде нижнего заглазничного шипа (у анкилозаврид)[33]. Роговые выросты являются отличительными особенностями семейства анкилозаврид[2]. Верхних шип обычно имеет треугольную форму, его длина и острота варьируются, как и угол, под которым он выступает из черепа. Нижний шип также значительно различается по размеру и углу выступа из черепа, его верхушка может быть острой или круглой. Таксономическое значение формы верхнего заглазничного шипа у анкилозаврид является спорным[34].
Основание черепа и затылочная область
.jpg.webp)
Основание черепа, или базикраний (лат. basicranium), состоит из базисфеноида, образующий переднюю часть основания черепа и основной затылочной кости, образующую заднюю часть основания черепа[21]. Контакт между базисфеноидом и основной затылочной костью разграничивается базальным бугром затылка (англ. basal tubera)[35].
Базисфеноид (англ. basisphenoid), или основная клиновидная кость, формируя основание среднего мозга, спереди примыкает к парасфеноиду (англ. parasphenoid), который в виде отростка поддерживал мембрану, разделявшую правое и левое глазные яблоки[35]. Верхняя часть контактирует с парными элементами латеросфеноида и передней ушной кости. По его бокам расположены птеригоиды (англ. pterygoid), или крыловидные кости, которые имеет достаточно сложную морфологию. Они делятся на три основных компонента: центральное тело, контактирующее с сошниками и базисфеноидом, птеригоидные фланги (англ. pterygoid flange) и квадратные отростки птеригоидов (англ. quadrate ramus)[33]. Птеригоидные фланги представляют собой вертикальную костную структуру, перпендикулярную сошнику, расположены в виде поперечных перегородок и образуют заднюю стенку нёбных пустот. Фланги птеригоидов у анкилозаврид направлены вбок, тогда как у нодозаврид они тянутся вперёд параллельно сошникам[36][33]. Птеригоидные фланги контактируют с эктоптеригоидами около границ окончания зубного ряда верхней челюсти. Эктоптеригоид (англ. ectopterygoid), или наружная крыловидная кость, представляет собой небольшой клиновидный элемент, образующий задний край вторичного нёба, а также передний край подглазничного окна (англ. suborbital fenestra)[37]. Эпиптеригоид (англ. epipterygoid) представляет собой небольшую тонкую, цилиндрическую структуру, отходящий от передней половины крыловидных отростков и соединяющийся с крышкой черепа. Отделяет крыловидную кость от верхней челюсти, и встречается не у всех видов[19][38]. Отходящий квадратный отросток птеригоида сочленяется с квадратной костью[19]. У некоторых представителей возможно срастание птеригоидов с базиптеригоидными отростками (англ. basipterygoid process), тогда как у всех динозавров обычно эти кости разделены пластиной чешуйчатой кости[2]. Эти короткие, тупые отростки, отклоняются от передних границ базисфеноида, охватывая задний край парасфеноида с обеих сторон[35].
Внутри глазниц анкилозавров находятся три базикраниальных элемента (межглазничные окостенения), которые вносят вклад в центральную стенку, разделяющую глазницы. Практически во всех случаях невозможно чётко разграничить орбитосфеноид (англ. orbitosphenoid), пресфеноид (англ. presphenoid) и спленэтмоид (англ. sphenethmoid)[35]. Самым передним из межглазничных окостенений является спленэтмоид[2].
Затылочный отдел (англ. occipital region) черепа состоит из основной затылочной кости, парных боковых затылочных костей и верхней затылочной кости, которые окружают большое затылочное отверстие. Основная затылочная кость (англ. basioccipital) представляет собой непарный элемент, образующая основание заднего мозга, и составляет нижний край большого затылочного отверстия. Она формирует большую часть затылочного мыщелока (occipital condyle), служащего для сочленения головы с первым шейным позвонком. У нодозаврид и анкилозаврин затылочный мыщелок состоит только из основной затылочной кости. Верхняя часть основной затылочной кости контактирует с парными боковыми затылочными и задними ушными костями. Верхняя затылочная кость (англ. supraoccipital) — это трапециевидный элемент, который срастается с нижней поверхностью теменных костей и образующий верхний край большого затылочного отверстия (лат. foramen magnum). Отверстие служит проходом для спинного мозга. При взгляде с затылочной стороны верхняя затылочная кость контактирует переднебоковыми сторонами с боковой затылочной костью и задней ушной костью, сверху она контактирует с теменной костью. Наличие верхней ушной кости (англ. epiotic), присутствующей в других группах динозавров, у анкилозавров неизвестно[37]. Область над большим затылочным отверстием в виде гребня (англ. nuchal crests) служила поверхностью прикрепления мышц musculus trapezius, которые способствовали поддержке черепа[39]. У большинства анкилозавров заметное развитие затылочного гребня позволяет предположить, что это были основные мышцы шеи[19]. Боковые затылочные кости (англ. exoccipital), сливаясь с впереди лежащими задними ушными костями, образуют самую последнюю (заднюю) структуру затылочной области (англ. otoccipital) и способствует расходящимся горизонтальным парокципитальным отросткам (англ. paroccipital process). У анкилозавров эти отростки обычно не обширны и не расположены на всю заднюю ширину черепа. Эти затылочные отростки иногда бывают отделены от соседних костей (например у Tarchia), тогда как у других анкилозавров они часто сливаются с чешуйчатой и квадратной костями[39][37].
Парные костные элементы слуховой капсулы — передняя ушная кость (англ. prootic) и задняя ушная кость (англ. opisthotic), в которых находятся вестибулярный аппарат — образуют верхне-боковые стенки базисфеноида и основной затылочной кости соответственно[37], и вместе с латеросфеноидом формируют боковую часть эндокраниальной полости, содержащей головной мозг[25]. Латеросфеноид (англ. laterosphenoid) образует боковое окружение переднего мозга, у некоторых видов может дополнительно формировать боковые стенки задней части эндокраниальной полости. Он также вносит вклад в поперечно ориентированные заглазничные стенки. Спереди латеросфеноид контактирует с парасфеноидом, верхним краем с крышей черепа (лобно-теменным комплексом), задним краем либо с передней ушной костью, либо может расширяется в заднебоковом направлении и сливаться с боковыми затылочными костями. Передняя ушная кость спереди контактирует с латеросфеноидом, верхне-задним краем с верхней затылочной костью, задним краем с задней ушной костью, которая окончательно сливается с позади лежащими боковыми затылочными костями[35][25]. Квадратные кости (англ. quadrate) нижним концом сочленяются с нижней челюстью, образуя челюстной сустав. Верхний конец квадратных костей сочленяется с суставной поверхностью чешуйчатой кости и концами парокципитальных отростков[35]. Это слияние является очень характерным для семейства нодозаврид[10]. В средней части от квадратной кости отходит крыловидный отросток (англ. pterygoid process), который соединяется с птеригоидом[35].
Эндокраниальная полость черепа имеет черепные отверстия (англ. cranial foramina). Эти отверстия образуют пространство для размещения различных черепных нервов и для выхода внутренней яремной вены[25][37].
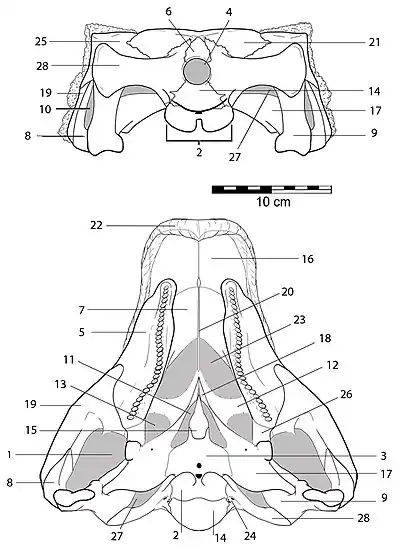
- 1 - аддукторная камера (adductor chamber)
- 2 - базальный бугор затылка (basal tubera)
- 3 - базисфеноид (basisphenoid)
- 4 - большое затылочное отверстие (foramen magnum)
- 5 - верхнечелюстная кость (maxilla)
- 6 - верхняя затылочная кость (supraoccipital)
- 7 - вторичное нёбо (secondary palate)
- 8 - квадратично-скуловая кость (quadratojugal)
- 9 - квадратная кость (quadrate)
- 10 - квадратное отверстие (quadrate foramen)
- 11 - межптеригоидная пустота (interpterygoid vacuity)
- 12 - нёбная кость (palatine)
- 13 - нёбное отверстие (palatal fenestra)
- 14 - основная затылочная кость (basioccipital)
- 15 - парасфеноид (parasphenoid)
- 16 - предчелюстная кость (premaxilla)
- 17 - птеригоид (pterygoid)
- 18 - птеригоидный отросток (rostral process of the pterygoid)
- 19 - скуловая кость (jugal)
- 20 - сошник (vomer)
- 21 - теменная кость (parietal)
- 22 - томий (tomium)
- 23 - хоаны (choanae)
- 24 - черепное отверстие подъязычного нерва (cranial foramina)
- 25 - чешуйчатая кость (squamosal)
- 26 - эктоптеригоид (ectopterygoid)
- 27 - cranio-quadrate canal
- 28 - otoccipital
Остеодермальные щитки черепа
Верхняя и боковая части черепа анкилозавров были покрыты остеодермальными щитками (костным «орнаментом»)[1]. У разных авторов они именуются по-разному: щитки, сегменты, остеодермы. В этой статье они обозначены термином «щиток» по работам Татьяны Тумановой[2]. Черепной «орнамент» анкилозавров может возникать либо в результате присоединения остеодерм к лежащим под ними костям черепа, либо в результате ремоделирования самих элементов черепа (например у Minotaurasaurus), или в результате комбинации обоих процессов[34][12][15][31][24].
У. Т. Блоу в 2001 году предложил термин caputegulum (в переводе с латинского «черепная плитка»; множественное число caputegulae) для обозначения плоских костей, покрывающих черепа анкилозавров. Этот термин обычно используется с дополнительными модификаторами, указывающими их местоположение (например prefrontal caputegulum, supraorbital caputegulae). Пирамидальные шипы чешуйчатых и квадратично-скуловых костей черепа у разных авторов называются по-разному: щитки, выступы, рога и венцы. В этой статье для упрощения применяется термин «шипы» по работам Татьяны Тумановой[2] и используются для обозначения чешуйчатого или квадратично-скулового «орнамента»[34].
Черепной «орнамент» полезен для выявления различий и сходств среди таксонов анкилозавров. Многие анкилозавры имеют плоский черепной «орнамент», разделённый неглубокими бороздами (например Ankylosaurus , Edmontonia)[34], а у некоторых анкилозавров эти отдельные участки имеют выпуклую или пирамидальную форму (например Akainacephalus johnsoni, Saichania chulsanensis, Tarchia teresae, Minotaurasaurus)[34][25]. Среди семейств у нодозаврид остеодермы достаточно крупные, симметрично расположенные, чётко разграниченные друг от друга глубокими бороздами. У анкилозаврид щитки (если они различимы и имеют определённые очертания) более мелкие, расположенные мозаично[2]. Предчелюстные кости могут быть полностью перекрыты остеодермами, в большинстве же случаев самый нижний край кости остаётся свободным от них. Позади глазниц на поверхности черепа остеодермы сливаются, образуя в этой области единый рельеф. Задний щиток (nuchal caputegulae) крышки черепа может нависать над затылком (например у Talarurus и в некоторой степени у Euoplocephalus и Shamosaurus) и закрывать его при виде сверху, а может заканчиваться на уровне затылка. Боковая часть черепа перед глазницами покрыта остеодермами несколько более крупных размеров[2]. Над глазницей у некоторых видов анкилозавров тянутся два продольных низких килеватых щитка, они составляют часть околорбитального комплекса (circumorbital) — закостеневших декоративных элементов, которые окружают глазницу[25][34]. Комплекс включает надглазничные щитки, небольшой слёзный щиток, полукруглую остеодерму скуловой кости и небольшие пластинчатые щитки по заднему краю глазницы. За глазницами, между верхним и нижним заглазничными шипами, расположены небольшие постокулярные остеодермы[25]. По крайней мере у нескольких родов — Euoplocephalus, Pawpawsaurus, Zuul[20], обнаружены костные пластинки в области век (ocular osteoderm или ciliary osteoderm)[40][10]. Хотя предназначение этого окостенения остаётся неясным, несколько таксонов современных крокодилов (Alligator, Paleosuchus) имеют аналогично расположенный, слабо выражающийся элемент[28].
Виды и места расположения щитков на черепе
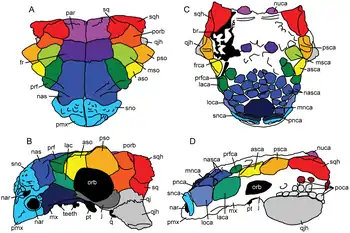
asca — передний надглазничный щиток (anterior supraorbital caputegulum); aso — передняя надглазничная кость (anterior supraorbital); br — изломы черепа/наложенный гипс; fr — лобная кость (frontal); frca — лобный щиток (frontal caputegulum); j — скуловая кость (jugal); lac — слёзная кость (lacrimal); laca — слёзный щиток (lacrimal caputegulum); loca — лореальный щиток (loreal caputegulum); mnca — средний носовой щиток (median nasal caputegulum); msca — средний надглазничный щиток (middle supraorbital caputegulum); mso — средняя надглазничная кость (middle supraorbital); mx — верхнечелюстная кость (maxilla); nar — наружные ноздри (naris); nas — носовая кость (nasal); nasca — носовой щиток (nasal caputegulum); nuca — затылочный щиток (nuchal caputegulum); orb — глазница или орбита (orbit); par — теменная кость (parietal); pmx — предчелюстная кость (premaxilla); pnca — постнариальный щиток (postnarial caputegulum); poca — постокулярные щитки (postocular caputegulum); porb — заглазничная кость (postorbital); prf — предлобная кость (prefrontal); prfca — предлобный щиток (prefrontal caputegulum); psca — задний надглазничный щиток (posterior supraorbital caputegulum); pso — задняя надглазничная кость (posterior supraorbital); pt — крыловидная кость (pterygoid); q — квадратная кость (quadrate); qj — квадратично-скуловая кость (quadratojugal); qjh — нижний заглазничный шип (quadratojugal horn); snca — супранариальный щиток (supranarial caputegulum); sno — супранариальный орнамент (supranarial ornamentation); sq — чешуйчатая кость (squamosal); sqh — верхний заглазничный шип (squamosal horn).
Наружные ноздри
Костные ноздри анкилозавров как правило крупные[2]. Они отделены друг от друга межносовым отростком — отходящим вверх продолжением предчелюстной кости[3]c.369. Наружные ноздри располагаются на кончике морды. У некоторых видов они бывают сдвинуты немного назад и ориентированы в бок, например у Gargoyleosaurus, нодозаврид и некоторых анкилозаврид (Nodocephalosaurus и Ankylosaurus), и полностью скрыты при виде спереди[25]. Для других видов характерна передняя или переднебоковая ориентацию внешние ноздрей, например у Pinacosaurus, Minotaurasaurus, Tsagantegia, Euoplocephalus и Anodontosaurus[2][25].
У большинства анкилозавров в носовой области одна пара отверстий — собственно наружные ноздри. У молодого экземпляра Pinacosaurus и Minotaurasaurus отмечается в этой области три пары отверстий[2][25]. Верхние и боковые края наружных ноздрей окаймлены супранариальным щитком, что является общим для многих нодозаврид[10], у некоторых видов нижние края также бывают ограничены щитком, сросшимся с предчелюстной костью. У рода Ankylosaurus ноздри зарастают остеодермами так, что остаются лишь небольшие боковые отверстия. От развития остеодерм в области наружных костных ноздрей зависит форма последних, размеры, положение и даже количество наружных отверстий[2].
Внутренние ноздри
Как и у других позвоночных, у анкилозавров можно выделить три основных части носовой полости: преддверие, собственно носовую полость и обонятельную область. Преддверие в значительной степени ограничено предчелюстной и носовыми костями[17]. У некоторых родов (Tsagantegia и Pinacosaurus) задний край межносового отростка сливается с костной носовой перегородкой. Кроме того, каждое преддверие может быть разделено одним или несколькими дополнительными отростками предчелюстной кости — интраназальные отростки (англ. intranasal processes). Разделение наружного носового канала интраназальными отростками предчелюстной кости приводит к образованию одного или нескольких смежных отверстий — околоносовых отверстий (англ. paranasal apertures) — они приводят к полостям придаточных пазух носа (англ. paranasal sinus), которые по-разному закрываются внутри предчелюстной кости и верхней челюсти[3]c.369. Например Edmontonia имеет двусторонние придаточные пазухи внутри предчелюстной кости и поверх верхнечелюстных рядов зубов[41].
Носовая полость разделена частично (например у Minotaurasaurus[19]) или полностью (например у Edmontonia) интерназальной костной перегородкой (англ. internasal septum), которая контактирует с крышей черепа, тем самым разделяя парные дыхательные пути. В образование данной костной перегородки носа переменно вносят вклады носовая и лобная кости[33]. Кроме того, у монгольских анкилозаврид мелового периода (пинакозавр и сайхания) были обнаружены свёрнутые структуры из тонкой кости в полости носа — турбиналии. Они присутствуют в носовых ходах многих млекопитающих. Тереза Марианская назвала их максиллотурбиндлиями и назотурбиналиями. Были обнаружены и более тонкие турбиналии, сравнимые с этмотурбиналиями млекопитающих. Автор описывает их образование за счёт окостенения турбиналий разнообразных пазух: верхнечелюстных и пердчелюстных[42]. Анкилозавриды, в отличие от нодозаврид, в целом имеют необычные и сильно расходящиеся носовые полости, которые проходят через всю морду. Точное функции сложных носовых ходов, пазух и турбиналий анкилозаврид в настоящее время неизвестны[43]c.142. Общий план строения носовой области, подобный анкилозаврам, не был отмечен ни для одной другой группы динозавров[42].
Верхняя челюсть
.jpg.webp)
Передняя часть морды (лат. rostrum) образованна предчелюстной костью (premaxillae). Она формирует клюв, наружные ноздри (и связанные с ними преддверие носа) и придаточные пазухи носа (если они есть), а также частично закрывает ротовую полость[3]c.368. Хотя предчелюстная кость варьируется в зависимости от вида, она часто занимает половину (до 50 %) от общей длины передней части морды[15][3]c.368. В нижней части предчелюстная кость образует большую часть томиума (лат. tomium) — острого края клюва. Этот гребень идёт назад по обе стороны челюсти через предчелюстной-верхнечелюстной контакт и сливаться с началом зубного ряда верхней челюсти (например у Edmontonia), либо заканчивается в положении, отклоняясь в бок от начала верхнечелюстного зубного ряда (например у Ankylosaurus)[3]c.368.
На переднем конце предчелюстной кости, в средней её части, имеется вырезка (incisura premaxillaris), соответствующая по форме выступу на верхней стороне нижней челюсти[26][3]c.368. При виде снизу на предчелюстной кости может присутствовать или отсутствовать срединная продольная борозда (англ. interpremaxillary notch), разделяющая её на две части, и свидетельствующая о контакте двух половинок кости. Нижняя поверхность предчелюстной кости образует часть вторичного нёба. Задние части предчелюстных костей контактируют со сошниками по средней линии[3]c.368.
Верхнечелюстные кости (лат. maxilla) передним краем сочленяются с предчелюстной костью, задним краем соединяются со скуловыми и слёзными костями[26], а также контактирует с эктоптеригоидами и нёбными костями[27]. Верхнечелюстная кость является основным элементом, вмещающим верхний зубной ряд. Характерной особенностью анкилозавров является изгиб зубного ряда внутрь от наружных боковых краёв верней челюсти[1], тем самым разделяя нёбную поверхность на щёчную и язычную части[3]c.370. Между зубными рядами, язычная часть верхней челюсти в виде гладкой сводчатой поверхности, образует углубление внутренних носовых отверстий — хоан[3]c.370. Сошник (англ. vomers) же образует срединную костную перегородку левой и правой хоан, и простирается от предчелюстной кости до контакта с птеригоидами. Представляет собой парные узкие пластинки, плотно прижатые друг к другу, контакт между которыми у некоторых видов почти не различим[33]. Поверхность язычной части верхней челюсти участвует в образовании вторичного нёба (англ. secondary palate). Оно представляет собой горизонтально плоскую пластину и образует дно носовой полости, отделяя её от ротоглотки[3]c.370. Это позволяло животному жевать пищу и дышать одновременно. Вторичное нёбо простирается от переднего кончика верхней челюсти до точки, расположенной прямо над орбитами. Задний край вторичного нёба образует переднюю и боковую границы хоан[17]. В отличие от единственного вторичного нёба у некоторых нодозаврид, образованного выступами предчелюстных и верхнечелюстных костей (англ. rostrodorsal palatal shelf), у многих анкилозаврид оно представляет собой двудольную структуру, второй элемент которого простирается вперёд от птеригоида и нёбных костей (англ. caudoventral palatal shelf) и частично лежит под впереди лежащим основным нёбным выступом[10][41]. В задней части верхней челюсти, с каждой стороны заднего конца сошника, располагаются нёбные кости (англ. palatine). Задним краем нёбные кости граничат с птеригоидами и эктоптеригоидами, наружными боковыми краями с верхнечелюстной костью[36][33].
.jpg.webp)
alv — альвеолы (alveolus); bas — основная затылочная кость (basioccipital); bpt — базиптеригоидный отросток (basipterygoid process); bs — базисфеноид (basisphenoid); ch — хоаны (choana); dpf — нисходящий лобный отросток (descending process of frontal); ee — эктетмоид (ectethmoid); fm — большое затылочное отверстие (foramen magnum); ls — латеросфеноид (laterosphenoid); ltf — латеральное височное отверстие (laterotemporal fenestra); mx — верхнечелюстная кость (maxilla); mx tom — верхнечелюстной томиум (maxillary tomium); nc — носовые полости (nasal canal); ns — носовая перегородка (nasal septum); nuc — затылочный гребень (nuchal crest); oc — затылочный мыщелок (occipital condyle); of — обонятельная область носовой полости (olfactory region of nasal canal); orb — глазница или орбита (orbit); orbs — орбитосфеноид (orbitosphenoid); pal — нёбная кость (palatine); pal apt — нёбное отверстие (palatal aperture); parocc — парокципитальный отросток (paroccipital process); pmx — предчелюстная кость (premaxilla); pmx tom — предчелюстной томиум (premaxillary tomium); ps — парасфеноид (parasphenoid); pt — крыловидная кость (pterygoid); ptq — квадратный отросток птеригоида (quadrate ramus of pterygoid); ptv — межптеригоидная пустота (interpterygoid vacuity); ptw — птеригоидные фланги (pterygoid wing); q — квадратная кость (quadrate); qh — головка квадратной кости (quadrate head); qj — квадратично-скуловая кость (quadratojugal); qjh — нижний заглазничный шип (quadratojugal horn); socc — верхняя затылочная кость (supraoccipital); sqh — верхний заглазничный шип (squamosal horn); v — сошник (vomer).
Нижняя челюсть
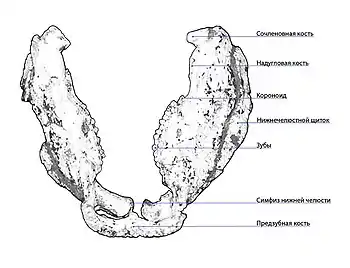
Нижняя челюсть у анкилозавров сравнительно изящная, с укороченным ретроартикулярным отростком и уменьшенной венечной костью, а также вертикально уплощённой предзубной костью. Её характерной особенностью является закрытие наружного нижнечелюстного отверстия (англ. mandibular fenestrae) и наличие небольшого симфиза — переднего соединения между двумя половинами нижней челюсти, в котором оба элемента никогда не срастаются между собой[1].
За исключением предзубной, все остальные кости нижней челюсти, включая зубную, пластинчатую, угловую, надугловую, венечную, предсочленовную и сочленовную, являются парными[38].
Предзубная кость (англ. predentary) образует широкий, уплощённый беззубый подклюв. Имея некое подобие полукруга, кость соединяет парные зубные кости. Верхняя и нижняя поверхности могут быть перфорированы множеством отверстий и неглубоких ямок. Зубная кость (англ. dentary) представляет собой единственную нижнечелюстную кость, на которой есть зубы. Зубной ряд нижней челюсти находится в пределах альвеолярного гребня, который проходит по всей длине зубной кости. У разных родов он варьируется от прямого (Gargoyleosaurus) до вогнутого внутрь (Euoplocephalus)[38]. Спереди зубная кость резко уплощается, образуя изгибающиеся внутрь части[20]. Передние концы зубных костей левой и правой ветвей нижней челюсти соединяются вместе, образуя симфиз нижней челюсти (англ. mandibular symphysis). При виде с внешней стороны зубная кость сочленяется задним краем с надугловой костью, задненижним краем с угловой костью. С внутренней стороны нижний край зубной кости контактирует с пластинчатой костью (англ. splenial). Она представляет собой продольно длинный, тонкий костный элемент. Задневерхнем краем пластинчатая кость образует дно аддукторной ямы и соединяется с передненижним краем предсочленовной кости. От переднего края, в месте контакта зубной и пластинчатой костей, к нижнечелюстному симфизу идёт продольная нижнечелюстная борозда (англ. mandibular sulcus)[38] или же Меккелева борозда (англ. Meckelian groove), в которой находились неокостеневшие остатки меккелева хряща, с наружной стороны имеется хорошо выраженная борозда для предзубной кости (англ. predentary sulcus)[20].
Угловая кость (англ. angular) располагается вдоль нижнего края с наружной стороны нижней челюсти. Характерная для анкилозавров угловая кость закрыта костным орнаментом, который скрывает морфологию сочленения с соседними элементами[38]. Этот костный орнамент представлен длинной узкой остеодермой (mandibular caputegulum), частично сросшейся с челюстью по её нижне-боковому краю. У некоторых видов она распространяется вверх на боковую поверхность челюсти, например у Saichania. Она может простираться почти по всей длине нижней челюсти продольно (Edmontonia), либо же покрывать только половину длины нижней челюсти (Tarchia, Saichania)[19]. Однако у молодых особей Pinacosaurus grangeri, у которых отсутствует обширное развитие орнамента, виден боковой профиль угловой кости с удлинённым нижним краем[13][38].
Надугловая кость (англ. surangular) расположена в задней части нижней челюсти, способствуя образованию боковой (наружной) стенки аддукторной ямы и ретроартикулярному отростку (англ. retroarticular process)[38]. Почти все анкилозавры обладают относительно коротким и глубоким ретроартикулярным отростком, за исключением Gargoyleosaurus, у которого он достаточно длинный и тонкий[23]. Кроме этого, надугловая кость с внешней стороны соединяется передним краем с венечной костью, образуя венечный отросток (англ. coronoid process)[6][38]. Этот отросток идёт вверх от верхней поверхности нижней челюсти за зубным рядом. Служит местом прикрепления мышц, закрывающих челюсть. Ретроартикулярный и венечный отростки нижней челюсти анкилозавров слабо развиты по сравнению с орнитоподами и цератопсами, что свидетельствует об ограниченной силы укуса[44]. Венечная кость (англ. coronoid) своим передним концом налегает на зубную кость, сзади в верхней части она граничит с надугловой костью, а внизу образует передний край внутренней стенки аддукторной ямы[45], в то время как предсочленовная кость образует её задний край. Данная кость трудно идентифицируемая и редко наблюдается у анкилозавров. Аддукторная яма (англ. adductor fossa) служит местом прикрепления основных приводящих мышц, закрывающих челюсть[38]. Предсочленовная (англ. prearticular) и сочленовная (англ. articular) кости находятся в самом заднем положении «ветвей нижней челюсти» (англ. mandibular ramus), и в сочетании с надугловой костью также вносят вклад в формирование ретроартикулярного отростка[23]. Ограниченная надугловой (с наружной стороны) и предсочленовной (с внутренней стороны) сочленовная кость вмещает квадратично-сочленовную ямку (англ. quadrate articular fossa)[38].
У некоторых анкилозавров в области щёк имелись щёчные пластинки, сохранившиеся в исходном положении, непосредственно с щёчной стороны зубных рядов (например у Panoplosaurus[46] и Edmontonia[47]). Более крупная, овальная, удлинённая продольно пластинка была расположена спереди, а несколько более мелких элементов — сзади. Эти пластинки не срастаются ни с нижней, ни с верхней челюстями[47], что указывает на некоторую ограниченную подвижность внутри щеки[46].
Подъязычный аппарат (англ. hyoid) анкилозавров изучен плохо[38]. У некоторых видов (например Pinacosaurus) известно наличие тонких треугольных косточек в горле динозавра — paraglossum. Этот набор костей, полученных от жаберных дуг четырёхногих рыб, обеспечивает место крепления мышц языка. У других родов анкилозавров — Saichania, Edmontonia, также были обнаружены параглоссалии. Эти кости у акнилозавров были намного больше и прочнее, чем те, что наблюдаются у современных птиц, это может свидетельствовать о наличии мускулистого языка[15][48]. В заднебоковых углах подъязычной области находятся небольшие роговидные выступы, которые сочленяются с палочковидными цератобранхиалами (англ. ceratobranchial)[38]. В области горла у некоторых видов известны многочисленные остеодермы диаметром от 5 до 15 мм (англ. gular ossicle; горловые косточки)[20][18].

- 1 — Альвеолярный гребень (alveolar border)
- 2 — Венечная кость (coronoid bone)
- 3 — Зубная кость (dentary bone)
- 4 — Меккелева борозда (Meckelian groove)
- 5 — Надугловая кость (surangular bone)
- 6 — Нижнечелюстная бороздка (predentary sulcus)
- 7 — Нижнечелюстное отвестие (caudal intermandibular foramen)
- 8 — Предзубной бугор (predentary tubera)
- 9 — Пластинчатая кость (splenial bone)
- 10 — Предсочленовная кость (prearticular bone)
- 11 — Ретроартикулярный отросток (retroarticular process)
- 12 — Симфиз нижней челюсти (mandibular symphysis)
- 13 — Сочленовная кость (articular bone)
- 14 — Угловая кость (angular bone)
Зубы
.jpg.webp)
Зубы анкилозавров были относительно небольшими по сравнению с размером тела[38]. Размер зубов варьируется от 3 до 17,5 мм в ширину[46]. Зубы нодозаврид как правило больше по размеру (от 9 мм у Sauropelta до 17,5 мм у Europelta), чем у анкилозаврид (например у Euoplocephalus 3-5 мм; Saichania 7-8 мм; Gobisaurus 9,5 мм)[46][49]. Число верхнечелюстных зубов у большинства анкилозавров составляет около 20: у анкилозавра наибольшее количество верхних зубов (34—35), тогда как у Liaoningosaurus наименьшее число (около 10)[50]. Количество зубов, вероятно, увеличивается во время роста[51], а количество постоянных зубов также варьируется между видами[52][50].
Предчелюстные зубы редко встречаются у анкилозавров, у тех родов, где они присутствуют (Gargoyleosaurus, Pawpawsaurus, Silvisaurus), они обычно имеют коническую форму и слегка загнуты назад, с гладкой коронкой[41]. Зубы верхней и нижней челюсти имели листообразный вид, сжатые с боков, с зубчиками по краю коронки[10][41]. Количество зубчиков у разных видов разнится от — 5 до 17 на зуб[10][49]. Морфология коронки может быть гладкой или рифлёной, вертикальные канавки обычно соответствуют расположению зубцов[41]. У некоторых нодозаврид, например у Edmontonia, на зубах может отсутствать отчётливый, широкий, гладкий центральный гребень, доходящий до вершины коронки. Зубы нодозаврид и анкилозаврид имеют округлые, вздутые основания коронки. Одиночные прямые корни имеют цилиндрическую форму и слегка сужаются к низу[53]. У основания коронки зуба, чуть выше корней, может присутствовать выпуклость в виде полочки, или цингулум (лат. cingulum). Он чаще встречается у нодозаврид, чем у анкилозаврид, однако даже у нодозаврид цингулум вариабелен и может отсутствовать в одном и том же зубном ряду. У анкилозаврид основание коронки обычно вздуто (цингулум присутствует в редких случаях). Зубы полакантид являются самыми примитивными среди анкилозавров, набухшая коронка и цингулум у них отсутствует[54]. Поскольку зубы анкилозавров имеют очень консервативную морфологию и ограниченное использование в таксономии[55], их идентификация возможна в лучшем случае на семейном уровне[10].
Верхние и нижние зубы у большинства анкилозавров однотипны[55]c.269, хотя у некоторых форм (например у Gargoyleosaurus) предчелюстные, верхнечелюстные и нижнечелюстные зубы имеют тонкие морфологические различия[46]. Прикус встречается редко[56], а морфологические вариации положения зубов верхней и нижней челюстей относительно друг друга остаются неоднозначными[55][41].
У ранних представителей нодозаврид зубы ещё присутствовали на предчелюстной кости, но они по-видимому были не очень функциональны и у более поздних представителей семейства остаются лишь гребни, соединяющие предчелюстную кость с началом каждого зубного ряда, а у анкилозаврид исчезающие вовсе[57][58]. Например зубы присутствуют у Gargoyleosaurus, некоторых нодозаврид, таких как Pawpawsaurus и Silvisaurus, и у примитивных анкилозаврид Cedarpelta[59], но отсутствуют на предчелюстных костях более развитых Gastonia sp. и Edmontonia[60][59][53].
Замещающие зубы можно увидеть у различных анкилозавров, включая Tarchia gigantea, Pinacosaurus grangeri и Gargoyleosaurus. Независимо от их альвеолярного положения замещающие зубы всегда являются лингвальными (находятся на язычной стороне) по отношению функционирующему зубу, однако степень их прорезывания вдоль зубного ряда варьируется у разных таксонов[61]. На зубах анкилозавров не обнаружено явных следов стирания, подобных сильным повреждениям на коронках протоцератопсов или гадрозавров. Однако стесывание зубчиков и небольшие следы износа известны[62][46].
Анкилозавры (особенно нодозавриды[3]) обладали необычными извилистыми рядами предчелюстных, верхнечелюстных и нижнечелюстных рядов. Это происходит из-за заметной кривизны зубных рядов как в горизонтальной, так и в вертикальной плоскостях и приводит к очень сложным механизмам челюстей, позволяющим полноценный контакт зубов, по крайней мере у некоторых видов[63][56][61]. Например у большинства анкилозавров в месте соединения предчелюстной и верхнечелюстной костей верхний зубной ряд постепенно изгибается внутрь, образуя щёчную выемку (англ. buccal emargination), а задний зубной ряд расходится в сторону щеки. Однако у некоторых анкилозаврид (например Euoplocephalus, Saichania chulsanensis) передняя часть зубного ряда верхней челюсти также расходится в сторону щеки, образуя зубной ряд в форме песочных часов[3]c.370. Ориентация передних зубов верхней челюсти анкилозавров, как у нодозаврид, так и анкилозаврид, вертикальная, тогда как задняя половина или одна треть зубов верхней челюсти имеет тенденцию ориентироваться заметно наружу[61]. Зубной ряд заканчивается задолго до орбиты (например Gastonia, Ankylosaurus, Scolosaurus, Panoplosaurus, Edmontonia) или же ниже орбиты (Saichania chulsanensis; ранее Tarchia)[17].
Посткраниальный скелет
Многие особенности строения посткраниального скелета анкилозавров связаны с ношением и поддержкой остеодермального панциря. Все элементы посткраниального скелета, как правило, имели массивное строение. Сравнительно лёгкой конструкцией обладали только анкилозаврид Pinacosaurus и нодозаврид Sauropelta[4].
Позвонки
Позвоночный столб делится на 7 или 8 шейных позвонков (англ. cervical), примерно от 12 до 18 спинных позвонков (англ. dorsal) и 3 или 4 крестцовых позвонка (англ. sacral) в области таза. От одного до трёх крестцовых позвонков срастаются с последними двумя-девятью[49]спинными позвонками, образуя структуру, называемую синсакрумом. Хвост имеет от 20 до 24 позвонков (англ. caudal) у анкилозаврид и до 40 у нодозаврид (для полакантид это число неизвестно)[41][64].
Шейные позвонки короткие, часто бывают равны ширине и длине, для удерживания достаточно большого и массивного черепа. Напротив, спинные позвонки обычно длиннее. Кроме того диапофизы (или поперечные отростки), рядом с которыми рёбра прикрепляются к позвонкам, наклонены вверх под углом между 30 и 50 градусами. Этот высокий угол заставляет рёбра выгибаться наружу, образуя бочкообразное тело[64]. Последние четыре или более ребра часто неподвижно срастаются с позвонками. Это слияние, по-видимому, связано с чрезвычайно широким тазом. Хвостовые позвонки удлинены, за исключением области около таза, где они намного больше в ширину, чем в длину. У эволюционно продвинутых анкилозаврид хвостовые позвонки в последней половине хвоста видоизменены, формирующие жёсткую «рукоять» для большой костяной «булавы» на конце хвоста. Они дополнительно укрепляются окостеневшими сухожилиями. Булава образована слиянием нескольких крупных остеодерм с последними несколькими хвостовыми позвонками. У нодозаврид, полакантид и возможно шамозаврин, хвостовые позвонки не модифицированы для ношения костяной булавы. Вместо этого хвост был длинным и тонким[64].
Каждый позвонок образован центральным элементом — тело позвонка (лат. centrum), в форме цилиндра, и невральной дугой (англ. neural arch) в верхней части тела позвонка, дугообразной формы. Невральные дуги позвонков у анкилозавров низкие, по сравнению с другими динозаврами, у которых они имеют высокие остистые отростки, вероятно, для увеличения поверхности прикрепления связок, в том числе и для поддержки хвоста в горизонтальном положении. Не исключено, что у анкилозавров, как и у крокодилов, эту функцию выполнял и остеодермальный панцирь[4][65]. Между телом позвонка и невральной дугой находилась полость — позвоночное отверстие (лат. foramen vertebrale). Эти отверстия в позвоночном столбе образовывали позвоночный канал (англ. neural canal)[34], через который проходил спинной мозг. Некоторые меловые анкилозавры (например Edmontonia) демонстрируют высокий и узкий нервный канал[66]. Однако известные позвонки юрских анкилозавров, таких как Myrmoorapelta maysi имеют нервные каналы, которые относительно несильно вытянуты вверх[67]. Даже у анкилозавров раннего мела, таких как полакантиды, это удлинение остаётся умеренным[68]. Все отделы позвоночного столба (шейные, спинные, крестцовые и хвостовые) сочленяются между собой при помощи передних сочленовных отростков, или презигапофизов (лат. praezygapophyses), и задних сочленовных отростков, или постзигапофизов (лат. postzygapophyses). В верхней части невральной дуги вертикально поднимаются остистые отростки (англ. neural spine). Они служили местом прикрепления для мышц и связок, идущих поверх позвоночника. Шейные, спинные и крестцовые позвонки соединяются с рёбрами посредством поперечных отростков (англ. transverse process) и боковых отростков (англ. parapophysis) по бокам их дуг и тел соответственно[34]. Нижняя часть поперечных отростков — диапофиз (лат. diapophysis; во множестве научной литературе однако, им называют весь поперечный отросток) — образует поверхность, с которой срастается одна из головок ребра, тогда как вторая головка сливается с парапофизами[25]. У большинства птицетазовых динозавров поперечное сечение верхних поперечных отростков имеет треугольную или Т-образную форму, что отличает их от позвонков анкилозавров, у которых они уплощены вертикально, и отсутствует диапофизная бороздка, которая принимает головку ребра. Ни у одной другой группы птицетазовых она не известна[69]. Хвостовые позвонки в нижней их части имели отростки — шевроны (англ. chevron), служившие защитой от повреждений важные элементы хвоста, такие, как нервы и кровеносные сосуды, когда животное опирается на хвост или ударяет им по твёрдой поверхности. Диагностирование позвонков внутри группы анкилозавров возможно только на семейном уровне[3].
Шейные позвонки
.png.webp)
Первый шейный позвонок — атлант (лат. atlas), имеет строение, отличное от прочих шейных позвонков, в связи с участием в подвижном сочленении с затылочной костью черепа. Он состоит из трёх отдельных элементов: неврапофиза (англ. neurapophyses; невральная дуга), интерцентра (лат. intercentrum) и тела позвонка. Неврапофиз и интерцентр прочно сливаются, а у некоторых родов анкилозаврид атлант может также сливаться с эпистрофеем (лат. axis; второй шейный позвонок), как и у неоцератопсий[70]. Это сращивание передних шейных позвонков, называемое syncervical, обеспечивает более надёжное сочленение с тяжёлым черепом[71]. По сравнению с другими шейными позвонками, атлант имеет большое позвоночное отверстие и крошечный остистый отросток. Эпистрофей имеет прочную невральную дугу с коротким остистым отростком и отличается от других позвонков наличием зубовидного отростка (англ. odontoid), которым он сочленяется с задней поверхностью атланта[41].
У последующих за эпистрофеем шейных позвонков тела цилиндрической формы, их длина меньше или равна ширине. Остистые отростки ориентированы продольно, отклоняются назад и часто имеют V- или U-образную форму при виде сверху[34], хотя могут быть выражены межвидовые вариации[70]. Поперечные отростки шейных позвонков прочные и расположены горизонтально. Суставные отростки каждого позвонка выходят далеко за пределы межпозвонковых сочленений, предполагая, что тела позвонков были разделены толстым хрящевым диском[70]. Один или несколько шейных позвонков могут иметь смещённые межпозвоночные суставные поверхности. Эти неправильные позвонки обычно встречаются в середине шейного ряда. В результате этих не типичных позвонков сочленённая шейная область образует небольшую сигмовидальную кривую[72].
В отличие от других шейных позвонков, ребро атланта одноглавое. Оно может быть коротким и крепким (Saichania) или длинным и тонким (Sauropelta)[41]. Все остальные шейные рёбра двуглавые, а в задней половине шейного отдела рёбра могут срастаться поочерёдно с позвонками[70]. У некоторых видов шейные рёбра не сращиваются с позвонками, например у анкилозаврид Saichania и Ankylosaurus[73].
Как и у примитивных архозавров, у эволюционно раннего рода Gargoyleosaurus был обнаружен проатлант (proatlas)[74] — парная плоская кость, треугольной формы, которая сочленяется с затылочным мыщелком и невральной дугой атланта. Основная функция кости состояла в укреплении затылочного сустава. Данный элемент ограничивал подвижность головы, движение которой и так было ограничено короткой шеей, в результате чего анкилозавры, по-видимому, потеряли проатлант, как и ряд других групп птицетазовых динозавров[71]. В дополнение к этому, из-за его небольшого размера и тонкого строения, проатлант легко теряется во время захоронения скелета и потому в окаменелостях встречается редко[75].
Спинные позвонки
.v2.jpg.webp)
Тела спинные позвонков становятся длиннее их ширины, имеют круглые суставные поверхности. Парапофиз первых спинных позвонков отличается от шейных. Он сдвигается вверх и расположен на стыке нервральной дуги и тела позвонка, и является переходным между шейным и спинным позвонками. Помимо расположения парапофиза и поперечных отростков, спинные позвонки можно отличить от шейных на основании морфологических различий остистых отростков, которые также ориентированы поперечно и наклонены назад, но представляет собой тонкую пластинку (при виде спереди или сзади). Поперечные отростки достаточно широкие и более длинные, чем в шейных позвонках. Ориентация, при которой они отходят из нервной дуги, варьируется от горизонтальной до угла равного 45° (задние спинные позвонки имеют больший угол, чем передние)[34]. Наклон поперечных отростков спадает ближе к крестцу. Как и у стегозавров, передние сочленовные отростки образуют U-образную впадину, при этом задние сочленовные отростки образуют стержень, который входит в это углубление, тем самым соединяя позвонки[70].
В то время как у большинства анкилозавров передние и задние межпозвоночные суставные поверхности тел варьируются от плоских до вогнутых, у некоторых родов нодозаврид есть небольшая выпуклость на переднем межпозвонковом сочленении, которая вписывается в сопутствующее углубление на предыдущем позвонке. Эта особенность была интерпретирована как остаток эмбриональной хорды[70][6].
Последние два-девять спинных позвонков соединяются, образуя переднюю часть крестцового стержня — синсакрума[6]c.151[49]. Эта область характеризуется наличием остистых отростков, сросшихся в единую пластинку, которая соединяется с остистыми отростками крестца[70].
Грудная клетка
От каждого спинного позвонка по обе стороны отходят спинные рёбра. Два или три передних ребра короткие, одно или несколько рёбер могут контактировать с коракоидом лопатки[70]. Спинные рёбра, расположенные дальше от шейного отдела, становятся все более крупными и крепкими, и как это часто бывает у птицетазовых динозавров, стержень ребра начинает изгибаться относительно основания ребра[76]. У большинства анкилозавров передние спинные рёбра имеют резко дугообразную форму, напротив, задние спинные рёбра имеют широкую полукруглую форму[73]. В сочленении рёбра образуют большую и очень широкую бочкообразную грудную клетку. Рёбра последних нескольких спинных позвонков расширяются на концах, чтобы контактировать с увеличенной преацетабулярной частью подвздошной кости[70]. Кроме того у рода Saichania обнаружено наличие широких пластинчатых костных межрёберных отростков, которые можно увидеть на средней части спинных рёбер. Эта особенность характерна только для Saichania[15]. Отростки были интерпретированы как средство повышения жесткости туловища у некоторых азиатских анкилозаврид, которая могла служить опорой для более длинного хвоста[49].
Грудина (лат. sternum) у анкилозавров, как и у многих других групп динозавров, так же известна. Она занимает отдел передней стенки грудной клетки. Вместе с рёбрами защищает от внешнего воздействия лёгкие, сердце. У анкилозаврид (Euoplocephalus, Pinacosaurus grangeri и Talarurus) она более окостеневшая, по сравнению со стегозаврами, состоит из двух сросшихся пластинок, объединённых в ромбовидную структуру, с парой заднебоковых выступов[77]. Среди нодозаврид, таких как Panoplosaurus, Sauropelta и Silvisaurus, пластинчатые элементы не сращиваются между собой. Некоторые нодозавриды также имеют мечевидные элементы (англ. xiphisternal) — длинные, тонкие кости в задней части грудины, с многочисленными отверстиями и выпуклыми зубчатыми боковыми краями. Левые и правые половинки соприкасались между собой, c грудиной и по меньшей мере c тремя рёберными хрящами. Морфология грудины Saichania уникальна среди анкилозавров, с широкой единой центральной областью, двусторонним набором парасагиттальных краниальных отростков и длинным хвостовым сегментом, который остаётся не слитым воедино. Кроме того, четыре или пять пар спинных рёбер сочленяются с грудиной — особенность, ранее неизвестная среди птицетазовых динозавров[78].
Крестцовые позвонки
_and_Edmontonia_longiceps_(NMC_8531).jpg.webp)
Характерной особенностью позвоночного столба анкилозавров является образованный крестцовыми позвонками сложно устроенный крестец — синсакрум (лат. synsacrum)[1]. Он включает последние спинные, крестцовые и иногда первые хвостовые позвонки, их количество разнится у разных видов[4][10]. Межпозвонковые грани тел всех спинно-крестцовых и крестцовых позвонков срастались у взрослых особей в единый стержень (не сросшиеся крестцовые позвонки известны у скелетов молодых особей), но тела последних крестцовых позвонков могли не сращиваться[34]. Как и у стегозавров, нервный канал в передней половине крестца заметно расширен[70].
Самый первый позвонок синсакрума имеет передний сочленовный отросток, который соединяется с последним спинным позвонком. Остистые отростки всех позвонков крестца сливаются в единую непрерывную вертикальную пластинку, так что передние и задние сочленовные отростки становятся нечёткими[34].
Начальные позвонки имеют более длинные, уплощённые вертикально рёбра. От крестцовых позвонков отходят более короткие крестцовые рёбра[34]. Поперечные отростки крестцовых позвонков приобретают более горизонтальную ориентацию, имеют удлинённую форму и сливаются с крестцовыми рёбрами по всей длине своей нижней стороны[70]. Рёбра контактируют друг с другом на дистальных концах, соединяясь с подвздошной костью. Часть крестцовых рёбер (обычно 1 и 2) образовывают нижний край вертлужной впадины (например у Gargoyleosaurus, Gastonia и Edmontonia)[79]. Концы поперечных отростков последних позвонков синсакрума контактировали или срастались с подвздошной костью[34][4].
Хвостовые позвонки
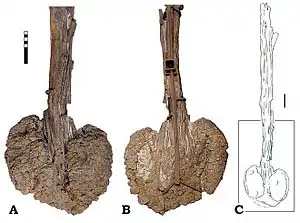
Первые хвостовые позвонки часто сливаются вместе с крестцом, внося свой вклад в образование синсакрума. Длина тел начальных хвостовых позвонков короче их ширины и постепенно увеличивается к середине хвоста. Форма тел передних хвостовых позвонков варьируется от эллипсоидной до круглой, которая затем становится квадратной или шестиугольной по направлению к концу хвоста[70][78].
Хвостовые позвонки включают выступающие вверх остистые отростки и выступающие вниз шевроны (гемальные отростки, присутствуют по всему хвосту). У начальных хвостовых позвонков поперечные отростки и рёбра по прежнему достаточно длинные, уменьшаются к середине хвоста и заканчиваются перед 15-м позвонком[70][34]. Начальные хвостовые рёбра обычно имеют горизонтальную ориентацию, тогда как далее назад эти элементы меняют конфигурацию с небольшим наклонном вниз[78]. Презигапофизы отделены друг от друга (в отличие от спинных позвонков, которые соединяются по средней линии) и представляют собой пальцеобразные выступы невральной дуги. Остистый отросток у анкилозаврид обычно тонкий (например у Euoplocephalus и Talarurus), у нодозаврид он может быть более широким (например у Polacanthus). Гемальная дуга часто сливается с задним нижним краем тела позвонка, а длина гемального отростка примерно равна длине остистого отростка[80].
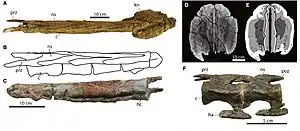
Задние хвостовые позвонки у базальных анкилозавров (например Mymoorapelta), базальных анкилозаврид (например Gastonia) и нодозаврид (например Nodosaurus, Sauropelta) широкие, более уплощённые сверху вниз относительно передних хвостовых позвонков[80]. Остистый отросток уменьшен по сравнению с передним хвостовым отделом. Гемальные отростки также уменьшены и приобретают округлую форму (например Hungarosaurus). Презигапофизы не выходят за передний край тела позвонка более чем на 25 % от общей длины тела (например Nodosaurus, Mymoorapelta), что делало заднюю часть хвоста этих анкилозавров гибкой[80].
Задние хвостовые позвонки анкилозаврин (Ankylosaurinae) уникальны среди анкилозавров. У нодозаврид и базальных анкилозаврид переход от передних к задним хвостовым позвонкам более плавный, в то время как у анкилозаврин этот переход резкий и происходит примерно в середине хвостового ряда[81][80]. Тела задних хвостовых позвонков анкилозаврин становятся более удлинёнными в продольном направлении[82]. Они плотно сцепляются друг с другом, образуя серию взаимосвязанных V-образных частей при виде сверху, за исключением двух образцов рода анкилозавр. С чем связаны отличия этих двух образцов от базального состояния, обнаруженного у других анкилозаврин неизвестно[80]. Презигапофиз каждого позвонка перекрывает соседний передний позвонок не менее чем на 50 % своей длины, в отличие от базальных анкилозаврид, нодозаврид и базальных анкилозавров, где перекрытие составляет около 25 % длины тела позвонка. Модифицированный остистый отросток и постзигапофизы предыдущего позвонка сливаются и полностью заполняют пространство между презигапофизами[83][80]. Поперечные отростки также отсутствуют на большинстве позвонков в этой области, хотя небольшие бугорки могут присутствовать на нескольких первых позвонках «ручки»[81][80]. Гемальные дуги подобным же образом превращаются в плотно сцепляющуюся серию. Гемальный отросток вертикально короткий, но по направлению от передней стороны позвонка к задней достаточно длинный[15][80]. Спереди он раздвоен, а в задней части сужается к концу. Гибкость дистальной части хвоста у анкилозаврин была сильно снижена, а там, где произошло слияние центральных позвонков, гибкости почти отсутствовала[80].
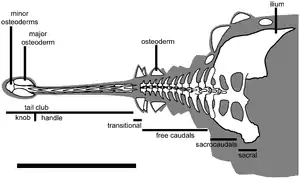
Хвостовая булава
У группы анкилозаврин конец хвоста был дополнен большой костяной булавой. Термин «хвостовая булава» относится не только к большому окостенению, но и ко всей задней структуре хвоста анкилозаврин[84][85][80]. Она состояла из остеодерм, сросшихся с хвостовыми позвонками, поддерживалась и укреплялась окостеневшими сухожилиями — «рукоятью»[83][80]. Позвонки, удерживавшие булаву, не похожи на позвонки любых других четвероногих, несущих похожую конструкцию, они обладали удлинёнными и крепкими презигапофизами, остистыми отростками, постзигапофизами и гемальными дугами. Центральная часть рукояти могла быть частично сужена, что также способствовало жёсткости дистального отдела хвоста у анкилозаврин[80]. Две расположенные сбоку остеодермы (главные остеодермы[83]) образуют основную часть хвостовой булавы, обычно килевидной формы и уплощённые сверху вниз[80]. Различное количество меньших остеодерм образуют заднюю часть выступа. Остеодермы булавы полностью покрывают и закрывают самые дальние позвонки[80]. Лишь немногие экземпляры сохраняют остеодермы вдоль более центральных частей хвоста, например Pinacosaurus[86].
Плечевой пояс
.jpg.webp)
Кости плечевого пояса анкилозавров массивные. Лопатки у анкилозавров вертикально достаточно широкие. У взрослых особей лопатка сливаясь с коракоидом (coracoid) образует скапулокоракоид (scapulocoracoid). Чашеобразный гленоид (glenoid), или плечевая впадина, распространяется как на коракоид, так и на лопатку. Гленоид принимает головку плечевой кости, образуя плечевой сустав[64].
Коракоид и лопатка — ещё одна важная часть, разделяющая три семейства. У семейства полакантид коракоид представляет собой пластинку в форме диска, которая пронизана коракоидным отверстием (coracoid foramen), тогда как обычно это удлинённый прямоугольник у нодозаврид и короткий вертикальный прямоугольник или квадрат у анкилозавридов. Лопатка представляет собой удлинённую структуру, которая слегка изогнута, чтобы соответствовать форме грудной клетки. Верхний и нижний края лопатки могут расходиться или быть параллельны друг другу, в зависимости от вида, например у Gastonia burgei края расходятся к концу лезвия[64].
На внешней стороне лопатки поверх суставной впадины (glenoid fossa) имеется выступ, который называется акромионом (acromion). Он служит местом прикрепления передней плечевой мышцы (scapulohumeralis muscle). У полакантид это довольно широкая кромка, выступающая под углом вниз и наружу, а у анкилозаврид либо короткий выступ, либо вздутие вдоль верхнего края лопатки. Напротив, у нодозаврид это гребень, расположенный ближе к гленоиду и заканчивающийся выступом — псевдоакромионом (pseudoacromion)[64][65]. Положение гребня относительно гленоида варьируется в семействе нодозаврид, что делает лопатку важной составляющей в определении видовых различий внутри семейства[6][64]. Небольшой выступ на нижнем крае лопатки, в центральной части изгиба, вероятно является местом крепления трицепса (enthesis M. triceps longus caudalis; например у Ankylosaurus)[52][34].
Передние конечности
.jpg.webp)
Передние конечности анкилозавров были короче и крепче задних. Плечевая кость (англ. humeral) у анкилозавров короткая и массивная, имеет форму песочных часов. Обычно плечевая кость нодозаврид пропорционально более тонкая, чем у анкилозаврид и полакантид. В верхней части плечевой кости имеется очень заметный выступ, называемый дельтопекторальным гребнем (англ. deltopectoral crest)[64][87], составляющий чуть менее половины длины плечевой кости[34]. Он относительно больше у анкилозаврид, чем отличается от такового у нодозаврид, которые имеют более короткий гребень[6][87]. Хотя этот гребень есть у всех динозавров, у анкилозавров (и других четвероногих динозавров) он особенно хорошо развит. Гребень был местом прикрепления нескольких основных мышц, используемых при передвижении — M. supracoracoideus и M. pectoralis[64][34]. Головка плечевой кости, которая входит в гленоид, расположена больше на задней стороне плечевой кости. Напротив, лучевой (англ. radial condyle) и локтевой (англ. ulnar condyle) мыщелки, с которыми соединяются кости нижней части передних конечностей, более развиты на передней стороне плечевой кости. Эти особенности говорят о том, что предплечья держались не прямыми, как у слонов, а были согнуты[64].
Нижняя часть передних конечностей состоит из большой крепкой локтевой кости и меньшей лучевой кости. Локтевая кость (лат. ulna) похожа на таковую у стегозавров: верхняя часть массивная, сужающаяся вниз. На верхнем конце кости имеется вогнутое углубление для сочленения с лучевой костью[87] и очень высокий локтевой отросток (лат. olecranon), к которому прикреплялись трёхглавые мышцы (M. triceps; мышцы-разгибатели предплечья)[64]. У молодых особей этот отросток локтевой кости низкий, тогда как у большинства крупных анкилозавров он высокий и сильно развит, что говорит о его изменении в течении роста[50].
Лучевая кость (лат. radius) является более короткой. Верхняя часть имеет вогнутую, округлую поверхность, куда входит лучевой мыщелок плечевой кости. На нижней части обычно видны морщинистости, возможно для прикрепления межкостной мышцы. У анкилозаврид соотношение лучевой кости к плечевой составляет примерно 0,60. Соотношение локтевой кости к плечевой значительно выше и обычно составляет примерно 0,85, либо больше, из-за наличия локтевого отростка[87]. Лучевая и локтевая кости продолжались вниз костьми передних стоп.
Кости кистей
Кости кистей (лат. manus) короткие и толстые, чтобы выдерживать вес животного[65]. Скелет передних стоп составляют кости запястья (англ. carpal), пястные кости (англ. metacarpals) и кости пальцев — фаланги (лат. phalanx). За исключением промежуточных у Pinacosaurus sp., запястные элементы у анкилозавров не известны[88].
Среди репрезентативных членов пястные кости с I по IV крепкие (особенно пястные I) и расширены на верхних и нижних суставных поверхностях. V пястная кость значительно короче, менее массивна и (в сочленении) заднебоковую ориентацию. Сесамовидные кости были идентифицированы на передних верхних суставных поверхностях пястных костей I и II у Saichania. Пястные кости контактируют друг с другом в верхней части и расходятся нижней, образуя широко раскинутую кисть[88]. Анкилозавры довольно часто изображаются с данной конфигурацией пястных костей. Другие предположения говорят о том, что кости могли иметь вертикальную, дугообразную конфигурацию, которая отсутствует у других птицетазовых динозавров, предполагая возможность того, что они унаследовали её от общего предка среди тиреофор, у которых кости располагались в форме полукруга[89].
Строение передних и задних стоп долгое время было источником путаницы у анкилозавров из-за плохой сохранности или полного отсутствия этих частей у большинства скелетов, и того факта, что многие виды по-видимому подверглись сокращению числа пальцев и фаланг[64][90]. Те, что известны, короткие и широкие. За исключением Liaoningosaurus, у которого отсутствует пятый палец, все анкилозавры показывают пять пальцев передних стоп[91][69]. Кеннет Карпентер предположил четырёхпалость передних конечностей у продвинутых видов[64], указывая на нодозаврида Edmontonia[92], в то время как Хавьер Переда-Субербиола с коллегами утверждали, что отсутствие пятого пальца в этом таксоне было результатом плохой сохранности[93]. Пальцы оканчивались широкими и плоскими копытообразными когтями (англ. ungual)[64], либо имели треугольные очертания с острыми концами[50].
Тазовый пояс
.png.webp)
Таз (лат. pelvis) анкилозавров значительно отличается от таза других птицетазовых динозавров[64][3][6][10]. У анкилозавров он достаточно широкий, а в продольном направлении достаточно длинный, имеет сильно расходящиеся парные подвздошные кости[34]. Происхождение этого необычного таза связывают с защитой внутренних органов или же с опорой тяжёлой дермальной брони[79].
Подвздошная кость (лат. ilium) очень большая и ориентирована почти горизонтально, охватывая толстую кишку внутренних органов. Образует протяжённую преацетабулярную (переднюю) часть, постацетабулярный (задний) отросток значительно короче. Мышечные рубцы на нижней стороне подвздошной кости показывают, что мышцы для движения задних конечностей были большими и мощными[64][65]. Внутренним боковым краем подвздошные кости контактирует с рёбрами сложно устроенного крестца. У рода Minmi средняя часть подвздошной кости соединяется с крестцом через широкий костный мостик, охватывающий от трёх до пяти позвонков, особенность, ранее неизвестная среди анкилозавров[94]. Лобковая ножка (англ. pubic peduncle), которой крепится лобковая кость, хорошо развита, имеет слегка закруглённый профиль. Седалищная ножка (англ. ischial peduncle) рудиментарна[50].
Лобковая кость (pubis), расположенная ниже подвздошной кости, сводится к небольшому блоку, а может и вовсе отсутствовать, что отличает анкилозавров от других динозавров. Она образует переднюю стенку вертлужной впадины. Напротив, седалищная кость (ischium) остаётся и выступает вниз, что более типично для ящеротазовых динозавров. Седалищная кость у анкилозаврид довольно прямая, но у нодозаврид и полакантид изогнута или изогнута в середине[64]. Запирательный отросток (англ. obturator process) седалищной кости у анкилозавров отсутствует, в отличие от других четвероноих птицетазовых динозавров[94].
Вертлужная впадина (лат. acetabulum), или тазобедренная впадина, представляет собой чашеобразное углубление, образованное подвздошной и седалищной костями, при этом лобковая кость практически отсутствует. У большинства динозавров вертлужная впадина открыта, что позволяет головке бедренной кости частично выходить наружу. Напротив, вертлужная впадина закрыта у нодозаврид и анкилозаврид, а головка бедренной кости больше похожа на шар, так что она образует шарнирное соединение с вертлужной впадиной. Однако у полакантид вертлужная впадина закрыта не полностью, особенно у примитивного рода Mymoorapelta; она также остаётся в виде небольшой щели у более эволюционно продвинутого рода Gastonia. Скорее всего, эти отверстия были закрыты хрящом, образующим чашевидную впадину[64].
Задние конечности
.png.webp)
Бедренная кость (femur) у анкилозавров столповидная и прямая, массивная, удерживающая огромный вес тела. Диафиз (средняя часть кости) имеет большую окружность по сравнению с таковой у двуногих птицетазовых динозавров[65][64].
Головка бедренной кости (femoral) представляет собой выпуклую, шарообразную суставную поверхность, расположенную под большим углом к стержню кости (у некоторых видов он достигает 145 °[50]), а не цилиндрическую, расположенную под прямым углом, как у двуногих динозавров[64]. На противоположной стороне от головки кости расположены черепной (англ. cranial trochanter) и большой (англ. greater trochanter) вертелы. Черепной вертел присутствует у молодых особей, таких как Anoplosaurus, но сливается с большим вертелом у большинства взрослых анкилозавров. Наличие черепного вертела, вероятно, является примитивным признаком анкилозаврид, а также зависит от стадий роста у некоторых таксонов[50]. Четвёртый вертел, к которому прикрепляется главная мышца-сгибатель конечности — musculus caudofemoralis longus, представляет собой шероховатый шишкообразный элемент в средней части стержня бедренной кости, а не крючковидную структуру, наблюдаемую у небольших двуногих динозавров, таких как гипсилофодон[64]. Нижний конец бедренной кости разделён на медиальный (большеберцовый) и латеральный (малоберцовый) мыщелки[50], и опирается на две короткие, но массивные кости голени — большеберцовую и малоберцовую[64][95]. Как правило, у анкилозаврид кости голени составляют менее двух третей длины их бёдер, в отличие от нодозаврид, которые имеют пропорционально более длинные элементы голени[73], около 80-87 % длины бедра[95].
Большеберцовая кость (лат. tibia) более крупная из двух костей голени, верхние и нижние концы кости значительно расширены относительно центральной её части, что является типичным для тиреофорных динозавров. Область прикрепления мышц-разгибателей представляет собой выступающий кнемиальный гребень (англ. cnemial) на переднем крае коленного сустава в верхней части кости[64]. Малоберцовая кость (лат. fibula) тонкая[95]. Голеностопный сустав состоит из астрагала (лат. astragalus) и пяточной кости (лат. calcaneum), которые по мере взросления сливаются у большинства анкилозавров с нижними концами большеберцовой и малоберцовой костей соответственно[3][64][95]. Почему это слияние происходит у взрослых анкилозавров, неизвестно. Как и у всех динозавров, лодыжка анкилозавров представляет собой простое шарнирное соединение. Подобное слияние происходит и у близкородственной группы стегозавров[64].
Кости стоп
Кости стоп (лат. pes) короткие и широкие, адаптированы под большой вес животных[95][4]. Скелет задних стоп состоит из костей предплюсны (лат. tarsus), плюсневых костей (англ. metatarsals) и костей пальцев — фаланг (лат. phalanx). По сравнению с пястными костями передних конечностей, плюсневые кости более удлинённые и тонкие, особенно (если они есть, например у Sauropelta) плюсневые кости V. У Liaoningosaurus длина стопы более чем в два раза превышает длину кисти. В нижней части пястные кости стоп менее расставлены, чем пястные кости кистей[96].
Как и у кистей передних конечностей, количество пальцев на задних конечностях плохо изучено. У нодозаврид по-видимому было четыре пальца (за исключением пятипалого Nodosaurus), тогда как у продвинутых анкилозаврид обычно три (у полакантид это число неизвестно)[64][91]. Е. А. Малеев сообщал, что у анкилозаврида Talarurus имелось четыре пальца на задних конечностях, однако находка состоит из нескольких особей, и неясно, как элементы конечности были связаны в действительности[97]. В настоящее время точное наличие четырёхпалости у анкилозаврид не известно[69].
Фаланги задних конечностей у анкилозавров достаточно короткие, толстые и напоминают фаланги стегозавров или цератопсид[95]. Формула фаланг разнится, например у нодозаврид (Niobrarasaurus, Sauropelta и Europelta) она равна 2-3-4-4-0[91]. Образцы задних стоп Pinacosaurus показывают только три пальца, у которых фаланговая формула 0−3−3/4−3/4−0. Количество фаланг в третьем и четвёртом пальцах задних конечностей может варьироваться от трёх до четырёх на каждом пальце у разных особей одного и того же вида[90]. Кроме того, наличие трёхпалости у обоих эволюционно продвинутых анкилозаврид (Pinacosaurus и Euoplocephalus) и морфологически отдалённого от них рода Liaoningosaurus указывают на то, что это состояние развивалось у анкилозавров независимо по крайней мере дважды в эволюционной истории.
Пальцы задних конечностей оканчивались широкими и плоскими когтями (aнгл. ungual), более похожими на копыта (например у трёхпалого Euoplocephalus фаланги пальцев оканчивались U-образными когтями) или же острыми треугольными когтями (например у Dyoplosaurus, Scolosaurus)[98], следовательно форма когтей может быть полезным признаком для таксономических различий[50].
Остеодермы тела
Основными отличительными и сравнительными характеристиками анкилозавров являются множественные продольные ряды остеодерм, которые присутствуют по всей поверхности шеи, тела, а также боковых сторон хвоста[10][1]. Наиболее известные остеодермы состояли из килевидных или плоских костных пластин, иногда эти остеодермы дополняются шипами на теле и хвосте, а также терминальной булавой на кончике хвоста[64].
Костные пластины

Остеодермы анкилозавров, покрывавшие посткраниальный скелет, обычно имеют овальную или прямоугольную форму, с гребнем на их вершине[99]. Размер пластин мог варьироваться от 2 до 35 сантиметров в диаметре[100], а толщина могла достигать 5 см[97].
Они шли вдоль спины многочисленными продольными и поперечными рядами, однако их точное расположение на теле часто не известно. Обычно, остеодермы расположенные непосредственно на спине за вторым шейным полукольцом, имитируют форму и расположение таковых в полукольце. Между большими пластинами находились маленькие промежуточные остеодермы (англ. interstitial osteoderms), прикрывавшие зазоры[8].
У полакантид панцирь состоит как минимум из двух рядов пластин на шее, овальных, килевидных пластин, расположенных чередующимися рядами вдоль спины, поперечных параллельных рядов на хвосте и атласных треугольных пластин с широким основанием или выступающих по бокам шипов, шеи, тела и хвоста[101]. Панцирь нодозаврид обычно состоит из трёх рядов больших пластинчатых остеодерм на шее, но в остальном он сильно варьируется и может включать длинные шипы, выступающие из шеи и плеч. Нодозавриды также обычно имеют толстые треугольные пластины, выступающие с боков тела, и параллельные ряды килевидных овальных пластин на спине. У анкилозаврид было два ряда шейных пластин и треугольные шипы или пластины, выступающих по бокам шеи, тела и хвоста. Спина обычно покрыта овальными килевидными пластинами, расположенными параллельно (Euoplocephalus) или поперечно параллельными и чередующимися рядами (Saichania)[57][102][101]. Длинные шипы у анкилозаврид отсутствуют, однако у некоторых видов на спине и плечах есть высокие тонкостенные шишки, внешне напоминающие шипы[103].
Шейные полукольца

На шее у анкилозавров было обычно два, иногда три, ряда больших килевидных пластин, вместе образовывающих форму полукольца, покрывающих верхнюю часть шейного отдела и надплечевую область. Шейные полукольца остеодерм являются синапоморфными для группы анкилозавров[104]. Пластины, составляющие плечевой пояс, значительно крупнее, чем пластины шейного пояса. У большинства анкилозаврид (Ankylosaurus, Pinacosaurus, Saichania) морфология первого и второго шейных полуколец схожа, а общая морфология шейных полуколец считается полезной с таксономической точки зрения[34].
Обычно шейные полукольца состоят из двух слоёв: глубокий (нижний) слой неостеодермальной кости и поверхностный (верхний) слой первичных остеодерм, подобных тем, которые встречаются на остальной части тела. Нижняя часть шейного полукольца состоит из сравнительно толстых и плотных костных сегментов (большинство шейных колец имеют шесть сегментов), квадратной или прямоугольных формы, соединёнными зубчатыми швами. Верхняя часть шейных полуколец в большинстве случаев также состоит из шести больших остеодерм, по три на каждую сторону (медиальные, латеральные и дистальные — самые нижние[105])[34]. Иногда вокруг этих основных остеодерм присутствуют многочисленные более мелкие остеодермы. Нижележащий сегмент и вышележащая остеодерма часто бывают слиты, либо слиты частично. Сегменты нижней части всегда имеют гладкую текстуру и больше похожи по внешнему виду на эндохондральную кость, чем на остеодермальную кость, которая обычно имеет ямки или морщинистую форму у анкилозаврид[34].
У разных видов верхние остеодермы полуколец различались. Рода Euoplocephalus и Anodontosaurus обычно имеют овальную форму у основания и киль размером от низкого до среднего; Scolosaurus имел круглую остеодерму с низкими коническими точками; Ziapelta имеет прямоугольные остеодермы с высокими килями. Ещё больше отличаются остеодермы рода Shamosaurus, который имеет сильно направленные вперёд вершины на дистальных остеодермах. Шея не была скована в движениях за счёт зазоров между самой шеей и пластинами. Эти промежутки частично перекрывались небольшими круглыми ороговевшими чешуйками[34].
Тазовый щит

Тазовый щит анкилозавров также относится к области остеодерм. Он представляет собой комплекс нерегулярно расположенных больших и малых костных пластин, слившихся в единое целое и располагался над тазовой областью. Характер тазового щита варьируется в разных таксонах анкилозавров и используется в качестве диагностического признака для различных групп анкилозавров. Морфология тазового щита предлагаются три категории: тазовые щиты 1-й категории имеют не сросшиеся, но плотно сцепленные остеодермы; тазовые щиты 2-й категории имеют сросшиеся остеодермы, образующие розетки, и ограничены поздним юрским и средним меловым периодом Северной Америки и Европы. Тазовые щиты 3-й категории имеют сросшиеся полигональные остеодермы аналогичного размера и встречаются в середине—конце мелового периода Северной Америки[106].
Наличие тазового щита используется в основном для характеристики группы Polacanthidae[106], например у базальных анкилозавров Gastonia и Mymoorapelta, но может присутствовать и у многих представителей семейства нодозаврид — например Taohelong и Polacanthus. Остеодермы таза также сохранились у нескольких поздних североамериканских — сколозавр, и азиатских — пинакозавр и тархия, анкилозаврид. Боковые остеодермы таза, в отличие от центральных остеодерм, имеют высокий киль и имеют треугольный профиль при виде сверху[8].
Шипы
У некоторых представителей семейства нодозаврид на шее и в области плеч остеодермы полуколец имели форму шипов (максимальная высота до 60 см[107]), с различной ориентацией их вершин[103]. Шипы по бокам шеи характеризуются наличием длинной бороздки вдоль их заднего края, функция которой, по-видимому, состоит в том, чтобы находить на передний край следующего шипа, напоминая некое подобие гильотины[108].
На боковой стороне тела, как продолжение шейных шипов, часто встречаются сильно гребневидные (до 20 см), с острыми вершинами, остеодермы (например у Gastonia[109]), идущие назад до кончика хвоста[107].
Хвостовые остеодермы анкилозавров сильно различаются по размеру и морфологии, с характерным общим рисунком, представленным коническими или уплощёнными шипами, окружёнными розеткой из более мелких остеодерм или неравномерно распределёнными зернистыми «косточками» (например у рода Zuul)[110]. У азиатских и североамериканских видов, например Pinacosaurus, Euoplocephalus, Dyoplosaurus и Zuul, сохранившиеся хвостовые остеодермы обычно килевидные или треугольные, расположены рядами, как и на остальной части тела. Азиатские анкилозаврины сохраняют треугольные боковые остеодермы вдоль задней части хвоста, у североамериканских образцов треугольные остеодермы хвоста пока известны только у рода Zuul[8].
Примечания
- Vickaryous, Maryańska, Weishampel, 2004, p. 363
- Туманова, 1987
- Vickaryous, M. K., Maryańska, T., Weishampel, D. B. 2004. Ankylosauria. Pp. 363—392. In D. B. Weishampel, P. Dodson, and H. Osmólska (eds) The Dinosauria, 2nd edition. University of California Press, Berkeley.
- Tumanova, T. A. 2012. Infraorder Ankylosauria. Pp. 310—324. In L. P. Tatarinov, E. I. Vorobyeva and E. N. Kurochkin (eds.). Fossil vertebrates of Russia and adjacent countries. Fossil reptiles and birds. Part 2. Moscow: Geos.
- Туманова, 1987, с. 25
- Coombs, W. P. 1978. The families of the ornithischian dinosaur order Ankylosauria. Paleontology, 21: 143—170.
- Weishampel et al., 2004, pp. 388—390
- Victoria M. Arbour, Jordan C. Mallon. Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris (англ.) // FACETS 2. — 2017. — P. 764–794. — doi:10.1139/facets-2017-0063.
- Weishampel et al., 2004, p. 390
- Coombs, W. P., Maryańska, T. 1990. Ankylosauria. Pp. 456—483. In: D. B. Weishampel, P. Dodson, H. Osmolska (eds.). The Dinosauria. 1st ed. Berkeley: University of California Press.
- Eaton, T. 1960. A new armoured dinosaur from the Creataceous of Kansas. Univ. Kans. Paleontol. Contrib., 8: 1—24.
- Maryańska, T. 1971. New data on the skull of Pinacosaurus grangeri (Ankylosauria). Palaeontologia Polonica 25: 45—53.
- Hill, R. V., Witmer, L. M., Norell, M. A. 2003. A new specimen of Pinacosaurus grangeri (Dinosauria: Ornithischia) from the Late Cretaceous of Mongolia: ontogeny and phylogeny of ankylosaurs. American Museum Novitates 3395: 1—29
- Parish, J. C. 2005. The evolution and palaeobiology of the armoured dinosaurs (Ornithischia: Ankylosauria). PhD Thesis, Oxford University.
- Maryanska, T. 1977. Ankylosauridae (Dinosauria) from Mongolia. Palaeontologia Polonica, 37: 85—151.
- Witmer, L. M., Ridgely, R. C. 2008. The paranasal air sinuses of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria) and their contribution to cephalic structure. The Anatomical Record Advances in Integrative Anatomy and Evolutionary Biology 291 (11): 1362—1388.
- Leahey, L. G., Molnar, R. E., Carpenter, K., Witmer, L. M. & Salisbury, S. W. 2015. Cranial osteology of the ankylosaurian dinosaur for merly known as Minmi sp. (Ornithischia: Thyreophora) from the Lower Cretaceous Allaru Mudstone of Richmond, Queensland, Australia. PeerJ 3: e1475.
- Zheng, W., Jin, X., Azuma, Y., Wang, Q., Miyata, K. & Xu, X. 2018. The most basal ankylosaurine dinosaur from the Albian-Cenomanian of China, with implications for the evolution of the tail club. Scientific Reports 8 (1): 3711.
- Miles, C. A. and Miles, C. J. 2009. Skull of Minotaurasaurus ramachandrani, a new Cretaceous ankylosaur from the Gobi Desert. Current Science 96 (1): 65—70.
- Arbour, V. M., Evans, D. C. 2017. A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA, based on an exceptional skeleton with soft tissue preservation. Royal Society Open Science 4 (5): 161086.
- Arbour, V. M., Burns, M. E., Sullivan, R. M., Lucas, S. G., Cantrell, A. K., Fry, J. and Suazo, T. L. 2014. A new ankylosaurid dinosaur from the Upper Cretaceous (Kirtlandian) of New Mexico with implications for ankylosaurid diversity in the Upper Cretaceous of western North America. PLoS ONE, 9: e108804.
- Arbour, V. M., Currie, P. J. 2016. Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs. Journal of Systematic Palaeontology 14 (5): 385—444.
- Kilbourne, B. & Carpenter, K. 2005. Redescription of Gargoyleosaurus parkpinorum, a polacanthid ankylosaur from the Upper Jurassic of Albany County, Wyoming. Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen, 237 (1): 111—160.
- Vickaryous, M. K., Russell, A. P. and Currie, P. J. 2001. Cranial ornamentation of ankylosaurs (Ornithischia: Thyreophora): Reappraisal of developmental hypotheses. Pp. 318—340. In: K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- Wiersma, J. P., Irmis, R. B. 2018. A New southern Laramidian Ankylosaurid, Akainacephalus johnsoni gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA. PeerJ 6: e5016.
- Туманова, 1987, с. 27
- Vickaryous, Maryańska, Weishampel, 2004, p. 371
- Vickaryous, Maryańska, Weishampel, 2004, p. 373.
- Туманова, 1987, с. 26
- Vickaryous, Maryańska, Weishampel, 2004, p. 372.
- Carpenter, K., Kirkland, J. I., Burge, D., Bird, J. 2001. Disarticulated skull of a new primitive ankylosaurid from the Lower Cretaceous of eastern Utah. Pp. 211—238. In: K. Carpenter (ed.). The armored dinosaurs. Bloomington: Indiana University Press.
- Vickaryous, Maryańska, Weishampel, 2004, p. 374.
- Vickaryous, Maryańska, Weishampel, 2004, p. 375.
- Arbour, V. M., Currie, P. J. 2013. Euoplocephalus tutus and the Diversity of Ankylosaurid Dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA. PLoS ONE, 8: e62421
- Vickaryous, Maryańska, Weishampel, 2004, p. 377.
- Туманова, 1987, с. 33
- Vickaryous, Maryańska, Weishampel, 2004, p. 376.
- Vickaryous, Maryańska, Weishampel, 2004, p. 378.
- A. Ősi, X. Pereda-Suberbiola, and T. Földes. 2014. Partial skull and endocranial cast of the ankylosaurian dinosaur Hungarosaurus from the Late Cretaceous of Hungary: implications for locomotion. Palaeontologia Electronica17.1.1A: 18.
- Coombs, W. P. 1972. The bony eyelid of Euoplocephalus (Reptilia, Ornithischia). Journal of Paleontology 46: 637—650.
- Vickaryous, Maryańska, Weishampel, 2004, p. 379
- Туманова, 1987, с. 31
- Lucas S. G. 2016. Dinosaurs. The Textbook. Columbia University Press. 159 p.
- Mallon, J. C. 2019. Competition structured a Late Cretaceous megaherbivorous dinosaur assemblage. Scientific Reports 9: 1—18.
- Туманова, 1987, с. 41
- Ősi, A., Prondvai, E., Mallon, J. & Bodor, E. R. 2016. Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia). Historical Biology 29 (4): 539—570. http://real.mtak.hu/48951/1/Osi_et_al_MS_corrected.pdf
- Vickaryous, M. K. 2006. New information on the cranial anatomy of Edmontonia rugosidens Gilmore, a Late Cretaceous nodosaurid dinosaur from Dinosaur Provincial Park, Alberta. Journal of Vertebrate Paleontology 26 (4):1011—1013.
- Hill, R. V., D’Emic, M. D., Bever, G. S., Norell, M. A. 2015. A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia. Zoological Journal of the Linnean Society 175 (4): 892—909.
- T. Kubo, W. Zheng, M. O. Kubo & X. Jin. 2021. Dental microwear of a basal ankylosaurine dinosaur, Jinyunpelta and its implication on evolution of chewing mechanism in ankylosaurs. PLoS ONE 16(3): e0247969
- Han, F., Zheng, W., Hu, D., Xu, X., Barrett, P. M. 2014. A new basal ankylosaurid (Dinosauria: Ornithischia) from the Lower Cretaceous Jiufotang Formation of Liaoning Province, China. PLoS ONE 9 (8)
- Xu X, Wang X-L, You H-L (2001) A juvenile ankylosaur from China. Naturwissenschaften 88: 297—300.
- Carpenter, K. 2004. Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America. Canadian Journal of Earth Sciences 41: 961—986.
- Blows, W. T. and Honeysett, K. 2014. New teeth of nodosaurid ankylosaurs from the Lower Cretaceous of Southern England. Acta Palaeontologica Polonica 59 (4): 835—841.
- Carpenter, 2012, р. 509
- Coombs, W. P. Jr. 1990. Teeth and taxonomy in ankylosaurs. Pp. 269—279. In: Carpenter, K., Currie, P. J. (eds.). Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press.
- Rybczynski, N., Vickaryous, M. K. 2001. Evidence of complex jaw movement in the Late Cretaceous ankylosaurid Euoplocephalus tutus (Dinosauria: Thyreophora) In: Carpenter K, ed. The armored dinosaurs. Bloomington: Indiana University Press. Pp. 299—317.
- Carpenter, K. 2001. Phylogenetic analysis of the Ankylosauria. Pр. 455—482. In Carpenter, Kenneth (eds). The Armored Dinosaurs. Indiana University Press.
- Туманова, 1987, с. 53
- Kinneer, B.; Carpenter, K.; Shaw, A. 2016. Redescription of Gastonia burgei (Dinosauria: Ankylosauria, Polacanthidae), and description of a new species. Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen. 282 (1): 37-80.
- Carpenter, 2012, р. 510
- Ősi, A., Barrett, P., Földes, T., Tokai, R. 2014. Wear pattern, dental function, and jaw mechanism in the Late Cretaceous ankylosaur Hungarosaurus. The Anatomical Record Advances in Integrative Anatomy and Evolutionary Biology 297 (7): 1165—1180.
- Туманова, 1987, с. 43
- Coombs, W. P. 1971. The Ankylosauria. PhD dissertation, Columbia University, New York. 487 p.
- Carpenter, K. 2012. Ankylosaurs. Pp. 505—525. In J. Farlow and M. Brett-Surman (eds.). The Complete Dinosaur. Edition: 2nd. Bloomington: Indiana University Press.
- Carpenter, K. 1997. Ankylosauria. Pp. 16—20. In P. Currie and K. Padian (eds.). The Encyclopedia of Dinosaurs. Academic Press, San Diego.
- Russell, L.S., 1940. Edmontonia rugosidens (Gilmore), an armoured dinosaur from the Belly River Series of Alberta. Univ. Toronto Stud. Geol. Ser. 43,1-28
- Kirkland, J. and Carpenter, K. 1994. North America’s first pre-Cretaceous ankylosaur (Dinosauria) from the Upper Jurassic Morrison Formation of Western Colorado. Brigham Young University Geology Studies, 40: 25—42
- Blows, W. T. 2015. British Polacanthid Dinosaurs. Siri Scientific Press, Manchester, 224 p.
- Arbour, V. M., Larson, D., Vavrek, M., Buckley, L., and Evans, D. 2020. An ankylosaurian dinosaur from the Cenomanian Dunvegan Formation of northeastern British Columbia, Canada. Fossil Record 23: 179—189.
- Makovicky, P. 1997. Postcranial axial skeleton, comparative anatomy. Pp. 584—585. In P. Currie and K. Padian (eds.). The Encyclopedia of Dinosaurs. Academic Press, San Diego.
- Currie and Padian, 1997, p. 588
- Vickaryous, Maryańska, Weishampel, 2004, p. 380
- Kirkland, J. I., Alcala, L., Loewen, M. A., Espilez, E., Mampel, L., and Wiersma, J. P. 2013. The basal nodosaurid ankylosaur Europelta carbonensis n. gen. n. sp., from theLower Cretaceous (Lower Albian) Escucha Formation ofnortheastern Spain. PloS ONE, 8(12): e80405.
- Killbourne, B. and Carpenter, K. 2005. Redescription of Gargoyleosaurus parkpinorum, a polacanthid ankylosaur from the Upper Jurassic of Albany County, Wyoming. Neues Jahrbuch für Geologie und Paläontologie 237: 111–160.
- Romer A. S. 1976. The Axial Skeleton. Pp. 218–297. Osteology of the Reptiles. The University of Chicago Press.
- Vickaryous, Maryańska, Weishampel, 2004, p. 382
- Currie and Padian, 1997, p. 534
- Vickaryous, Maryańska, Weishampel, 2004, p. 383
- Carpenter, K., DiCroce, T., Kinneer, B., Simon, R. 2013. Pelvis of gargoyleosaurus (Dinosauria: Ankylosauria) and the origin and evolution of the ankylosaur pelvis. PLoS One, 8 (11): e79887.
- Arbour, V. M. & Currie, P. J. 2015. Ankylosaurid dinosaur tail clubs evolved through stepwise acquisition of key features. Journal of Anatomy 227 (4): 514—523.
- Arbour, V. M., Burns, M. E., Sissons, R. L. 2009. A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus. Journal of Vertebrate Paleontology 29: 1117—1135.
- Arbour, V. M. 2009. Estimating impact forces of tail club strikes by ankylosaurid dinosaurs. PLoS ONE 4 (8): e6738.
- Coombs, W. P. 1995. Ankylosaurian tail clubs of middle Campanian to early Maastrichtian age from western North America, with description of a tiny club from Alberta and discussion of tail orientation and tail club function. Canadian Journal of Earth Sciences 32: 902—912.
- Arbour, V. M., Snively, E. 2009. Finite element analyses of ankylosaurid dinosaur tail club impacts. The Anatomical Record Advances in Integrative Anatomy and Evolutionary Biology 292 (9): 1412—1426.
- Carpenter, K., Hayashi, S., Kobayashi, Y., et al. 2011. Saichania chulsanensis (Ornithischia, Ankylosauridae) from the Upper Cretaceous of Mongolia. Palaeontographica, Abteilung A 293, 1—61.
- Arbour, V. M., Currie, P. J. 2013. The taxonomic identity of a nearly complete ankylosaurid dinosaur skeleton from the Gobi Desert of Mongolia. Cretaceous Research 46, 24—30.
- Christiansen, P. 1997. Forelimbs and Hands. Pp. 245—253. In P. Currie and K. Padian (eds.). The Encyclopedia of Dinosaurs. Academic Press, San Diego.
- Vickaryous, Maryańska, Weishampel, 2004, p. 385.
- Senter, F. 2011. Evidence for a sauropod−like metacarpal configuration in ankylosaurian dinosaurs. Acta Palaeontologica Polonica 56 (1): 221—224. http://www.app.pan.pl/archive/published/app56/app20100041.pdf
- Currie, P. J., Badamgarav, D., Koppelhus, E. B., Sissons, R. & Vickaryous, M. K. 2011. Hands, feet, and behaviour in Pinacosaurus (Dinosauria: Ankylosauridae). Acta Palaeontologica Polonica 56: 489—504.
- Francischini, H., Sales, M., Schultz, C. L. 2017. The Presence of Ankylosaur Tracks in the Guará Formation (Brazil) and Remarks on the Spatial and Temporal Distribution of Late Jurassic Dinosaurs. Ichnos 25 (3): 177—191.
- Carpenter, K. 1990. Ankylosaur systematics: example using Panoplosaurus and Edmontonia. Pp. 281—298. In K. Carpenter and P. J. Currie (eds.). Dinosaur Systematics: Approaches and Perpectives. Cambridge University Press, Cambridge.
- Pereda-Suberbiola, X., Dantas, P., Galton, P. M., and Sanz, J. L. 2005. Autopodium of the holotype of Dracopelta zbyszewskii (Dinosauria, Ankylosauria) and its type horizon and locality (Upper Jurassic: Tithonian, western Portugal). Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen, 235 (2): 175—196.
- Vickaryous, Maryańska, Weishampel, 2004, p. 386.
- Christiansen, P. 1997. Hindlimbs and Feet. Pp. 320—328. In P. Currie and K. Padian (eds.). The Encyclopedia of Dinosaurs. Academic Press, San Diego.
- Vickaryous, Maryańska, Weishampel, 2004, p. 387.
- Малеев, Е. А. 1956. Панцирные динозавры верхнего мела Монголии (Семейство Анкилозавриды). Труды Палеонтологического Института Академии Наук СССР, 62: 51—91.
- Penkalski, P. & Blows, W. T. 2013. Scolosaurus cutleri (Ornithischia: Ankylosauria) from the Upper Cretaceous Dinosaur Park Formation of Alberta, Canada. Canadian Journal of Earth Sciences 50 (2): 171—182.
- Carpenter, 2012, р. 505
- Ankylosaurus- The Bullet-Proof Dinosaur ¦ Newsgrape.com. web.archive.org (10 мая 2012). Дата обращения: 7 января 2021.
- Carpenter, 2012, pр. 515—516
- Gaston, R. W., Schellenbach, J. and Kirkland, J. I. 2001. Mounted Skeleton of the Polacanthine Ankylosaur Gastonia burgei. Pp. 386—398. In K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- Туманова, 1987, с. 49—50
- Blows, W. T. 2013. Notice of nodosaur (Dinosauria, Ankylosauria) remains from the mid-Cretaceous of Cambridge, England, with comments on cervical half-ring armour. Proceedings of the Geologists Association 125 (1): 56—62.
- Burns, M. E., Currie, P. J. 2014. External and internal structure of ankylosaur (Dinosauria, Ornithischia) osteoderms and their systematic relevance. Journal of Vertebrate Paleontology 34: 835—851.
- Arbour, V. M., Burns, M. E. and Currie, P. J. 2011. A review of pelvic shield morphology in ankylosaurs (Dinosauria: Ornithischia). Journal of Paleontology 85 (2): 298—302.
- Botfalvai, G., Prondvai, E. and Osi, A. 2020. Living alone or moving in herds? A holistic approach highlights complexity in the social lifestyle of Cretaceous ankylosaurs. Cretaceous Research 118
- Carpenter, 2012, р. 516
- Blows, W. T. 2001. Dermal armor of the polacanthine ankylosaurs. Pp. 363—385. In: K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
- Arbour, V. M., Zanno, L. E. 2019. Tail Weaponry in Ankylosaurs and Glyptodonts: An Example of a Rare but Strongly Convergent Phenotype. The Anatomical Record Advances in Integrative Anatomy and Evolutionary Biology 303.
Литература
- Carpenter, K. Ankylosaurs. (англ.) // J. Farlow and M. Brett-Surman. The Complete Dinosaur. Edition: 2nd. — Bloomington: Indiana University Press, 2012. — P. 505—525.
- Weishampel D. B., Dodson, P., Osmólska H., eds. The Dinosauria (англ.). — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p.
- Туманова, Т. А. Панцирные динозавры Монголии. Совместная Советско-Монгольская палеонтологическая экспедиция. // Наука. — 1987. — Вып. 37. — С. 1—85.