Лабиринтодонты
Лабиринтодонты[1][2][3] или лабиринтозубые[1] (лат. Labyrinthodontia, от др.-греч. λαβύρινθος — «лабиринт», ὀδούς — «зуб»), — группа вымерших амфибий, обитавших на Земле в палеозойскую и мезозойскую эры (около 390—150 миллионов лет назад). Традиционно имела ранг подкласса; в современной литературе часто рассматривается как безранговое и неформальное объединение в связи с парафилетичностью. Тем не менее выделение этой группы оказалось удобным и её продолжают использовать[4][5].
| † Лабиринтодонты | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Реконструкция Seymouria | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Класс: Подкласс: † Лабиринтодонты |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Labyrinthodontia Owen, 1860 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 390—150 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Основные отличительные черты

Лабиринтодонты процветали на протяжении более чем 200 миллионов лет. И хотя разные виды существенно отличались один от другого, всё же можно выделить некоторые их общие черты:
|
Строение организма

Общее строение
По строению лабиринтодонты были подобны амфибиям. У большинства из них были короткие конечности, крупная голова и относительно короткий хвост. Представители многих групп лабиринтодонтов (особенно ранние формы) имели крупные размеры. Примитивные виды всех групп были, судя по имеющимся данным, водными хищниками. Лабиринтодонты вели земноводный образ жизни, который у разных видов значительно отличался и мог приближаться к чисто сухопутному или к водному[9]. Некоторые эволюционные линии лабиринтодонтов остались или вторично стали полностью водными, с редуцированными конечностями и угреобразным телом.
Череп

За исключением змеевидных Aistopoda, лабиринтодонты имели массивный череп. Широкая голова и короткая шея указывают на то, что лёгочное дыхание лабиринтодонтов (как и у современных земноводных) осуществлялось с помощью колебания ротоглоточной полости[10]. Череп имел заметные ушные вырезки позади каждого глаза и теменное отверстие.
Челюсти были усыпаны маленькими острыми коническими зубами, а по краю нёба были крупные клыки. Замена зубов происходила волнами, таким образом, что рядом с каждым сформированным зубом был подрастающий, готовый его заменить[11]. У всех видов лабиринтодонтов зубы обладали своеобразной структурой (изгибы складок дентина, которые на поперечном срезе зуба образуют извилистый узор), что и дало название этим существам. Единственным исключением были зубилообразные зубы поздних фитофагов — Diadectomorpha[10]. Подобный складчатый дентин получил название «плицидентин». По мере приближения к вершине зуба лабиринтовый узор упрощался и исчезал. Снаружи зубы несли продольные бороздки, отражающие складки дентина, и иногда были покрыты тонкой эмалью[12][13].
Лабиринтовые зубы были унаследованы лабиринтодонтами от рыбообразных предков (рипидистий)[14]. Кроме рипидистий и лабиринтодонтов, такие зубы встречаются у панцирных щук, ихтиозавров, хампсозавров, варанообразных и мозазавров и, видимо, независимо возникали не менее 4 раз[12][13]. По версии Роберта Кэрролла, такое строение зуба служило для его упрочнения в связи с высокими нагрузками при резком схлопывании челюстей, а у более продвинутых тетрапод исчезло из-за более тонкого и точного орудования челюстями[15][10]. Согласно исследованию 1991 года, лабиринтовое строение упрочняет недосформированные зубы на десятки процентов, хотя практически не влияет на прочность готовых. Это могло быть связано с тонкими челюстями лабиринтодонтов, внутри которых растущие зубы поместиться не могли. Как следствие, зубы начинали работать и нуждаться в высокой прочности до окончательного формирования[12].
Посткраниальный скелет
Позвонки были сложными и относительно хрупкими, они состояли из многочисленных элементов[6]. Трубчатые кости конечностей были короткими и широкими, голени обладали ограниченной подвижностью, а окончания конечностей не несли когтей, что ограничивало манёвренность этих животных[6][16]. Во взрослом возрасте большинство видов, скорее всего, старались держаться около водоёмов. Некоторые группы, жившие на закате палеозойской эры, в частности, микрозавры и сеймуриаморфы, были небольшими или среднего размера существами и, по современным представлениям, обитали на суше. Поздние Diadectomorpha и ранние формы пермского периода были полностью сухопутными формами с прочными скелетами и наиболее массивными обитателями суши своего времени. Мезозойские лабиринтодонты были в основном водными животными с преимущественно хрящевым скелетом[17].
Органы чувств
Глаза большинства лабиринтодонтов были высокопосаженными, что давало хороший обзор в верхней полусфере, но очень ограниченное боковое зрение. Теменной глаз был хорошо развит, хотя неясно, мог ли он давать изображение или только различать свет и тьму, как у современных гаттерий.
Большинство лабиринтодонтов обладали специальными сенсорными органами на коже — органом боковой линии для восприятия потоков и давления воды, как у рыб и некоторых современных амфибий[18]. Это позволяло им улавливать вибрации и звуки, издаваемые добычей во время охоты в мутной воде, в условиях плохой видимости.
Более древние лабиринтодонты имели массивные слуховые косточки, вероятно, прикреплявшиеся к черепу, в том числе к дерматокраниуму. Является дискуссионным, имели ли ранние наземные виды лабиринтодонтов слуховые косточки, связанные с барабанной перепонкой, покрывавшей их ушную вырезку, если, конечно, лабиринтодонты в принципе могли слышать, находясь на суше[19].
Барабанная перепонка бесхвостых (Anura) и амниот, судя по всему, возникла независимо. Это говорит в пользу того, что большинство лабиринтодонтов, если не все они, не могли слышать звуков, распространяющихся по воздуху[20].
Дыхательная система

У ранних лабиринтодонтов были хорошо развитые внутренние жабры, а также лёгкие, унаследованные от предков-рыб[21]. Они могли дышать воздухом, что должно было быть большим подспорьем для обитателей прогреваемых солнцем отмелей с низким уровнем содержания кислорода в воде. Диафрагма у лабиринтодонтов отсутствовала, а рёбра многих видов были слишком короткими или расположенными близко одно к другому, чтобы лёгкие могли расширяться. Скорее всего, воздух нагнетался в лёгкие при помощи сокращений горлового мешка, как у современных амфибий. Это могло стать причиной сохранения очень плоского черепа у поздних форм. Выдох при помощи рёбер, возможно, был актуален только для видов, являвшихся предками амниот[10]. Многие водные формы лабиринтодонтов сохраняли личиночные жабры и во взрослом возрасте.
В период, когда атмосферное давление и содержание в воздухе кислорода и углекислого газа оставались высокими, примитивное дыхание при помощи горлового мешка могло быть эффективным даже для крупных существ. Но в воздушной среде выделение диоксида углерода (CO2) затруднено по сравнению с водной, и крупные лабиринтодонты, вероятно, могли выдерживать высокую концентрацию CO2 в крови и периодически возвращались в воду для его выделения через кожу[10]. Потеря кожной брони у предков рыб и современное дополнительное дыхание через кожу у амфибий делают такое объяснение вероятным[22].
Поведение
Охота и питание
Как и их предки, лопастепёрые рыбы, лабиринтодонты были плотоядны. Широкий череп и относительно слабая подчелюстная мышца не позволяли им открывать пасть очень широко. Вероятно, большинство из них в отличие от современных амфибий использовали тактику засад[23]. Когда потенциальная добыча оказывалась в воде в пределах досягаемости, открытая заранее челюсть захлопывалась, а острые зубы лабиринтодонта впивались в тело жертвы. Большие нагрузки, которые при этом приходились на зубы, требовали их упрочнения, а это могло привести к тому их строению, которое и дало название группе[15][10]. Глотание добычи производилось закидыванием головы назад и вверх, как это делают современные амфибии и крокодилы.
Эволюция параметров черепа, лучший контроль челюсти и редуцирование нёбных клыков заметны только у более продвинутых рептилиеобразных видов. Возможно, это было связано с повышением эффективности дыхания, что сделало возможным более изящный стиль охоты[10].
Воспроизводство
Воспроизводство лабиринтодонтов было устроено по типу амфибий. Они откладывали в воду яйца (икру), из которых затем выводились головастики. Последние оставались в воде до метаморфоза, после которого могли выбраться на сушу. Ископаемые головастики для нескольких видов лабиринтодонтов известны, они представляют собой неотеничные взрослые особи с перистыми наружными жабрами, подобными тем, что сегодня присутствуют у головастиков беспанцирных, мальков двоякодышащих и многопёровых. Наличие личиночной стадии как примитивной стадии развития у всех групп лабиринтодонтов можно предположить с достаточной степенью уверенности (подобное отмечалось у Discosauriscus, близких родственников амниот)[24].
Подгруппы лабиринтодонтов
Систематическое расположение различных групп внутри Labyrinthodontia известно своим непостоянством[25][26]. Выделяют несколько групп, но по поводу их родственных связей нет консенсуса[27]. Представители многих ключевых групп невелики и обладали не полностью окостеневшими скелетами; более того, существует разрыв в летописи находок останков в начале каменноугольного периода (известный как разрыв Ромера (англ. Romer's gap), когда развились большинство групп[25][28]. Ещё более усложняет картину то, что личиночная и взрослая стадия развития предков амфибий существенно отличались друг от друга, что делает возможными случаи ошибочной классификации личинок как отдельных видов[29] Представляется, что Labyrinthodontia состоит из нескольких клад[30]. Две наиболее изученные группы, ихтиостегалий и рептилиоморф, обычно считают парафилетическими[6]. Систематика лабиринтодонтов была главной темой первого собрания Международного общества филогенетической номенклатуры (International Society for Phylogenetic Nomenclature)[31][уточнить].
Ichthyostegalia

Ранние лабиринтодонты известны с девона и, возможно, из разрыва Ромера в начале каменноугольного периода. Этих лабиринтодонтов обычно помещают в отряд Ichthyostegalia, хотя данная группа является скорее градой (эволюционной ступенью), нежели кладой[32]. Представители отряда были в основном водными жителями, и большинство из них даже во взрослом возрасте сохраняли внутренние жабры. Скорее всего, они нечасто выбирались на сушу. Их конечности были устроены несколько иначе, нежели у четвероногих, и по форме были веслообразными[33]. Хвост был похож на рыбий[34]. Позвонки сложные и довольно хрупкие. В конце девона появились формы с более сильными конечностями и крепкими позвонками, которые во взрослом возрасте уже не имели функциональных жабр. Все они были преимущественно водными видами, а некоторые и вовсе проводили всю жизнь в воде.
Рептилиоморфы
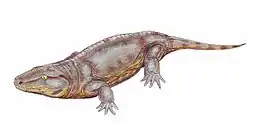
Ранними представителями лабиринтодонтов были рептилиоморфы (Reptiliomorpha), иногда синонимизируемые с Anthracosauria. Древнейшим известным их представителем может быть позднедевонский Tulerpeton; если это так, то рептилиоморфы возникли ещё до начала карбона[35]. Их черепа были глубже и уже, чем у других лабиринтодонтов. Передние и задние лапы большинства видов имели по пять пальцев. Некоторые ранние группы, обитавшие в солоноватой или морской водной средах, вернулись затем к более или менее полностью водной жизни[36].
За исключением Diadectomorpha, наземные формы имели средние размеры и появились в начале каменноугольного периода. Позвонки соответствовали таковым у примитивных рептилий, с маленькими плевроцентрами, впоследствии они эволюционировали. Наиболее известен род Seymouria. Некоторые члены наиболее эволюционно продвинутой группы, Diadectomorpha, были травоядными и вырастали до нескольких метров в длину. Они имели крупные бочкообразные тела. Более мелкие родственники Diadectomorpha стали предками первых рептилий. Это произошло в конце каменноугольного периода[37][38].
Темноспондильные (Temnospondyli)
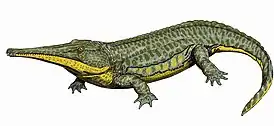
Наиболее разнообразной группой лабиринтодонтов были темноспондильные. Они появились в позднем девоне и различались по размеру от саламандроподобных Stereospondyli, которые сновали вдоль кромки воды и обитали в прибрежных зарослях, до гигантского бронированного Archegosauroidea, больше напоминавшего крокодила. Девятиметровый Prionosuchus был крупнейшей известной амфибией всех времён[39].

Передние ноги темноспондильных имели по четыре пальца, а задние по пять, как у современных амфибий[17]. У темноспондильных были консервативные позвоночные столбы, в которых плевроцентры оставались маленькими у примитивных форм, полностью исчезая у более продвинутых. Интерцентры принимали на себя вес животного, были крупными и образовывали полное кольцо[6]. Все они были более или менее плоскоголовыми с сильными или вторично слабыми позвоночниками и конечностями. Были также и полностью водные формы, как, например, Dvinosauria, и даже морские, например, Trematosauridae. В конце пермского или начале триасового периода темноспондильные могли дать начало лягушкам и саламандрам[30].
Тонкопозвонковые (Lepospondyli)

Тонкопозвонковые — небольшая группа не вполне ясного происхождения — развились в основном как некрупные животные. Их останки относятся к карбону и ранней перми Европы и Северной Америки. В отличие от прочих лабиринтодонтов, они имели простые позвонки из одного элемента[40]. Большинство из них вели водный образ жизни и сохранили наружные жабры. Тонкопозвонковые чаще всего были саламандрообразны, но одна группа, Aistopoda, была змеевидной, с гибкими, редуцированными черепами. Некоторые микрозавровые тонкопозвонковые были приземистыми и короткохвостыми. Представляется, что они были приспособлены к жизни на суше. Наиболее известен род Diplocaulus с бумерангообразной головой.
Положение тонкопозвонковых по отношению к другим группам лабиринтодонтов неясно, иногда их даже выделяют в отдельный подкласс[41]. Зубы представителей этой группы не лабиринтовые. Не вполне ясно, являются ли тонкопозвонковые естественным объединением или это сборная группа, включающая педоморфные формы и головастиков других лабиринтодонтов[42].
Эволюционная история

Лабиринтодонты появились в девонском периоде (398—392 миллиона лет назад) или раньше. Они произошли от костных рыб, а именно от лопастепёрых — вероятно, из группы Rhipidistia. Ближайшие современные родственники лабиринтодонтов (не считая их потомков — тетрапод) — двоякодышащие рыбы, тоже иногда относимые к рипидистиям. Наиболее ранние находки наземных форм — следы возрастом 395 миллионов лет, найденные в местечке Захелме, Польша. Их оставили животные, чьи конечности очень схожи с таковыми у ихтиостеги[43][44].
Болотные хищники
К середине девона наземные растения стабилизировали пресноводные экологические ниши, в результате чего развились болотные экосистемы, что привело к усложнению пищевых цепочек, которое давало видам новые возможности[45]. Ранние лабиринтодонты жили исключительно в водной среде, охотясь около берега или в каналах. Они унаследовали от рыбовидных предков лёгкие (до сих пор сохранившие дыхательную функцию у двоякодышащих и многопёровых рыб). Это позволяло им охотиться в воде, бедной кислородом (то есть в водоёмах с высокой температурой или гниющей органикой). У самых ранних форм, таких, как Acanthostega, позвоночник и конечности были практически непригодны для жизни на суше. Это контрастирует с принятыми ранее представлениями о том, что рыбы впервые вышли на сушу или в поисках пищи, как это делает современный илистый прыгун, или чтобы добраться до нового водоёма, когда пересох тот, в котором они жили изначально. Остатки ранних тетрапод обнаружены в морских отложениях, что позволяет предположить, что изначально они жили в морской или солоноватоводной среде. Это подкрепляется тем, что остатки ранних лабиринтодонтов находят по всему миру, что указывает на их распространение не только через пресные водоёмы, но и вдоль берегов.
Все первые лабиринтодонты были крупными или весьма крупными. Это делало их пребывание на суше затруднительным. Они сохраняли жабры и рыбоподобные плавники и хвосты. Тем не менее лабиринтодонтов можно уверенно отличить от рипидистиевых рыб, так как их клейтрум и лопатка отделены от черепа и формируют плечевой пояс, который берёт на себя вес передней части животного[18]. Все они были плотоядными, поедая рыбу и, возможно, выходя на берег, чтобы полакомиться морскими организмами в зоне прибоя, лишь позже превратившись в классических хищников девона[43]. Различные ранние формы для удобства сгруппированы в Ichthyostegalia.
Хотя размеры тела и пропорции представителей Ichthyostegalia оставались в течение их эволюции неизменными, конечности претерпели быстрые изменения. Прото-тетраподы, например, Elginerpeton и Tiktaalik, имели похожие на плавники конечности без явно выраженных пальцев, предназначенные в первую очередь для передвижения в воде, но подходящие и для того, чтобы перебираться из одного водоёма в другой по суше или преодолевать заполненные растениями каналы. Ichthyostega и Acanthostega имели веслообразные полидактильные конечности с крупными костяными пальцами, которые должны были помочь им перебираться по суше. Водные Ichthyostegalia процветали в приливных каналах и топях до конца девона, однако полностью исчезают из летописи находок при переходе к каменноугольному периоду[18].
Наземные формы
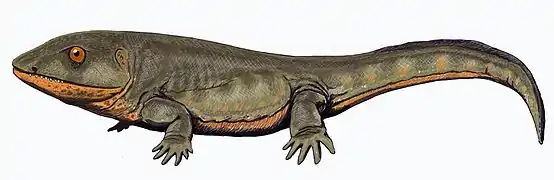
После массового вымирания в позднем девоне в летописи ископаемых имеется разрыв в 15 миллионов лет, известный как пробел Ромера. В результате исчезли Ichthyostegalia, а также более примитивные лабиринтодонты[9][18]. Известны находки этого периода из шотландского местонахождения East Kirkton Quarry своеобразных, возможно, повторно водных Crassigyrinus, которые могут быть сестринской группой позднейших лабиринтодонтов[46].
Ранний каменноугольный период был ознаменован появлением Loxommatidae — загадочной группы, которая также могла быть родственна более поздним и более продвинутым лабиринтодонтам[47]. К середине этого периода лабиринтодонты разделились, как минимум, на три ветви: темноспондильных, лепоспондильных и рептилиоморф. Последние были предками амниот.
Пока большинство лабиринтодонтов вело водный или наполовину водный образ жизни, некоторые рептилиеобразные амфибии начали покушаться на наземные экологические ниши, выступая как мелкие или средние хищники. В течение каменноугольного периода они эволюционировали в направлении обитания на суше, развивая более крепкие позвонки и стройные конечности, также видоизменялись череп и посадка глаз. Скорее всего, эти существа имели водонепроницаемую кожу с роговым эпидермисом, покрывающим небольшие костные узелки, образуя щитки, как у современных безногих земноводных. Современному наблюдателю эти животные показались бы грузными ящерицами, отличающимися от рептилий только отсутствием когтей и специфическими яйцами, откладываемыми в воду. В середине позднего карбона более мелкие формы эволюционировали в первых рептилий[37].
В конце каменноугольного периода произошло глобальное сокращение площади тропических лесов, в результате которого выиграли более приспособленные к жизни на суше животные. Те же лабиринтодонты, что сохранили амфибийный образ жизни, проиграли или вовсе не смогли приспособиться к новой реальности[48]. Некоторые рептилиеобразные амфибии, впрочем, процветали и в новых условиях. Рептилиоморфные Diadectidae с тяжёлыми бочкообразными телами, ставшие травоядными, стали крупнейшими наземными животными своего времени[18]. Также существовало семейство относительно крупных плотоядных Limnoscelidae, короткий расцвет которого пришёлся на конец каменноугольного периода.
Золотой век лабиринтодонтов

Травоядные Diadectidae достигли максимума разнообразия в позднем каменноугольном и раннем пермском периоде, после чего быстро ушли со сцены, а их место заняли ранние травоядные рептилии — Pareiasauridae и Edaphosauridae[40]. В отличие от рептилиеобразных амфибий, темноспондильные оставались в основном обитателями рек и болот. Питались рыбой и, возможно, другими лабиринтодонтами. Они пережили серьёзную диверсификацию после исчезновения большей части тропических лесов и в позднем каменноугольном и раннем пермском периоде достигли большого разнообразия, обитая в реках и солоноватых водоёмах угольных лесов в экваториальной Пангее и в окрестностях океана Тетис.
Изменения, способствующие питанию рыбой, развились у групп, в итоге развивших крокодилообразные черепа с вытянутыми мордами, и, возможно, соответствующий образ жизни (Archegosauridae, Melosauridae, Cochleosauridae, Eryopidae, и рептилиеобразные из подотряда Embolomeri)[40]. Другие же развились в водных хищников, использующих тактику засад, с короткими и широкими черепами, которые позволяли открывать пасть необходимым для охоты образом (Plagiosauridae и Dvinosauria)[49]. Они охотились как современные удильщики, а некоторые группы известны тем, что сохранили личиночные жабры на взрослой стадии развития, ведя полностью водный образ жизни. Так, Metoposauridae приспособились к охоте на отмелях и туманных болотах. Со своим ∩-образным черепом они были похожи на своих девонских предков.
В Евроамерике в каменноугольном периоде появились лепоспондильные — многочисленные мелкие, преимущественно водные амфибии неясного происхождения. Они обитали в подлеске тропических лесов и небольших прудах, то есть в экологической нише, аналогичной занимаемой современными амфибиями. В пермском периоде Nectridea распространились из Евроамерики в Гондвану.
Вымирание
С середины пермского периода климат стал более сухим, что осложнило выживание амфибий. Наземные рептилиоморфы исчезли, хотя водные крокодилообразные формы, такие как Embolomeri, смогли продержаться до триасового периода[6]. Из палеонтологической летописи исчезают и лепоспондильные, включая змееобразных Aistopoda.
С окончанием палеозойской эры большинство пермских групп пресеклись, за исключением Mastodonsauroidea и Metoposauridae, а также вызывающих особый интерес Plagiosauridae, которые продолжили существовать и в триасовом периоде. В раннем триасе эти группы пережили короткий серебряный век в водоёмах и на континентальных отмелях вместе с крупными видами, такими как тоозух[50] (Thoosuchus), бентозух (Benthosuchus) и эриозух[50] (Eryosuchus). Их экологические ниши были, возможно, теми же, что у современных крокодилов — речных хищников, питавшихся рыбой[40]. Все группы имели слабые позвонки, уменьшенное окостенение конечностей и более плоские черепа с выступающей боковой линией, что говорит о том, что темноспондильные триаса вряд ли проводили много времени в воде. Необычно крупный представитель брахиопид (вероятно, плагиозавр или близкий его родственник) достигал семиметровой длины, и возможно был столь же массивен как и Prionosuchus из пермского периода[51].
С началом процветания крокодилов в середине триасового периода наступил закат последних темпоспондильных, хотя некоторым из них удалось просуществовать как минимум до мелового периода в регионах со слишком холодным для крокодилов климатом на юге Гондваны[52].
Происхождение современных амфибий

В настоящее время существует консенсус учёных, что современные амфибии (Lissamphibia) так или иначе произошли от лабиринтодонтов. Но на этом консенсус и заканчивается[30]. Хрупкие кости амфибий плохо сохраняются в породе, а разрозненные костные останки лабиринтодонтов редки в принципе, что делает их сравнение трудным делом[18].
Традиционно предками современных амфибий считаются лепоспондильные. И те, и другие обычно невелики, имеют простые позвонки и занимают сходную экологическую нишу. Биологи считают, что лепоспондильные произошли от лабиринтодонтов, а амфибии — от лепоспондильных[38][53]. Некоторые учёные считают лепоспондильных близкими родственниками или даже предками рептилиеобразных амфибий[54].[55][56] Основной проблемой для таких мнений является вопрос о том, монофилетичны ли сами лепоспондильные[28][57].
В других работах рассматриваются отношения темноспондильных и амфибий[18][58][59][60]. Группа темноспондильных Amphibamidae считается возможным предком современных амфибий. Один из входящих в неё видов, Gerobatrachus, описанный в 2008 году, может быть переходной формой между темноспондильными и Anura (то есть, лягушками и жабами) и саламандрами. Он похож и на лягушек, и на саламандр, включая широкий череп, короткий хвост и некрупные зубы[61].
Картину осложняет вопрос о том, являются ли сами Lissamphibia полифилетическими. На этот счёт существует несколько мнений. «Стокгольмская школа» вслед за Gunnar Säve-Söderbergh и Erik Jarvik в течение большей части XX столетия утверждала, что амфибии в целом бифилетичны, ссылаясь на строение назальной капсулы и черепных нервов. По мнению этих учёных, лепоспондильные — предки лягушек, в то время как саламандры и безногие земноводные развивались независимо и происходят от поролепообразных рыб[62]. Роберт Лин Кэррол предлагал считать, что хвостатые амфибии (саламандры и безногие земноводные) происходят от лепоспондильных микрозавров, а лягушки — от темноспондильных[63]. Кладистический анализ Gerobatrachus показывает, что саламандры и лягушки развились из темноспондильных, а безногие земноводные являются сестринской группой для рептилеобразных амфибий[61]. Скорее всего, консенсус о происхождении амфибий и отношениях между разными группами лабиринтодонтов будет достигнут не скоро.
Происхождение амниот

Эволюционный переход от раннекарбоновых лабиринтодонтов к амниотам с начала XX века считался хорошо отражённым в палеонтологической летописи. Неясным оставалось в первую очередь то, где в этой последовательности форм произошла смена способа размножения с амфибийного на рептильный. В конце столетия работы Кэрролла и Лоурина очень помогли прояснению этого вопроса[64][65].
Ранние рептилиоморфы вели в основном водный образ жизни, первыми хорошо приспособленными к существованию на суше группами стали Seymouriamorpha и Diadectomorpha. Представители первой из них были животными небольшого или среднего размера, с крепкими конечностями. Их останки обнаруживали в сухих средах. При этом их кожа или чешуя имела водонепроницаемое покрытие (например, у Discosauriscus)[66]. Их скелеты очень похожи на кости ранних рептилий, а судя по находкам головастиков Seymouriamorpha, они сохраняли амфибийный способ воспроизводства[24]. Diadectomorpha считаются самыми близкими к амниотам. При этом их всё же располагают «на стороне» амфибий, хотя находки головастиков для этой группы пока неизвестны[67]. Однако анализ новых находок позволяет некоторым говорить, что филогенетика лабиринтодонтов может быть отличной от традиционно принятой в науке сегодня[26].
Некоторые авторы высказывали мнение, что «наземные» яйца развились из амфибийных, откладываемых на земле во избежание поедания хищниками — например, другими лабиринтодонтами[68][69]. Яйца амниот должны были обязательно развиться из яиц похожей структуры, как у современных амфибий[65]. При этом для успешного выделения углекислого газа яйца должны были быть очень маленькими, примерно по 1 сантиметру в диаметре, что накладывало серьёзные ограничения на размер взрослого животного. Таким образом, амниоты должны были развиться из очень маленьких существ[64]. Известно некоторое количество сильно фрагментированных останков возможных предков амниот из группы Diadectomorpha — Gephyrostegus[70] Solenodonsaurus[67], Westlothiana[71] и Casineria[37]. Ископаемые следы из Нью-Брансуика указывают на формирование первых рептилий не менее 315 миллионов лет назад[72].
История классификации

Термин «лабиринтодонт» был придуман Германом Бурмейстером в связи со структурой зубов этих существ[73]. Название Labyrinthodontia было впервые использовано в систематике Ричардом Оуэном в 1860 году, а в следующем году он отнёс их к Amphibia[74]. Лабиринтодонты были помещены в качестве отряда в клас Amphibia Уотсоном в 1920 году и — в качестве надотряда — Ромером в 1947[75][76]. Альтернативное название — Stegocephalia — было придумано в 1868 году американским палеонтологом Эдвардом Копом. Оно образовано из др.-греч. στέγος — «кровля» и κεφᾰλή — «голова» и связано с защитной бронёй некоторых крупных лабиринтодонтов[77]. Этот термин широко использовался в литературе XIX — начала XX века.
Классифицировать самые ранние находки пытались на основе крыши черепа, которой эти находки нередко и исчерпывались. Однако из-за изменчивости и быстрой эволюции этой структуры у лабиринтодонтов эти попытки не привели к успеху[75]. Родственные связи различных групп друг с другом и с современными амфибиями (а в некоторой степени и с рептилиями) до сих пор дискуссионны[30][60]. Существует несколько возможных схем этих взаимоотношений.
Классификация по позвонкам
В начале XX века лабиринтодонтов было принято классифицировать на основе морфологии элементов, составляющих их сложные позвонки[41]. Эта классификация быстро доказала свою несостоятельность, так как у далёких видов могло независимо возникать похожее строение, а близкие могли сильно отличаться. К середине столетия учёные заподозрили некоторые группы мелких лабиринтодонтов в том, что в действительности это головастики или неотеничные формы других групп[78]. Представленная здесь классификация позаимствована из работы Watson, 1920[75]:
- Отряд Stegocephalia (то есть лабиринтодонты)
- «Подразделение» Rachitomi (примитивные комплексные позвонки, все Ichthyostegalia, большинство крупных темноспондильных и некоторые рептилиеобразные амфибии)
- «Подразделение» Embolomeri (интерцентры и плевроцентры — цилиндры равного размера, сегодня считаются подотрядом вторично водных рептилиеобразных амфибий)
- «Подразделение» Stereospondyli (упрощённые позвонки, состоящие лишь из интерцентра и дуги; до сих пор признаются полноправной группой)
- Отряд Phyllospondyli (маленькие, хлипкие позвонки; сегодня считаются собранием головастиков и педоморфных форм)
- Отряд Lepospondyli (позвонки в форме песочных часов или цилиндрической формы, с середины каменноугольного до середины пермского периода, филогенетика неясна)
- Отряд Adelospondyli (позвонки цилиндрической формы с коническими впадинами на каждом конце, встречающимися в середине, сейчас их относят к лепоспондильным)
- Отряд Gymnophiona (существует поныне)
- Отряд Urodela (существует поныне)
- Отряд Anura (существует поныне)
Традиционная классификация
Традиционная классификация была введена Säve-Söderbergh в 1930-е годы. Он считал, что Amphibia бифелетичны и что саламандры и безногие земноводные развились из рыб независимо[62]. Эти взгляды разделялись немногими, но его схема взаимоотношений лепоспондильных и темноспондильных была взята за основу Ромером и использовалась в дальнейших исследованиях[38], а также с небольшими вариациями и другими авторами:[79] Daly 1973[80], Carroll 1988[81] и Hildebrand & Goslow 2001[82]. Ниже представлена классификация из работы Romer & Parson, 1985[6]:
- Подкласс Labyrinthodontia
- Отряд Ichthyostegalia (примитивные предки, например, Ichthyostega — с середины до конца девона)
- Отряд Temnospondyli (поздний девон и несколько позже, напр., Eryops, возможные предки современных амфибий)
- Отряд Anthracosauria (каменноугольный и пермский периоды, напр., Seymouria, предки ранних рептилий)
- Подкласс Lepospondyli (каменноугольный и пермский периоды, напр., Diplocaulus, малая группа, возможно, предки современных амфибий)
- Подкласс Lissamphibia (от пермского периода до настоящего времени)
- Отряд Gymnophiona (существует поныне)
- Отряд Urodela (существует поныне)
- Отряд Anura (существует поныне)
Филогенетическая классификация
Лабиринтодонты выпали из фавора в классификациях последнего времени, так как являются парафилетической группой — не включают всех потомков своего общего предка. Различные группы, традиционно относимые к Labyrinthodontia, сегодня разные авторы на основе кладистического анализа относят к стволовым тетраподам, базальным тетраподам, неамниотическим Reptiliomorpha и темноспондильным, которые оказываются монофилетическими или парафилетическими. Это отражает акцент современной кладистики на выяснении родственных связей. Термин «лабиринтодонты», потеряв строгий таксономический смысл, остаётся в употреблении как привычное название ранних земноводных[82], особенно имевших соответствующее строение зубов[83], и продолжает использоваться некоторыми современными учеными для удобства[5].
По большей части синонимичное название Stegocephalia Мишель Лорин применил не только к традиционно относимым к лабиринтодонтам существам, но и к их потомкам, и в таком значении этот термин используется кладистами[8]. Существует и неформальный термин с более широким значением — стволовые тетраподы (англ. stem Tetrapoda), включающий все виды, стоящие к современным тетраподам ближе, чем к двоякодышащим, но за исключением вершинной группы. Эта группа включает как традиционных «лабиринтодонтов», так и более базовых тетраподоморф, хотя её объём дискуссионен, так как родственные связи этих животных неясны[26].
Ниже представлено эволюционное древо, предложенное в работах Colbert, 1969[79] и Caroll, 1997[84]. Пунктирные линии обозначают отношения, относительно которых у разных авторов отсутствует консенсус.
| Sarcopterygii |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Хорошая справка (с диаграммой), дающая представление о характеристиках и основных эволюционных трендах для приведенных выше видов дается в работе Colbert 1969 pp. 102—103, но также следует изучить работу Kent & Miller (1997), в которой представлено альтернативное классификационное древо[41].
Примечания
- Основы палеонтологии : Справочник для палеонтологов и геологов СССР : в 15 т. / гл. ред. Ю. А. Орлов. — М. : Наука, 1964. — Т. 12 : Земноводные, пресмыкающиеся и птицы / под ред. А. К. Рождественского, Л. П. Татаринова. — С. 64. — 724 с. — 3000 экз.
- Банников А. Г., Денисова М. Н., Даревский И. С. Класс Земноводные, или Амфибии (Amphibia) // Жизнь животных. Том 5. Земноводные. Пресмыкающиеся / под ред. А. Г. Банникова, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1985. — С. 27. — 399 с.
- Лабиринтодонты // Куна — Ломами. — М. : Советская энциклопедия, 1973. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 14).
- Allaby M. Labyrinthodontia // Oxford Dictionary of Zoology. — Oxford University Press, 2003. — P. 285. — 597 p. — ISBN 9780198607588.
- Fins into limbs : evolution, development, and transformation (англ.) / edited by Brian K. Hall. — [Online-Ausg.].. — Chicago: University of Chicago Press, 2007. — P. 187. — ISBN 0226313379.
- Parsons, Alfred Sherwood Romer, Thomas S. The vertebrate body. — 6th ed.. — Philadelphia: Saunders College Pub., 1986. — ISBN 978-0-03-910754-3.
- Clack J. A. Devonian climate change, breathing, and the origin of the tetrapod stem group (англ.) // Integrative and Comparative Biology. — 2007. — 1 October (vol. 47, no. 4). — P. 510—523. — ISSN 1540-7063. — doi:10.1093/icb/icm055.
- Laurin M. (1998): The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I-systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: pp 1—42.
- Clack, J. A. (2002): Gaining ground: the origin and evolution of tetrapods. Indiana University Press, Bloomington, Indiana. 369 pp
- Janis, C.M.; Keller, J.C. Modes of ventilation in early tetrapods: Costal aspiration as a key feature of amniotes (англ.) // Acta Palaeontologica Polonica : journal. — Polish Academy of Sciences, 2001. — Vol. 46, no. 2. — P. 137—170.
- Osborn, J.W. The ontogeny of tooth succession in Lacerta vivipara Jacquin (1787) (англ.) // Proceedings of the Royal Society B : journal. — 1971. — 14 December (vol. 179, no. 56). — P. 261—289. — doi:10.1098/rspb.1971.0097. — PMID 4400215.
- Preuschoft H., Reif W.E., Loitsch C., Tepe E. The Function of Labyrinthodont Teeth: Big Teeth in Small Jaws // Constructional Morphology and Evolution / Schmidt-Kittler N., Vogel K. (eds). — Springer, 1991. — P. 151—171. — ISBN 978-3-642-76158-4. — doi:10.1007/978-3-642-76156-0_12.
- Warren A. A., Davey L. (1992). “Folded teeth in temnospondyls — a preliminary study”. Alcheringa: An Australasian Journal of Palaeontology. 16 (2): 107—132. DOI:10.1080/03115519208619036.
- Кэрролл Р. Палеонтология и эволюция позвоночных. — Москва: Мир, 1992. — Т. 1. — С. 204. — 280 с. — ISBN 5-03-001819-0.
- Carroll, R.L. Problem of the origin of reptiles (англ.) // Biological Reviews. — 1969. — 1 July (vol. 44, no. 3). — P. 393—431. — doi:10.1111/j.1469-185X.1969.tb01218.x.
- R. L. Paton, T. R. Smithson and J. A. Clack, «An amniote-like skeleton from the Early Carboniferous of Scotland», (abstract), Nature 398, 508—513 (8 April 1999)
- White, T. & Kazlev, M. A. (2006): Temnospondyli: Overview from Palaeos website
- Benton, Michael J. Vertebrate palaeontology. — 3rd ed.. — Oxford: Blackwell Science, 2004. — ISBN 978-0-632-05637-8.
- Laurin, M. (1996): Hearing in Stegocephalians, from the Tree of Life Web Project
- Lombard, R. E. & Bolt, J. R. (1979): Evolution of the tetrapod ear: an analysis and reinterpretation. Biological Journal of the Linnean Society No 11: pp 19—76. doi:10.1111/j.1095-8312.1979.tb00027.x
- Ромер А., Парсонс Т. Анатомия позвоночных. — Москва: Мир, 1992. — Т. 1. — С. 85. — 358 с. — ISBN 5-03-000291-X.
- Gordon, M.S.; Long, J.A. The Greatest Step In Vertebrate History: A Paleobiological Review of the Fish-Tetrapod Transition (англ.) // Physiological and Biochemical Zoology : journal. — 2004. — Vol. 77, no. 5. — P. 700—719. — doi:10.1086/425183. — PMID 15547790.
- Frazetta, T.H. Adaptive problems and possibilities in the temporal fenestration of tetrapod skulls (англ.) // Journal of Morphology. — Wiley-VCH, 1968. — Vol. 125, no. 2. — P. 145—158. — doi:10.1002/jmor.1051250203. — PMID 4878720.
- Špinar, Z. V. (1952): Revision of some Morovian Discosauriscidae. Rozpravy ustrededniho Uštavu Geologickeho no 15, pp 1—160
- Carroll, R. L. (2001): The origin and early radiation of terrestrial vertebrates. Journal of Paleontology no 75(6), pp 1202—1213 PDF Архивировано 30 сентября 2012 года.
- Ruta, M., Jeffery, J.E. and Coates, M.I. (2003): A supertree of early tetrapods. Proceedings of the Royal Society, London, series B no 270, pp 2507—2516 doi:10.1098/rspb.2003.2524
- Marjanović, David; Laurin, Michel. The origin(s) of extant amphibians: a review with emphasis on the "lepospondyl hypothesis" (англ.) // Geodiversitas : journal. — 2013. — 1 March (vol. 35, no. 1). — P. 207—272. — doi:10.5252/g2013n1a8.
- Carroll, R. L. (1995): Problems of the phylogenetic analysis of Paleozoic choanates. Bulletin du Muséum national d’Histoire naturelle de Paris, 4ème série 17: pp 389—445. Summary Архивная копия от 12 июня 2011 на Wayback Machine
- Steyer, J. S. (2000): Ontogeny and phylogeny in temnospondyls: a new method of analysis. Zoological Journal of the Linnean Society no 130, pp 449—467 PDF
- Laurin, M. (1996): Phylogeny of Stegocephalians, from the Tree of Life Web Project
- Foer, J. (2005): Pushing PhyloCode: What if we decide to rename every living thing on Earth?, Discovery Magazine, April issue
- Clack, J. A. (1997): Ichthyostega, from the Tree of Life Web Project
- Coates, M. I. & Clack, J. A. (1990): Polydactyly in the earliest known tetrapod limbs. Nature no 347, pp 66—67
- Jarvik, E. (1996): The Devonian tetrapod Ichthyostega. Fossils & Strata no 40: pp 1—213
- Lebedev, O.A., Coats, M.I. (2008): The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev. Zoological Journal of the Linnean Society no 114, Issue 3, pp 307—348. doi:10.1111/j.1096-3642.1995.tb00119.x abstract
- Garcia W.J., Storrs, G.W. & Grebe, S.F. (2006): The Hancock County tetrapod locality: A new Mississippian (Chesterian) wetlands fauna from Western Kentucky (USA). In Grebe, S.F. & DeMichele, W.A. (eds) Wetlands through time. pp 155—167. Geological Society of America, Boulder, Colorado.
- R. L. Paton, R. L., Smithson, T. R. & Clack, J. A. (1999): An amniote-like skeleton from the Early Carboniferous of Scotland (abstract), Nature 398, 508—513 (8 April 1999)
- Romer, A. S., (1947, revised ed. 1966) Vertebrate Paleontology, University of Chicago Press, Chicago
- Cox C. B., Hutchinson P. Fishes and amphibians from the Late Permian Pedrado Fogo Formation of northern Brazil (англ.) // Palaeontology : journal. — 1991. — Vol. 34. — P. 561—573.
- Colbert, E. H. & Morales, M. (1990): Evolution of the Vertebrates: A history of the Backboned Animals Through Time, John Wiley & Sons Inc (4th ed., 470 pp)
- Kent, G. C. & Miller, L. (1997): Comparative anatomy of the vertebrates. 8th edition. Wm. C. Brown Publishers. Dubuque. 487 pages. ISBN 0-697-24378-8
- White, T. & Kazlev, M. A. (2009): Lepospondyli: Overview, from Palaeos website.
- Niedźwiedzki G., Szrek P., Narkiewicz K. et al. Tetrapod trackways from the early Middle Devonian period of Poland (англ.) // Nature : journal. — 2010. — Vol. 463, no. 7277. — P. 43—48. — doi:10.1038/nature08623. — PMID 20054388.
- Uppsala University (2010, January 8). Fossil footprints give land vertebrates a much longer history. ScienceDaily. Retrieved January 8, 2010, from http://www.sciencedaily.com/releases/2010/01/100107114420.htm
- Beerbower, J. R., Boy, J. A., DiMichele, W. A., Gastaldo, R. A., Hook, R. & Hotton, N., Illustrations by Phillips, T. L., Scheckler, S. E., & Shear, W. A. (1992): Paleozoic terrestrial ecosystems. In: Behrensmeyer, A. K., Damuth, J. D., DiMichele, W. A., Potts, R., Sues, H. D. & Wing, S. L. (eds.) Terrestrial Ecosystems through Time, pp. 205—235. Chicago: Univ. Chicago Press summary from «Devonian Times»
- Milner, A. R. (1993): Amphibian-grade Tetrapoda. In The Fossil Record vol 2, (ed. Benton, M. J.) Chapman & Hall, London, pp 665—679
- Clack, J. A. (2006): Baphetidae, from the Tree of Life Web Project
- Sahney, S., Benton, M.J. & Falcon-Lang, H.J. Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica (англ.) // Geology : journal. — 2010. — Vol. 38, no. 12. — P. 1079—1082. — doi:10.1130/G31182.1.
- Jenkins, F. A., Jr., Shubin, N. H., Gatesy, S. M., Warren, A. (2008): Gerrothorax pulcherrimus from the Upper Triassic Fleming Fjord Formation of East Greenland and a reassessment of head lifting in temnospondyl feeding. Journal of Vertebrate Paleontology No 28 (4): pp 935—950. News article with animation of skull Архивная копия от 26 июля 2011 на Wayback Machine
- Палеонтологический музей имени Ю. А. Орлова / отв. ред. А. В. Лопатин. — М.: ПИН РАН, 2012. — 320 [376] с. — ISBN 978-5-903825-14-1.
- Steyer, J. S.; Damiani, R. A giant brachyopoid temnospondyl from the Upper Triassic or Lower Jurassic of Lesotho (англ.) // Bulletin de la Societe Geologique de France : journal. — 2005. — Vol. 176, no. 3. — P. 243—248. — doi:10.2113/176.3.243.
- Warren A. A., Kool L., Cleeland M. & al. (1991): An Early Cretaceous labyrinthodont. Alcheringa: An Australasian Journal of Palaeontology, Volume 15, Issue 4, pp 327—332. doi:10.1080/03115519108619027. Article from Informaworld
- Panchen, A.L. (1967): Amphibia, In Harland, W.B., Holland, C.H., House, M.R., Hughes, N.F., Reynolds, A.B., Rudwick, M.j.S., Satterthwaite, G.E., Tarlo, L.B.H. & Willey, E.C. (eds.): The Fossil Record, Geological Society, London, Special Publications, vol 2, chapter 27: pp 685—694; doi:10.1144/GSL.SP.1967.002.01.46 extract
- Laurin, M. & Reisz, R. R. (1997): A new perspective on tetrapod phylogeny. In Sumida, S. & Martin, K. (eds.) Amniotes Origins: Completing the Transition to Land, pp 9—59. London: Academic Press.
- Anderson, J. S. (2001): The phylogenetic trunk: maximal inclusion of taxa with missing data in an analysis of the Lepospondyli (Vertebrata, Tetrapoda). Systematic Biology No 50, pp 170—193 Article as PDF
- Vallin G. & Laurin, M. (2004): Cranial morphology and affinities of Microbrachis, and a reappraisal of the phylogeny and lifestyle of the first amphibians. Journal of Vertebrate Paleontology No 24: pp 56—72 Article as PDF
- Panchen, A. L. & Smithson, T. R. (1988): The relationships of the earliest tetrapods. In Benton, M. J. (ed.) The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds, pp 1—32. Oxford: Clarendon Press.
- Trueb, L. & Cloutier, R. (1991): A phylogenetic investigation of the inter- and intrarelationships of the Lissamphibia (Amphibia: Temnospondyli). In: Schultze, H-P & Trueb, L. (eds.) Origins of the higher groups of tetrapods—Controversy and Consensus, pp 223—313. Comstock Publishing Associates, Ithaca
- Ahlberg, P. E.; Milner, A. R. The origin and early diversification of tetrapods (англ.) // Nature. — 1994. — Vol. 368, no. 6471. — P. 507—514. — doi:10.1038/368507a0. — .
- Sigurdsen, T.; Green, D.M. The origin of modern amphibians: a re-evaluation (англ.) // Zoological Journal of the Linnean Society. — Oxford University Press, 2011. — Vol. 162, no. 2. — P. 457—469. — doi:10.1111/j.1096-3642.2010.00683.x.
- Anderson, J. S.; Reisz, R. R.; Scott, D.; Fröbisch, N. B.; Sumida, S. S. A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders (англ.) // Nature : journal. — 2008. — Vol. 453, no. 7194. — P. 515—518. — doi:10.1038/nature06865. — PMID 18497824.
- Säve-Söderbergh G. Some points of view concerning the evolution of the vertebrates and the classification of this group (англ.) // Arkiv för Zoologi : journal. — 1934. — Vol. 26A. — P. 1—20.
- Carroll, R. L.; Holme, R. The skull and jaw musculature as guides to the ancestry of salamanders (англ.) // Zoological Journal of the Linnean Society : journal. — 1980. — Vol. 68. — P. 1—40. — doi:10.1111/j.1096-3642.1980.tb01916.x.
- Carroll R.L. (1991): The origin of reptiles. In: Schultze H.-P., Trueb L., (ed) Origins of the higher groups of tetrapods — controversy and consensus. Ithaca: Cornell University Press, pp 331—353.
- Laurin, M. The Evolution of Body Size, Cope's Rule and the Origin of Amniotes (англ.) // Systematic Biology : journal. — 2004. — Vol. 53, no. 4. — P. 594—622. — doi:10.1080/10635150490445706.
- Klembara J. & Meszáros, S. (1992): New finds of Discosauriscus austriacus (Makowsky 1876) from the Lower Permian of Boskovice furrow (Czecho-Slovakia). Geologica Carpathica No 43: pp 305—312
- Laurin, M. and Rize R.R. (1999): A new study of Solenodonsaurus janenschi, and a reconsideration of amniote origins and stegocephalian evolution. Canadian Journal of Earth Science, no 36 (8): pp 1239—1255 (1999) doi:10.1139/cjes-36-8-1239
- Romer, A. S. Origin of the amniote egg (англ.) // The Scientific Monthly. — 1957. — Vol. 85. — P. 57—63.
- Carroll, R. L. Quantitative aspects of the amphibian-reptilian transition (англ.) // Forma et. Functio : journal. — 1970. — Vol. 3. — P. 165—178.
- Margaret C. Brough and J. Brough. The Genus Gephyrostegus (англ.) // Philosophical Transactions of the Royal Society B. — 1967. — 1 June (vol. 252, no. 776). — P. 147—165. — doi:10.1098/rstb.1967.0006. — .
- Smithson, T.R.; Rolfe, W.D.I. Westlothiana gen. nov. :naming the earliest known reptile (англ.) // Scottish Journal of Geology. — 1990. — Vol. 26. — P. 137—138. — doi:10.1144/sjg26020137.
- Falcon-Lang, H.J.; Benton, M.J.; Stimson, M. Ecology of early reptiles inferred from Lower Pennsylvanian trackways (англ.) // Journal of the Geological Society. — 2007. — Vol. 164, no. 6. — P. 1113—1118. — doi:10.1144/0016-76492007-015.
- Burmeister, H. (1850): Die Labyrinthodonten aus dem Saarbrücker Steinkohlengebirge, Dritte Abtheilung: der Geschichte der Deutschen Labyrinthodonten Archegosaurus. Berlin: G. Reimer, 74 pp.
- Owen, R. (1861): Palaeontology, or a Systematic Summary of Extinct Animals and their Geological Relations. Adam and Charles Black, Edinburgh, pages 1—463
- Watson, D. M. S. The Structure, Evolution and Origin of the Amphibia. The "Orders' Rachitomi and Stereospondyli (англ.) // Philosophical Transactions of the Royal Society B : journal. — 1920. — 1 January (vol. 209, no. 360—371). — P. 1—73. — doi:10.1098/rstb.1920.0001.
- A. S. Romer (1947): Review of the Labyrinthodontia. Bulletin of the Museum of Comparative Zoology no 99 (1): pp 1—368, cited in The Paleobiology Database: Labyrinthodontia, Amphibia — Apsidospondyli
- Cope E. D. 1868. Synopsis of the extinct Batrachia of North America. Proceedings of The Academy of Natural Sciences of Philadelphia: pp 208—221
- Case, E. C. A Census of the determinable Genera of Stegocephalia (англ.) // Transactions of the American Philosophical Society : journal. — 1946. — Vol. 35, no. 4. — P. 325—420. — doi:10.2307/1005567.
- Colbert, E. H., (1969), Evolution of the Vertebrates, John Wiley & Sons Inc (2nd ed.)
- Daly, E. (1973): A Lower Permian vertebrate fauna from southern Oklahoma. Journal of Paleontology no 47(3): pages 562—589
- Carroll, R. L. (1988), Vertebrate Paleontology and Evolution, WH Freeman & Co.
- Hildebrand, M. & G. E. Goslow, Jr. Principal ill. Viola Hildebrand. Analysis of vertebrate structure (англ.). — New York: Wiley, 2001. — P. 429. — ISBN 0-471-29505-1.
- Buell, Donald R. Prothero ; with original illustrations by Carl. Evolution : what the fossils say and why it matters (англ.). — New York: Columbia University Press, 2007. — P. 224. — ISBN 0231139624.
- Carroll R. L. (1997). Patterns and Processes of Vertebrate Evolution. Cambridge University Press, Cambridge. 464 pages
Ссылки
- Кладограмма Reptiliomorpha на Palaeos.com.
- Подкласс Labyrinthodontia.
- Сравнительная анатомия позвоночных.
| Таксономия |
|---|